
Host Cyanobacteria Killing by Novel Lytic Cyanophage YongM: A Protein Profiling Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Separation, Purification, and Suspension Preparation
2.3. Genome Enrichment and Extraction
2.4. Infection and Sample Collection
2.5. Protein Extraction and Digestion
2.6. Proteome Sequencing
2.7. Data Analysis
3. Results and Analysis
3.1. Effect of YongM Infection on Host Growth
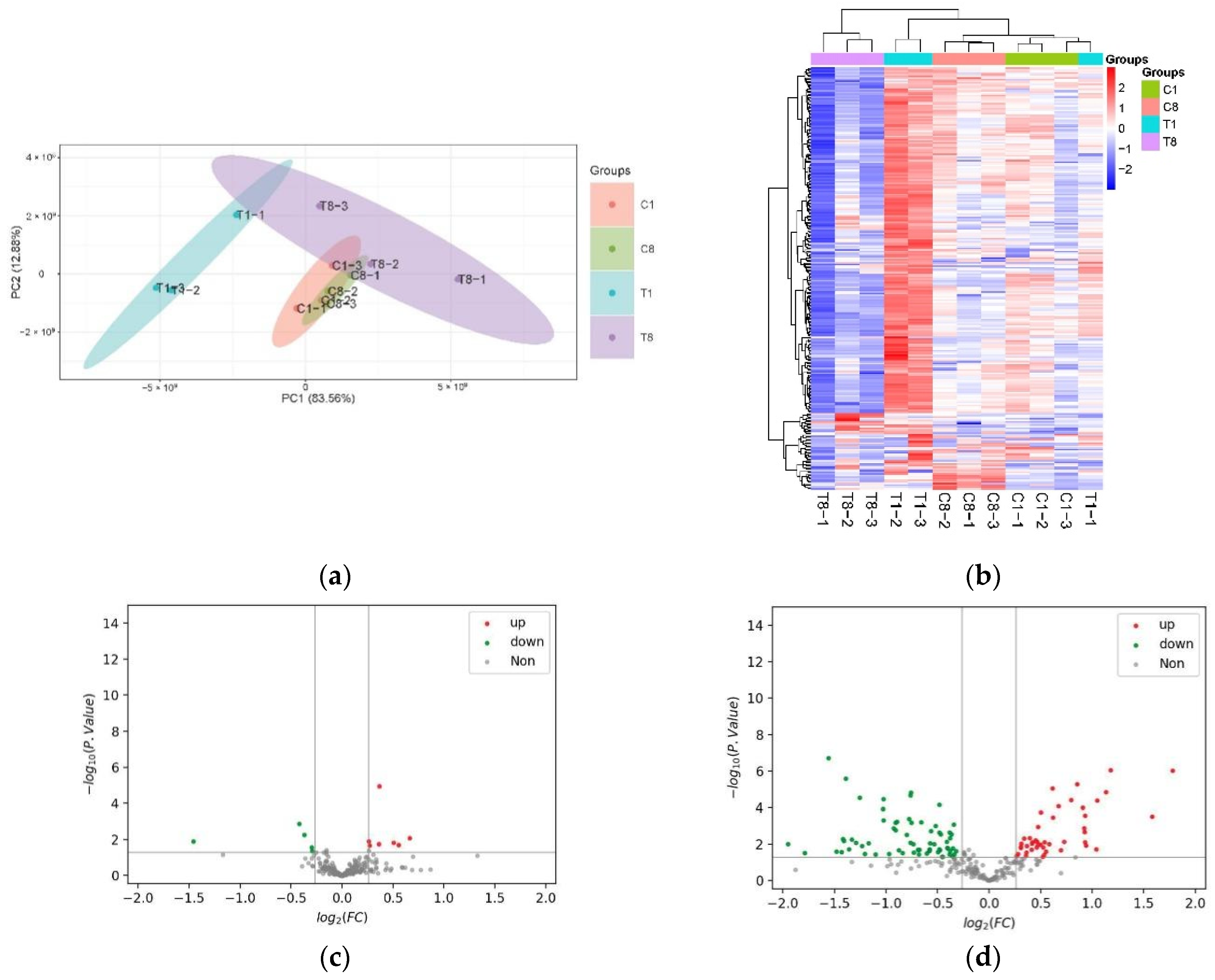
3.2. Altered FACHB-596 Protein Expression Profile Caused by YongM Infection
3.3. Functional Analysis of DEPs
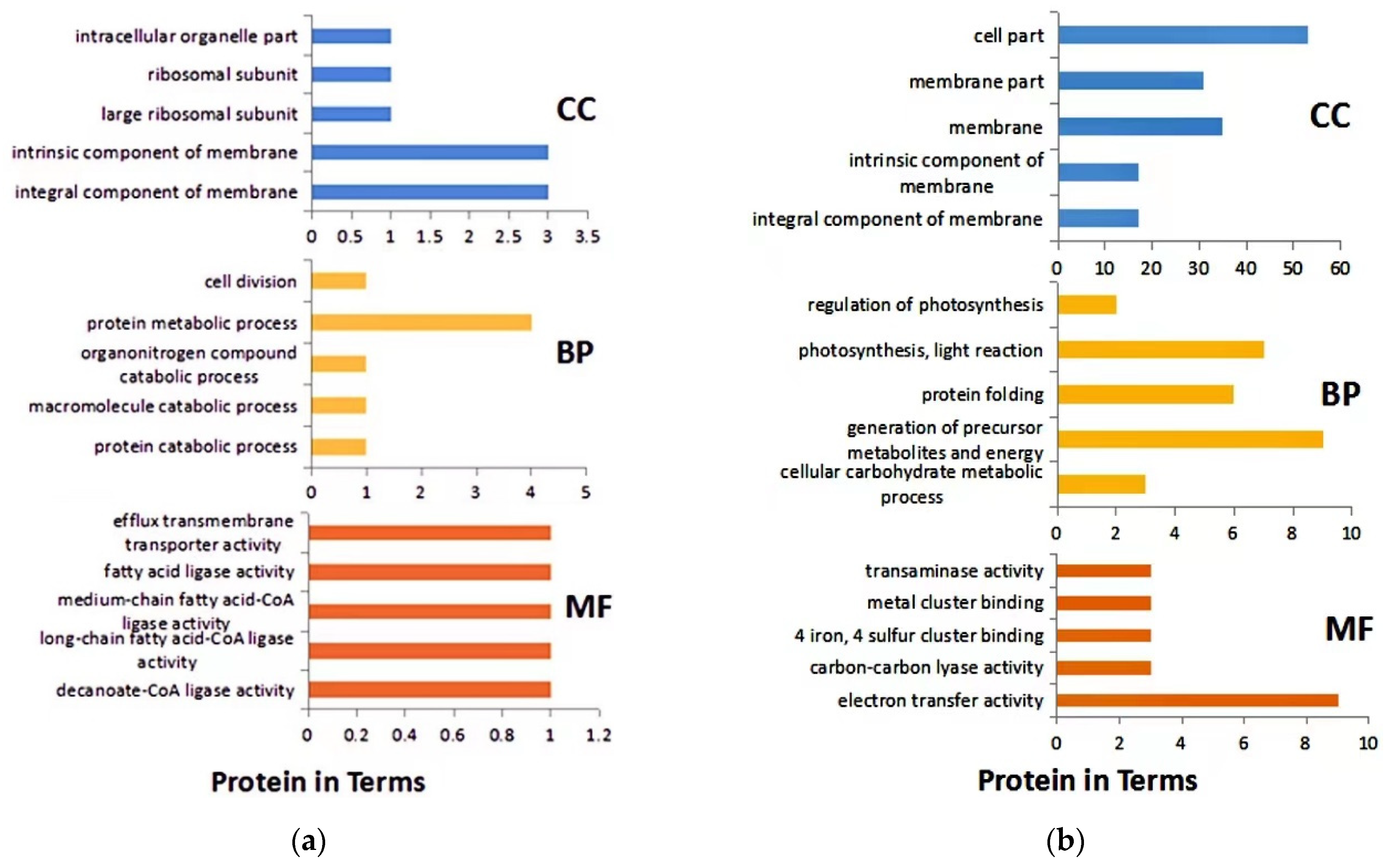
3.4. GO Analysis of DEPs
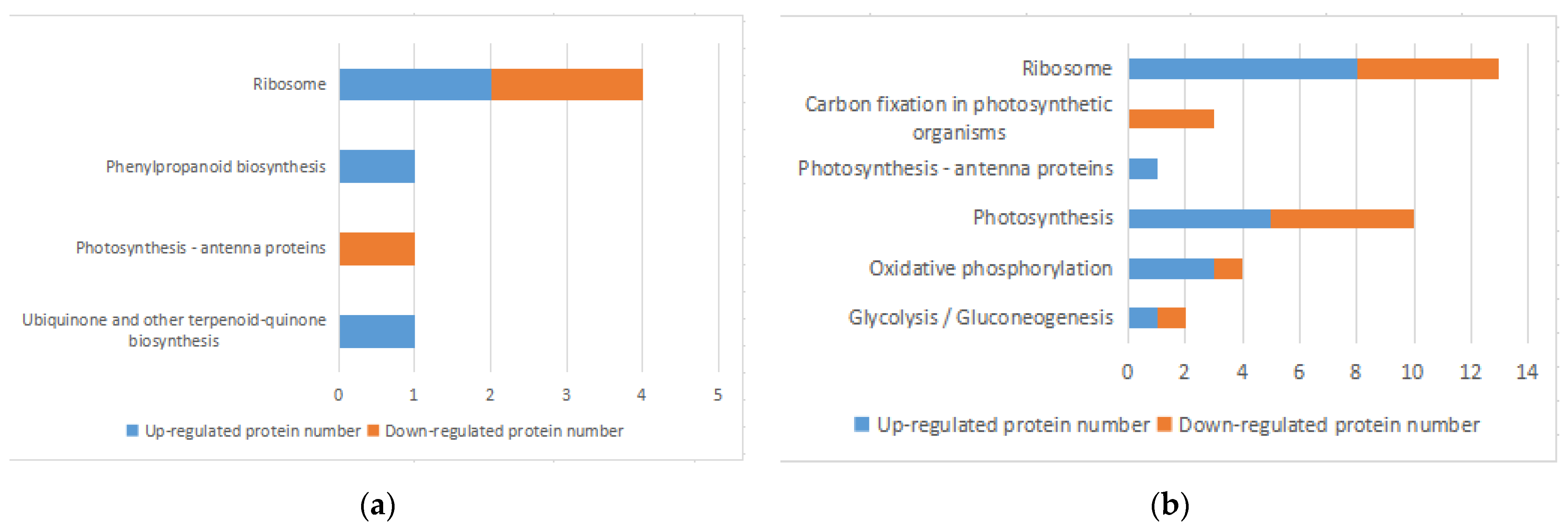
3.5. KEGG Pathway Analysis of DEPs
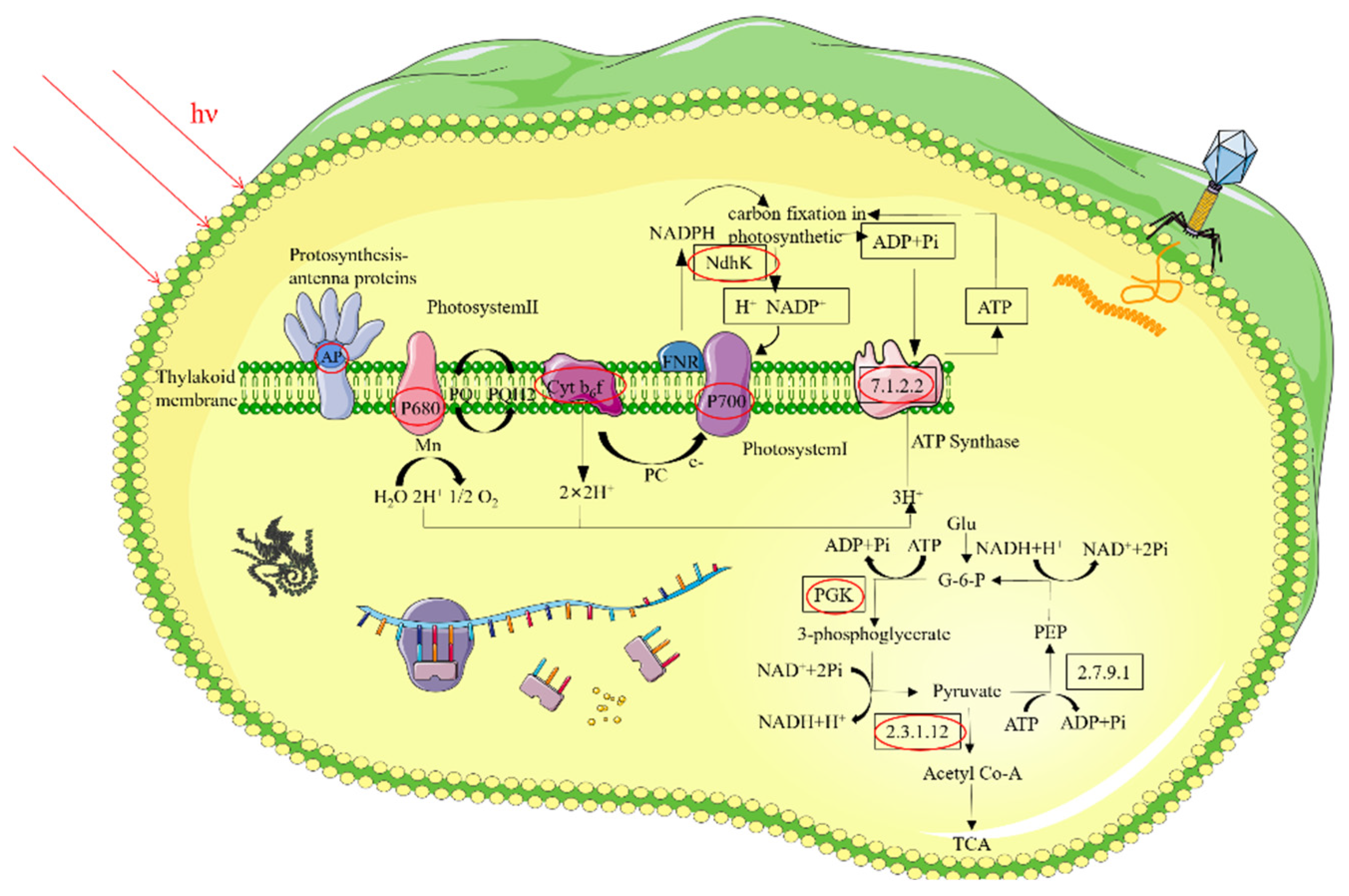
3.5.1. Analysis of Photosynthetic DEPs
3.5.2. Analysis of Energy Metabolism DEPs
3.5.3. Analysis of Translation Function DEPs
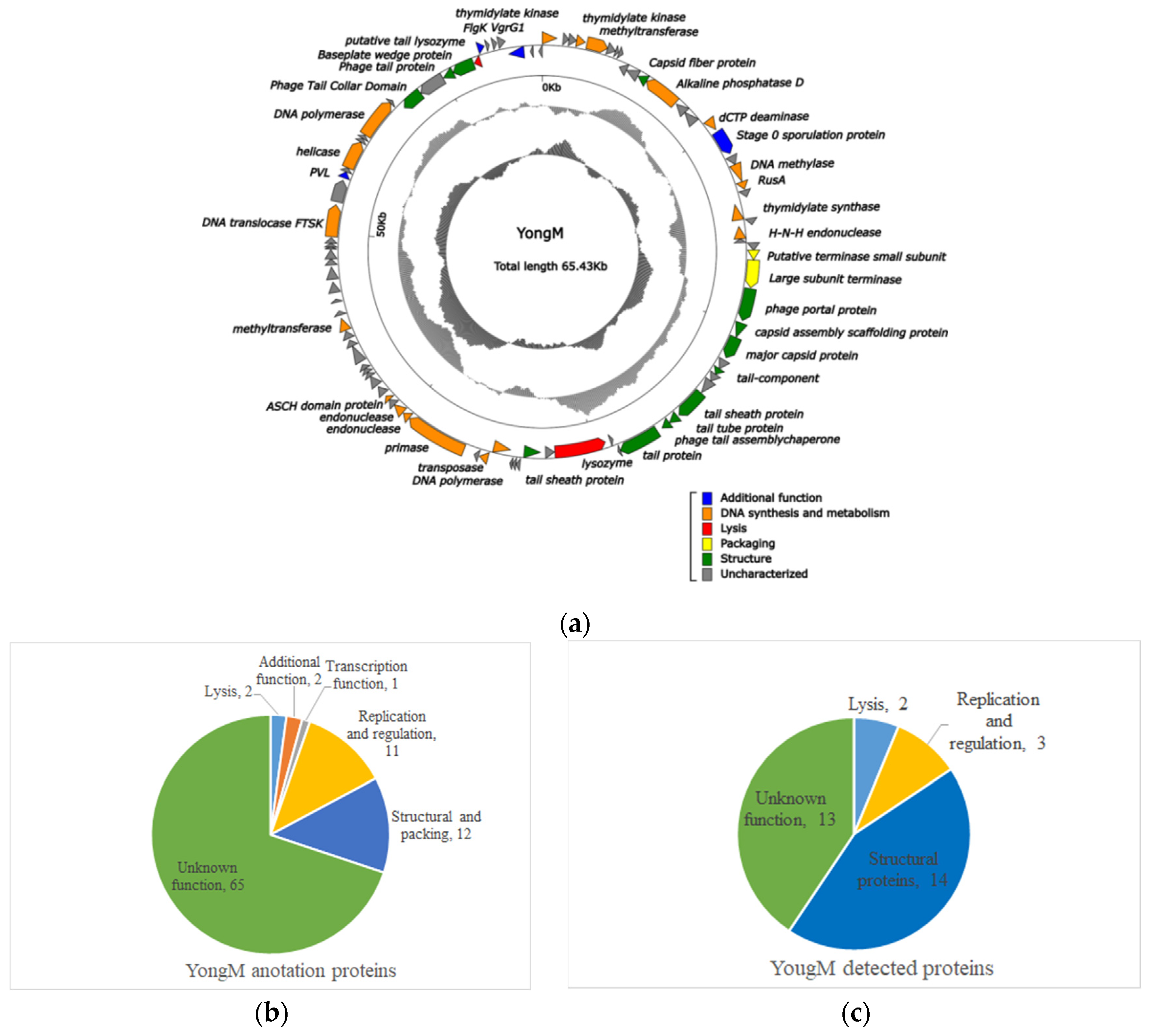
3.6. Structural and Functional Proteins of YongM
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilbert, P.M. Eutrophication, harmful algae and biodiversity—Challenging paradigms in a world of complex nutrient changes. Mar. Pollut. Bull. 2017, 124, 591–606. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, R.; Ye, J.; Xu, X.; Zhang, Y.; Wu, C. Monitoring and research of microcystins and environmental factors in a typical artificial freshwater aquaculture pond. Environ. Sci. Pollut. Res. 2018, 25, 5921–5933. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Chen, J.; Zhang, X.; Xie, P. A review of reproductive toxicity of microcystins. J. Hazard. Mater. 2016, 301, 381–399. [Google Scholar] [CrossRef]
- Suttle, C. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Safferman, R.S.; Cannon, R.E.; Desjardins, P.R.; Gromov, B.V.; Haselkorn, R.; Sherman, L.A.; Shilo, M. Classification and nomenclature of viruses of cyanobacteria. Intervirology 1983, 19, 61–66. [Google Scholar] [CrossRef]
- Mirzaei, M.K.; Eriksson, H.; Kasuga, K.; Haggård-Ljungquist, E.; Nilsson, A.S. Genomic, Proteomic, Morphological, and Phylogenetic Analyses of vB_EcoP_SU10, a Podoviridae Phage with C3 Morphology. PLoS ONE 2014, 10, e116294. [Google Scholar] [CrossRef] [Green Version]
- Safferman, R.S.; Morris, M.E. Algal virus:isolation. Science 1963, 140, 679–680. [Google Scholar] [CrossRef]
- Brussaard, C.; Wilhelm, S.W.; Thingstad, F.; Weinbauer, M.G.; Bratbak, G.; Heldal, M.; Kimmance, S.A.; Middelboe, M.; Nagasaki, K.; Paul, J.H.; et al. Global-scale processes with a nanoscale drive: The role of marine viruses. ISME J. 2008, 2, 575–578. [Google Scholar] [CrossRef] [Green Version]
- Roux, S.; Brum, J.R.; Dutilh, B.E.; Sunagawa, S.; Duhaime, M.B.; Loy, A.; Poulos, B.T.; Solonenko, N.; Lara, E.; Poulain, J.; et al. Ecogenomics and potential biogeochemical impacts of globally abundant ocean viruses Ocean microbes drive biogeochemical cycling on a global scale. Nature 2016, 537, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Li, D.F.; Sun, Z.T.; Tong, Y.; Yan, X.; Wang, C.; Zhang, X.; Pei, G. A novel freshwater cyanophage vB_MelS-Me-ZS1 infecting bloom-forming cyanobacterium Microcystis elabens. Mol. Biol. Rep. 2020, 47, 7979–7989. [Google Scholar] [CrossRef] [PubMed]
- Jonge, P.A.; Nobrega, F.L.; Brouns, S.J.J.; Dutilh, B.E. Molecular and Evolutionary Determinants of Bacteriophage Host Range. Trends Microbiol. 2019, 27, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Silpe, J.E.; Bassler, B.L. A Host-Produced Quorum-Sensing Autoinducer Controls a Phage Lysis-Lysogeny Decision. Cell 2019, 176, 268–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Nagasaki, K.; Takashima, Y.; Shirai, Y.; Tomaru, Y.; Takao, Y.; Sakamoto, S.; Hiroishi, S.; Ogata, H. Ma-LMM01 infecting toxic Microcystis aeruginosa illuminates diverse cyanophage genome strategies. J. Bacteriol. 2008, 190, 1762–1772. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Li, T.; Deng, F.; Hu, Z. Freshwater cyanophages. Virol. Sin. 2013, 28, 253–259. [Google Scholar] [CrossRef]
- Yoshida-Takashima, Y.; Yoshida, M.; Ogata, H.; Nagasaki, K.; Hiroishi, S.; Yoshida, T. Cyanophage infection in the bloom-forming cyanobacteria Microcystis aeruginosa in surface freshwater. Microbes Environ. 2012, 27, 350–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Kong, S.; Shi, M.; Fu, L. Genomic Analysis of Freshwater Cyanophage Pf-WMP3 Infecting Cyanobacterium Phormidium foveolarum: The Conserved Elements for a Phage. Microb. Ecol. 2008, 56, 671–680. [Google Scholar] [CrossRef]
- Liu, X.; Shi, M.; Kong, S.; Gao, Y.; An, C. Cyanophage Pf-WMP4, a T7-like phage infecting the freshwater cyanobacterium Phormidium foveolarum: Complete genome sequence and DNA translocation. Virology 2007, 366, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Zhong, K.X.; Suttle, C.A.; Baudoux, A.C.; Derelle, E.; Colombet, J.; Cho, A.; Caleta, J.; Six, C.; Jacquet, S. A New Freshwater Cyanosiphovirus Harboring Integrase. Front. Microbiol. 2018, 9, 2204. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; You, F.; He, Y.; Shu, H.; Gin, Y. Isolation and Characterization of the First Freshwater Cyanophage Infecting Pseudanabaena. J. Virol. 2020, 94, e00682-20. [Google Scholar] [CrossRef]
- Yang, F.; Jin, H.; Wang, X.; Li, Q.; Zhang, J.; Cui, N.; Jiang, Y.; Chen, Y.; Wu, Q.; Zhou, C.; et al. Genomic Analysis of Mic1 Reveals a Novel Freshwater Long-Tailed Cyanophage. Front. Microbiol. 2020, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Liu, X.; Wang, Q.; Lin, T.; Wang, M.; Liu, Q.; Shao, H.; Jiang, Y. The isolation and genome sequencing of a novel cyanophage S-H68 from the Bohai Sea, China. Mar. Genom. 2020, 53, 100739. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Liu, Q.; Liu, X.; Wang, Q.; Jiang, Y. Isolation and Complete Genome Sequence of a Novel Cyanophage S-B68. Curr. Microbiol. 2020, 77, 2385–2390. [Google Scholar] [CrossRef]
- Sukenik, A.; Kaplan, A. Cyanobacterial Harmful Algal Blooms in Aquatic Ecosystems: A Comprehensive Outlook on Current and Emerging Mitigation and Control Approaches. Microorganisms 2021, 9, 1472. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef]
- Ulius, S.; Mazur-Marzec, H.; Vitonyt, I.; Kvederaviciute, K.; Holmfeldt, K. Insights into cyanophage-mediated dynamics of nodularin and other non-ribosomal peptides in Nodularia spumigena. Harmful Algae 2018, 78, 69–74. [Google Scholar]
- Eigemann, F.; Schwartke, M.; Schulz-Vogt, H. Niche separation of Baltic Sea cyanobacteria during bloom events by species interactions and autecological preferences. Harmful Algae 2018, 72, 65–73. [Google Scholar] [CrossRef]
- Murata, K.; Zhang, Q.; Galaz-Montoya, J.G.; Fu, C.; Coleman, M.L.; Osburne, M.S.; Schmid, M.F.; Sullivan, M.B.; Chisholm, S.W.; Chiu, W. Visualizing Adsorption of Cyanophage P-SSP7 onto Marine Prochlorococcus. Sci. Rep. 2017, 7, 44176. [Google Scholar] [CrossRef]
- Waldbauer, J.R.; Coleman, M.L.; Rizzo, A.I.; Campbell, K.L.; Zhang, L. Nitrogen sourcing during viral infection of marine cyanobacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 15590–15595. [Google Scholar] [CrossRef] [Green Version]
- Fridman, S.; Flores-Uribe, J.; Larom, S.; Alalouf, O.; Liran, O.; Yacoby, I.; Béjà, O. A myovirus encoding both photosystem I and II proteins enhances cyclic electron flow in infected Prochlorococcus cells. Nat. Microbiol. 2017, 2, 1350–1357. [Google Scholar] [CrossRef]
- Wei, Y.; Xia, X. Unique Shine-Dalgarno Sequences in Cyanobacteria and Chloroplasts Reveal Evolutionary Differences in Their Translation Initiation. Genome Biol. Evol. 2019, 11, 3194–3206. [Google Scholar] [CrossRef] [PubMed]
- Šulčius, S.; Šimoliūnas, E.; Alzbutas, G.; Gasiūnas, G.; Jauniškis, V.; Kuznecova, J.; Miettinen, S.; Nilsson, E.; Meškys, R.; Roine, E.; et al. Genomic Characterization of Cyanophage vB_AphaS-CL131 Infecting Filamentous Diazotrophic Cyanobacterium Aphanizomenon flos-aquae Reveals Novel Insights into Virus-Bacterium Interactions. Appl. Environ. Microbiol. 2019, 85, e01311-18. [Google Scholar]
- Maccoll, R. Allophycocyanin and energy transfer. Biochim. Biophys. Acta (BBA)-Bioenerg. 2004, 1657, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Zhang, J.P.; Chang, W.R.; Liang, D.C. Crystal structure of R-phycocyanin and possible energy transfer pathways in the phycobilisome. Biophys. J. 2001, 81, 1171–1179. [Google Scholar] [CrossRef] [Green Version]
- Thornton, L.E.; Ohkawa, H.; Roose, J.L.; Kashino, Y.; Keren, N.; Pakrasi, H.B. Homologs of plant PsbP and PsbQ proteins are necessary for regulation of photosystem ii activity in the cyanobacterium Synechocystis 6803. Plant Cell 2004, 16, 2164–2175. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.X.; Schröder, W.P. The low molecular mass subunits of the photosynthetic supracomplex, photosystem II. Biochim. Biophys Acta 2004, 1608, 75–96. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, N.; Shen, J.R. Crystal structure of oxygen-evolving photosystem II from Thermosynechococcus vulcanus at 3.7-A resolution. Proc. Natl. Acad. Sci. USA 2003, 100, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Raymond, J.; Blankenship, R.E. The evolutionary development of the protein complement of photosystem 2. Biochim. Biophys. Acta (BBA)-Bioenerg. 2004, 1655, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Xiao, Y.; Pi, X.; Zhao, L.; Zhu, Q.; Wang, W.; Kuang, T.; Han, G.; Sui, S.; Shen, J. Supplementary Information for Structural insights into a dimeric Psb27-photosystem II complex from a cyanobacterium Thermosynechococcus vulcanus.pdf. Proc. Natl. Acad. Sci. USA 2021, 118, e2018053118. [Google Scholar] [CrossRef]
- Webber, A.N.; Malkin, R. Photosystem I reaction-centre proteins contain leucine zipper motifs. A proposed role in dimer formation. FEBS Lett. 1990, 264, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Panavas, T.; Pikula, A.; Reid, P.D.; Rubinstein, B.; Walker, E.L. Identification of senescence-associated genes from daylily petals. Plant Mol. Biol. 1999, 40, 237–248. [Google Scholar] [CrossRef]
- Watson, H.C.; Walker, N.P.; Shaw, P.J.; Bryant, T.N.; Wendell, P.L.; Fothergill, L.A.; Perkins, R.E.; Conroy, S.C.; Dobson, M.J.; Tuite, M.F. Sequence and structure of yeast phosphoglycerate kinase. EMBO J. 1982, 1, 1635–1640. [Google Scholar] [CrossRef]
- Tarnawski, M.; Krzywda, S.; Bialek, W.; Jaskolski, M.; Szxzepaniak, A. Structure of the RuBisCO chaperone RbcX from the thermophilic cyanobacterium Thermosynechococcus elongatus. Acta Cryst. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 851–857. [Google Scholar] [CrossRef] [Green Version]
- Battchikova, N.; Zhang, P.; Rudd, S.; Ogawa, T.; Aro, E.M. Identification of NdhL and Ssl1690 (NdhO) in NDH-1L and NDH-1M complexes of Synechocystis sp. PCC 6803. J. Biol. Chem. 2005, 280, 2587–2595. [Google Scholar] [CrossRef] [Green Version]
- Rumeau, D.; Bécuwe-Linka, N.; Beyly, A.; Louwagie, M.; Garin, J.; Peltier, G. New subunits NDH-M, -N, and -O, encoded by nuclear genes, are essential for plastid Ndh complex functioning in higher plants. Plant Cell 2005, 17, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Battchikova, N.; Paakkarinen, V.; Katoh, H.; Iwai, M.; Ikeuchi, M.; Pakrasi, H.; Ogawa, T.; Aro, E.M. Isolation, subunit composition and interaction of the NDH-1 complexes from Thermosynechococcus elongatus BP-1. Biochem. J. 2005, 390, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Wilkens, S.; Zhang, Z.; Zheng, Y. A structural model of the vacuolar ATPase from transmission electron microscopy. Micron 2005, 36, 109–126. [Google Scholar] [CrossRef]
- Leyva, J.; Bianchet, M.A.; Amzel, L.M. Understanding ATP synthesis: Structure and mechanism of the F1-ATPase (Review). Mol. Membr. Biol. 2003, 20, 27–33. [Google Scholar] [CrossRef]
- Mattevi, A.; Obmolova, G.; Kalk, K.H.; Westphal, A.H.; Kok, A.D.; Hol, W.G. Refined Crystal Structure of the Catalytic Domain of Dihydrolipoyl Transacetylase (E2p) from Azotobacter vinelandii at 2·6 Å Resolution. J. Mol. Biol. 1993, 230, 1183–1199. [Google Scholar] [CrossRef]
- Maguire, B.A.; Zimmermann, R.A. The Ribosome in Focus. Cell 2001, 104, 813–816. [Google Scholar] [CrossRef]
- Sanyal, S.C.; Liljas, A. The end of the beginning: Structural studies of ribosomal proteins. Curr. Opin. Struct. Biol. 2000, 10, 633–636. [Google Scholar] [CrossRef]
- Chan, Y.L.; Suzuki, K.; Olvera, J.; Wool, I.G. Zinc finger-like motifs in rat ribosomal proteins S27 and S29. Nucleic Acids Res. 1993, 21, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Hertveldt, K.; Lavigne, R.; Pleteneva, E.; Sernova, N.; Kurochkina, L.; Korchevskii, R.; Robben, J.; Mesyanzhinov, V.; Krylov, V.N.; Volckaert, G. Genome comparison of pseudomonas aeruginosa large phages. J. Mol. Biol. 2005, 354, 536–545. [Google Scholar] [CrossRef]
- Ma, Y.; Allen, L.Z.; Palenik, B. Diversity and genome dynamics of marine cyanophages using metagenomic analyses. Environ. Microbiol. Rep. 2015, 6, 583–594. [Google Scholar] [CrossRef]
- Hevroni, G.; Enav, H.; Rohwer, F.; Beja, O. Diversity of viral photosystem-I psaA genes. ISME J. 2015, 9, 1892–1898. [Google Scholar] [CrossRef]
- Lemay, M.L.; Otto, A.; Maa, S.; Plate, K.; Becher, D.; Moineau, S. Investigating Lactococcus lactis MG1363 Response to Phage p2 Infection at the Proteome Level. Mol. Cell. Proteom. 2019, 18, 704–714. [Google Scholar] [CrossRef] [Green Version]
- Gao, E.; Huang, Y.; Ning, D. Metabolic Genes within Cyanophage Genomes: Implications for Diversity and Evolution. Genes 2016, 7, 80. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orders | Family | Species | Strains | Susceptibility | Origin |
|---|---|---|---|---|---|
| Chroococcales | Microcystaceae | Microcystis aeruginosa | FACHB-905 | + | China |
| FACHB-925 | − | Australia | |||
| FACHB-942 | − | China | |||
| FACHB-469 | + | France | |||
| FACHB-924 | − | Australia | |||
| FACHB-1326 | − | China | |||
| FACHB-912 | − | China | |||
| M. wesenbergii | FACHB-908 | + | China | ||
| FACHB-1112 | − | China | |||
| FACHB-1317 | − | China | |||
| FACHB-1318 | − | China | |||
| FACHB-929 | + | Japan | |||
| M. viridis | FACHB-979 | + | Japan | ||
| M. Flos-aquae | FACHB-1028 | + | China | ||
| Microcystis sp. | FACHB-915 | + | France | ||
| M. elabens | FACHB-916 | − | Japan | ||
| M. Panniformis | FACHB-1757 | − | China | ||
| M. ichthyoblabe Kutz | FACHB-1294 | − | China | ||
| FACHB-1409 | + | China | |||
| Chroococcacaea | Chroococcus sp. | FACHB-193 | + | China | |
| Nostocales | Aphanizomenonaceae | Aphanizomenon flos-aquae | FACHB-1039 | − | China |
| FACHB-1040 | + | China | |||
| Anabaena flos-aquae | FACHB-245 | + | USA | ||
| Anabaena sp. | FACHB-418 | + | France | ||
| Dolichospermum flos-aquae | FACHB-1255 | + | China | ||
| Nostocaceae | Nostoc sp. | FACHB-596 | + | China | |
| Oscillatoriale | Microcoleaceae | Planktothrix agardhii | FACHB-1166 | − | China |
| FACHB-920 | + | Japan | |||
| Planktothricoides raciborskii | FACHB-881 | + | China | ||
| Oscillatoriaceae | Oscillatoria planctonica | FACHB-708 | + | China | |
| Hormogonales | Scytonemataceae | Plectonema | FACHB-402 | − | USA |
| FACHB-240 | − | USA | |||
| Synechococcales | Synechococcaceae | Synechococcus sp. | FACHB-805 | + | Australia |
| Accession | Protein Name | p Value | Adj. p-Value | T1:C1 | T8:C8 |
|---|---|---|---|---|---|
| Q8YXF2 | ATP-dependent zinc metalloproteinase FtsH | 0.008218777 | 0.52091175 | 1.5848247↑ | |
| Q8YPZ8 | All4042 protein | 0.020302562 | 0.55631214 | 1.4674468↑ | |
| A0A6P0S0D5 | 30S ribosomal protein S14 | 0.014660408 | 0.53405772 | 1.4211440↑ | |
| Q8YT39 | Alr2887 protein | 0.005383913 | 0.45763262 | 0.7740167↓ | |
| Q8YTI1 | All2736 protein | 0.001327486 | 0.16925451 | 0.7467759↓ | |
| A0A1Z4KLJ5 | 50Sribosomal protein L23 | 0.01222975 | 0.52091175 | 0.3643173↓ | |
| A0A1Z4M5R2 | Photosystem I P700 chlorophyll-a apolipoprotein A1 | 0.000000967 | 0.0000825 | 3.42825728↑ | |
| Q8YYI1 | Carbon Dioxide Concentration Mechanism protein | 0.000305335 | 0.00390828 | 2.98863046↑ | |
| Q8YTI1 | All2736 protein | 0.000000887 | 0.0000825 | 2.26210112↑ | |
| A0A1Z4IAM0 | Unknown functional protein | 0.000000195 | 0.0000499 | 0.33966838↓ | |
| A0A1Z4KN79 | Argininosuccinate synthase | 0.030938777 | 0.07920326 | 0.28915421↓ | |
| Q8YWR3 | Peroxidase 2 family of proteins/glutaminase | 0.009777517 | 0.04120970 | 0.25880121↓ |
| KEGG Pathway | Protein Name | Protein Description | Upward/Downward |
|---|---|---|---|
| Photosynthesis-antennal proteins | ApcB | Allophycocyanin β subunit | Down |
| CpcB | Phycocyanin associated rod junction protein | Up | |
| Ribosomal | L23 | Ribosomal large subunit protein L23 | Down |
| L2 | Ribosomal large subunit protein L2 | Down | |
| S14 | Ribosomal small subunit protein S14 | Up | |
| L35 | Ribosomal large subunit protein L35 | Up | |
| L3 | Ribosomal large subunit protein L3 | Down | |
| L4 | Ribosomal large subunit protein L4 | Down | |
| L22 | Ribosomal large subunit protein L22 | Down | |
| L29 | Ribosomal large subunit protein L29 | Up | |
| L5 | Ribosomal large subunit protein L5 | Down | |
| S5 | Ribosomal small subunit protein S5 | Up | |
| L13 | Ribosomal large subunit protein L13 | Up | |
| L1 | Ribosomal large subunit protein L1 | Up | |
| L35 | Ribosomal large subunit protein L35 | Up | |
| S6 | Ribosomal small subunit protein S6 | Down | |
| L21 | Ribosomal large subunit protein L21 | Up | |
| S16 | Ribosomal small subunit protein S16 | Up | |
| S1 | Ribosomal small subunit protein S1 | Up | |
| Oxidative phosphorylation | NdhK | NAD (P) H-quinone oxidoreductase subunit K(EC:7.1.1.2) | Down |
| alpha | F-type H+/Na+ transporter ATPase subunit α (EC:7.1.2.2 7.2.2.1) | Up | |
| b | ATPase B subunit | Up | |
| OSCP | F type H~+ transport ATPase subunit | Up | |
| Photosynthesis | Psb-A | Photosystem II P680 reaction center D1 protein (EC:1.10.3.9) | Down |
| Psb-O | Photosystem II oxygen evolution enhancer protein 1 | Down | |
| Psb-U | Photosystem IIPsbU protein | Down | |
| Psb-V | Photosystem II cytochrome C550 | Down | |
| Psb27 | Photosystem IIPsb27 protein | Up | |
| PsaA | Photosystem I P700 chlorophyll-a apolipoprotein A1 | Up | |
| PetA | Acocytochrome F | Up | |
| PetE | plastocyanin | Down | |
| alpha | F-type H+/Na+ transporter ATPase subunit α (EC:7.1.2.2 7.2.2.1) | Up | |
| b | ATPase B subunit | Up | |
| Carbon fixation | 2.7.9.1 | Pyruvate, orthophosphate dikinase | Down |
| 4.1.1.39 | Ribulose bisphosphate carboxylase large chain | Down | |
| 2.7.2.3 | phosphoglyceric kinase | Down | |
| 2.7.9.1 | Pyruvate, orthophosphate dikinase | Down | |
| Glycolysis and gluconeogenesis | 2.7.2.3 | phosphoglyceric kinase | Down |
| 2.3.1.12 | Pyruvate dehydrogenase E2 component (dihydrolipamidoacetyltransferase) | Up |
| No. | Location | Description | Score | Coverage |
|---|---|---|---|---|
| 1 | 20651..21748 | putative major capsid protein (Nostoc phage A1) | 10,458 | 86% |
| 2 | 23869..25389 | tail sheath protein (Nostoc phage A1) | 4372 | 46% |
| 3 | 57421..58554 | tail collar protein (Nostoc phage A1) | 2265 | 24% |
| 4 | 25487..25996 | tail tube protein (Nostoc phage A1) | 2101 | 91% |
| 5 | 18162..19766 | hypothetical protein (Nostoc phage A1) | 1811 | 62% |
| 6 | 29345..32047 | lysozyme (Anabaena phage Elbi) | 1151 | 30% |
| 7 | 5775..7658 | alkaline phosphatase D family protein (Salinivenus lutea) | 1132 | 35% |
| 8 | 26714..28780 | tail protein (Nostoc phage A1) | 1119 | 34% |
| 9 | 60572..61747 | baseplate J tail protein (Nostoc phage N1) | 1048 | 44% |
| 10 | 5256..5762 | Capsid fiber protein | 1037 | 61% |
| 11 | 58577..59965 | tail fiber protein (Nostoc phage N1) | 995 | 42% |
| 12 | 21893..22378 | hypothetical protein (Nostoc phage A1) | 925 | 49% |
| 13 | 19898..20644 | putative outer membrane protein (Nostoc phage A1) | 912 | 49% |
| 14 | 61784..62131 | lysozyme (Nostoc phage N1) | 672 | 73% |
| 15 | 4580..5233 | hypothetical protein (Nostoc phage A1) | 453 | 24% |
| 16 | 32813..33661 | hypothetical protein (Nostoc phage N1) | 376 | 37% |
| 17 | 8358..9086 | hypothetical protein (Nostoc phage A1) | 288 | 20% |
| 18 | 59997..60572 | tail collar protein (Nostoc phage A1) | 198 | 30% |
| 19 | 23110..23832 | hypothetical protein (Nostoc phage A1) | 169 | 25% |
| 20 | 7738..8361 | hypothetical protein (Nostoc phage A1) | 156 | 40% |
| 21 | 62202..62531 | hypothetical protein | 101 | 64% |
| 22 | 32074..32541 | hypothetical protein | 100 | 26% |
| 23 | 63660..64472 | Baseplate structural protein | 71 | 11% |
| 24 | 22375..22701 | putative tail-component | 66 | 19% |
| 25 | 22706..23113 | hypothetical protein | 57 | 18% |
| 26 | 15042..15680 | hypothetical protein (Nostoc phage A1) | 29 | 6% |
| 27 | 42619..43221 | hypothetical protein (Microcystis phage Me-ZS1) | 29 | 6% |
| 28 | 55094..57100 | DNA polymerase delta catalytic subunit | 19 | 1% |
| 29 | 3..722 | Thymidylate kinase (EC 2.7.4.9) | 19 | 7% |
| 30 | 20661..26102 | tail length tape-measure protein (Microcystis phage Me-ZS1) | 16 | 2% |
| 31 | 41382..41699 | hypothetical protein (Microcystis phage Me-ZS1) | 14 | 6% |
| 32 | 26900..27886 | tail tube protein (Microcystis phage Me-ZS1) | 13 | 4% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Zhao, B.; Li, J.; Song, X.; Tong, Y.; An, W. Host Cyanobacteria Killing by Novel Lytic Cyanophage YongM: A Protein Profiling Analysis. Microorganisms 2022, 10, 257. https://doi.org/10.3390/microorganisms10020257
Zhang S, Zhao B, Li J, Song X, Tong Y, An W. Host Cyanobacteria Killing by Novel Lytic Cyanophage YongM: A Protein Profiling Analysis. Microorganisms. 2022; 10(2):257. https://doi.org/10.3390/microorganisms10020257
Chicago/Turabian StyleZhang, Shanshan, Baohua Zhao, Jing Li, Xiaofei Song, Yigang Tong, and Wenlin An. 2022. "Host Cyanobacteria Killing by Novel Lytic Cyanophage YongM: A Protein Profiling Analysis" Microorganisms 10, no. 2: 257. https://doi.org/10.3390/microorganisms10020257