
Role of the Vibriolysin VemA Secreted by the Emergent Pathogen Vibrio europaeus in the Colonization of Manila Clam Mucus
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Media
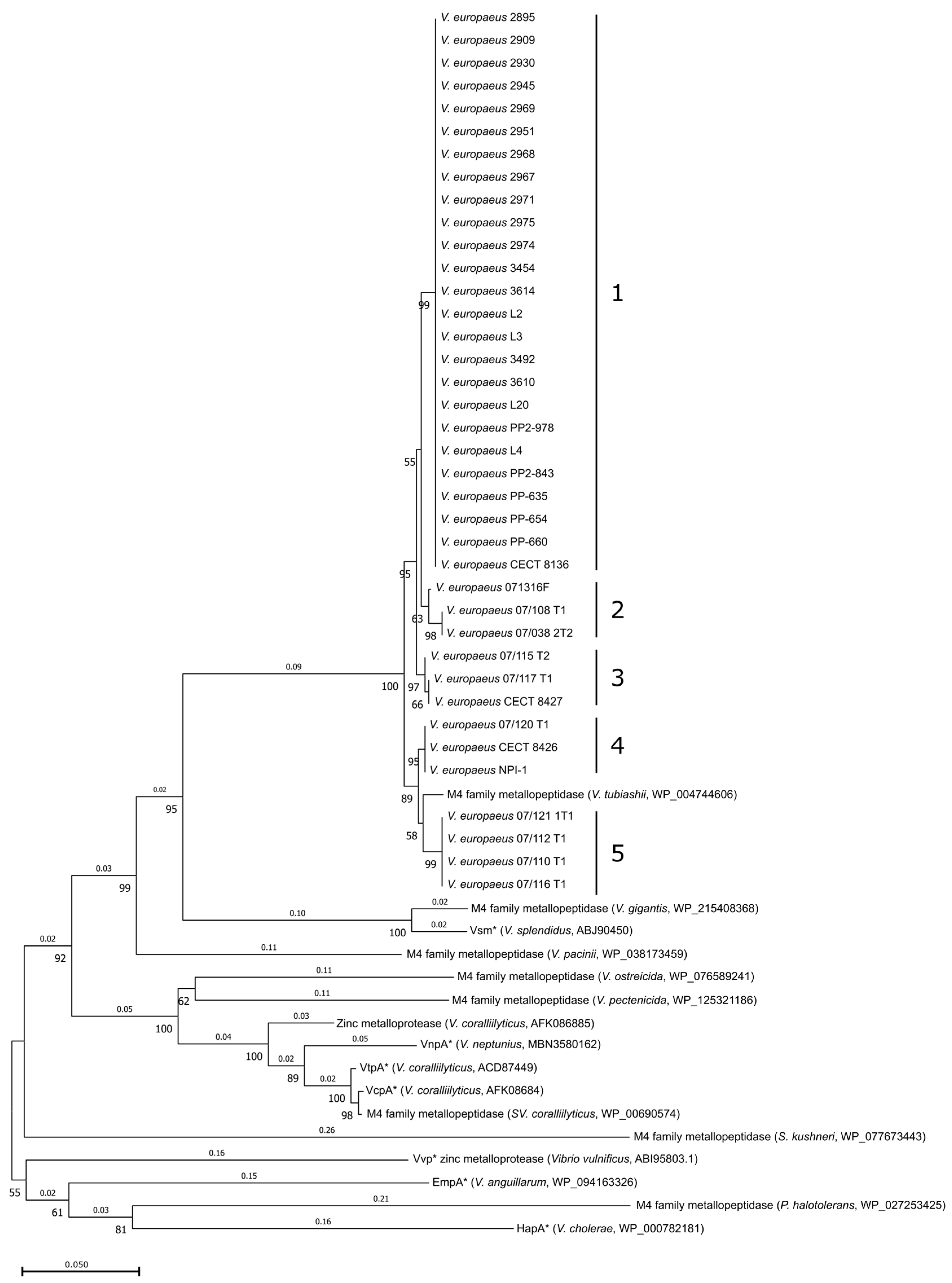
2.2. Search of VemA Homologous and Phylogenetic Analysis
2.3. Construction of ΔvemA Mutants by Double Allelic Exchange
2.4. Virulence Challenges
2.5. Mucus Extraction
2.6. Evaluation of the Bactericidal Activity of the Surface Mucus on Solid Media
2.7. Adherence to Mucus and Biofilm Formation on Polystyrene
2.8. Chemotaxis Assays
2.9. Bacterial Survival and Growth with Mucus
2.10. Statistical Analyses
3. Results
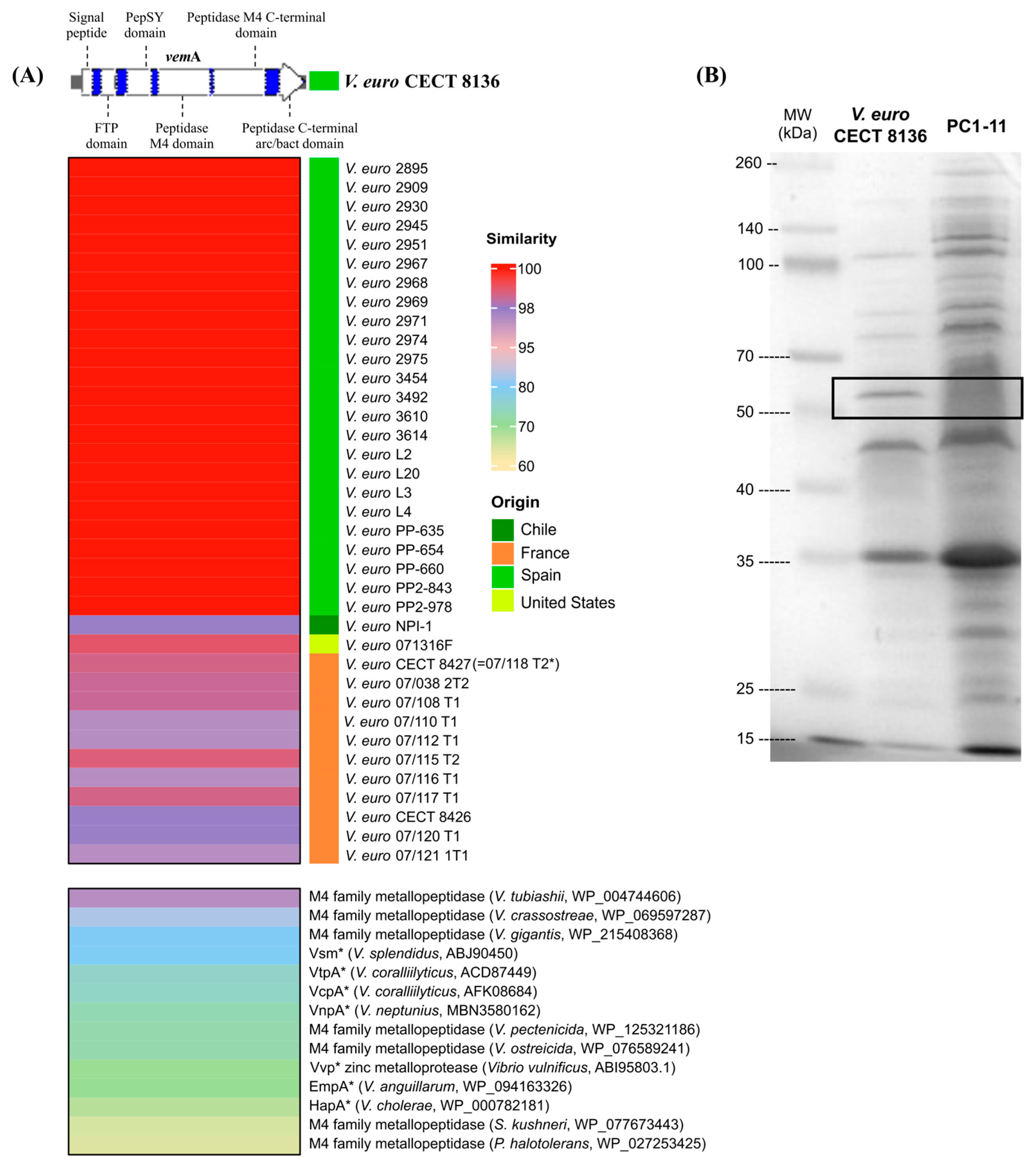
3.1. VemA Is a Vibriolysin-like Protein, Highly Conserved in V. europaeus with Homologues in Other Pathogenic Vibrio Species
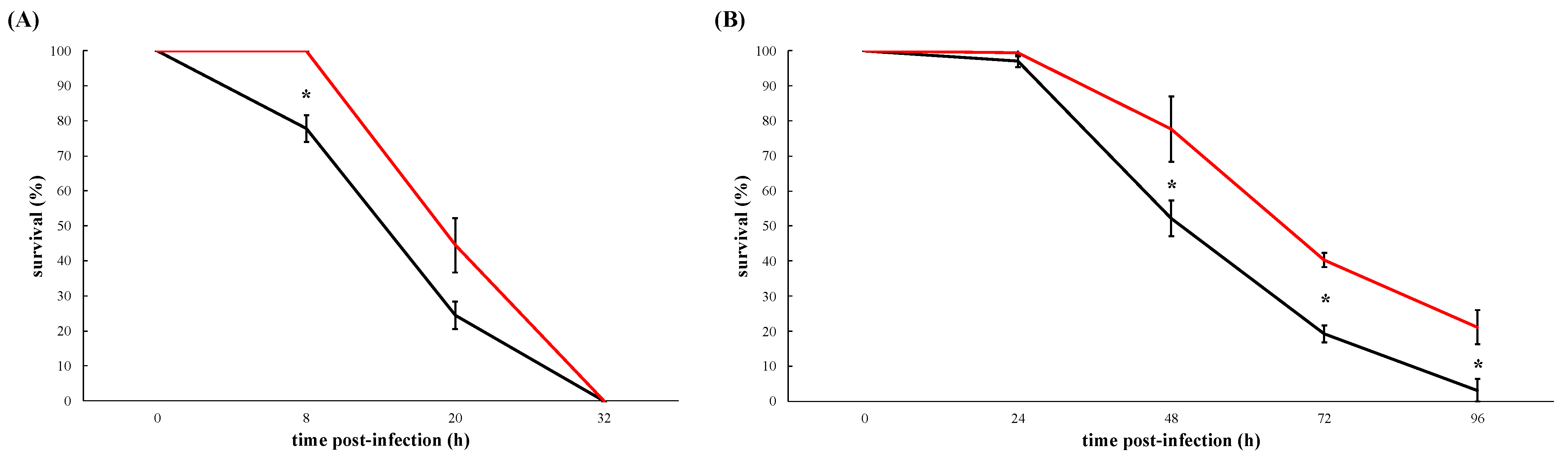
3.2. VemA Is an Extracellular Protein Necessary for the Full Virulence of V. europaeus Although It Is Not a Major Virulence Factor
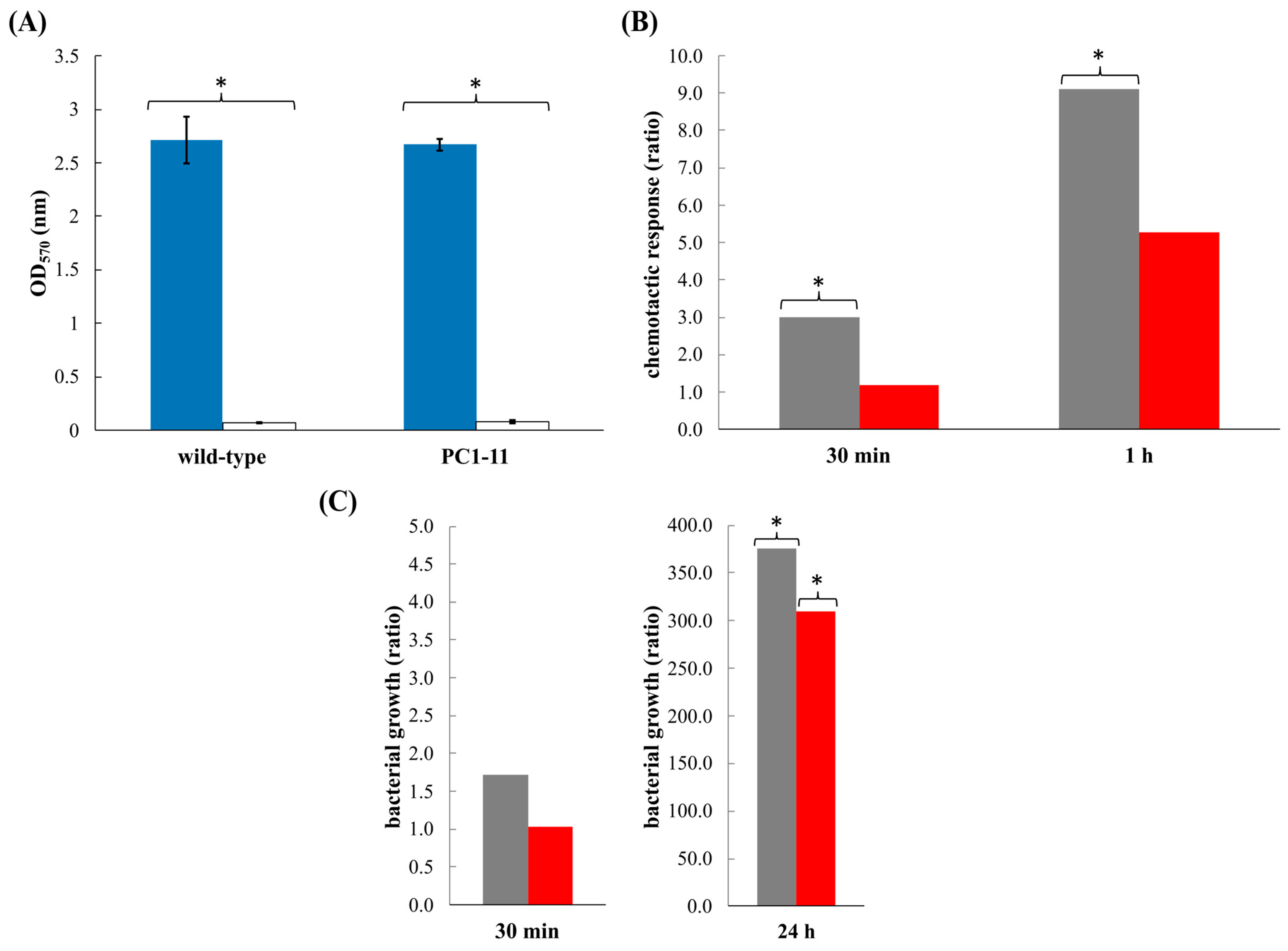
3.3. V. europaeus Forms Biofilms on Manila Clam Mucus
3.4. V. europaeus Is Attracted by Mucus and VemA Participates in Bacterial Chemotaxis
3.5. VemA Promotes the Proliferation of V. europaeus over the Mucus Matrix
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Brousseau, D.J.; Braun, P.C.; Harper-Leatherman, A.S.; Sullivan, E.; Baglivo, J.A. Antimicrobial activity in the pallial cavity fluids of the oyster Crassostrea virginica (Gmelin) from a highly impacted harbor in western long island sound. J. Shellfish Res. 2014, 33, 719–725. [Google Scholar] [CrossRef]
- Braun, P.C.; Brousseau, D.J.; Lecleir, G.R. Microbial inhibition by bacteria isolated from pallial cavity fluids and associated mucus of the Eastern oyster Crassostrea virginica (Gmelin). J. Shellfish Res. 2019, 33, 565–572. [Google Scholar] [CrossRef]
- Dubert, J.; Barja, J.L.; Romalde, J.L. New insights into pathogenic vibrios affecting bivalves in hatcheries: Present and future prospects. Front. Microbiol. 2017, 8, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanella, C.; Mosca, F.; Mariani, F.; Franci, G.; Folliero, V.; Galdiero, M.; Tiscar, P.G.; Galdiero, M. Microbial diseases of bivalve mollusks: Infections, immunology and antimicrobial defense. Mar. Drugs 2017, 15, 182. [Google Scholar] [CrossRef]
- Prado, S.; Romalde, J.L.; Montes, J.; Barja, J.L. Pathogenic bacteria isolated from disease outbreaks in shellfish hatcheries. First description of Vibrio neptunius as an oyster pathogen. Dis. Aquat. Organ. 2005, 67, 209–215. [Google Scholar]
- Saulnier, D.; De Decker, S.; Haffner, P.; Cobret, L.; Robert, M.; Garcia, C. A large-scale epidemiological study to identify bacteria pathogenic to Pacific Oyster Crassostrea gigas and correlation between virulence and metalloprotease-like activity. Microb. Ecol. 2010, 59, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Travers, M.A.; Mersni-Achour, R.; Haffner, P.; Tourbiez, D.; Cassone, A.L.; Morga, B.; Doghri, I.; Garcia, C.; Renault, T.; Fruitier-Arnaudin, I.; et al. First description of French, V. tubiashii strains pathogenic to mollusk: I. Characterization of isolates and detection during mortality events. J. Invertebr. Pathol. 2014, 123, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Prado, S.; Dubert, J.; Barja, J.L. Characterization of pathogenic vibrios isolated from bivalve hatcheries in Galicia, NW Atlantic coast of Spain. Description of Vibrio tubiashii subsp. europaensis subsp. nov. Syst. Appl. Microbiol. 2015, 38, 26–29. [Google Scholar] [CrossRef]
- Dubert, J.; Aranda-Burgos, J.A.; Ojea, J.; Barja, J.L.; Prado, S. Mortality event involving larvae of the carpet shell clam Ruditapes decussatus in a hatchery: Isolation of the pathogen Vibrio tubiashii subsp. europaeus. J. Fish Dis. 2017, 9, 1185–1193. [Google Scholar] [CrossRef]
- Rojas, R.; Blanco-Hortas, A.; Kehlet-Delgado, H.; Lema, A.; Miranda, C.D.; Romero, J.; Martinez, P.; Barja, J.L.; Dubert, J. First description outside Europe of the emergent pathogen Vibrio europaeus in shellfish aquaculture. J. Invertebr. Pathol. 2021, 180, 107542. [Google Scholar] [CrossRef] [PubMed]
- Mersni-Achour, R.; Imbert-Auvray, N.; Huet, V.; Cheikh, Y.B.; Faury, N.; Doghri, I.; Rouatbi, S.; Bordenave, S.; Travers, M.A.; Saulnier, D.; et al. First description of French, V. tubiashii strains pathogenic to mollusk: II. Characterization of properties of the proteolytic fraction of extracellular products. J. Invertebr. Pathol. 2014, 123, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mersni-Achour, R.; Cheikh, Y.B.; Pichereau, V.; Doghri, I.; Etien, C.; Dégremont, L.; Saulnier, D.; Fruitier-Arnaudin, I.; Travers, M.A. Factors other than metalloprotease are required for full virulence of French Vibrio tubiashii isolates in oyster larvae. Microbiology 2015, 161, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Dubert, J.; Romalde, J.L.; Spinard, E.J.; Nelson, D.R.; Gomez-Chiarri, M.; Barja, J.L. Reclassification of the larval pathogen for marine bivalves Vibrio tubiashii subsp. europaeus as Vibrio europaeus sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 4791–4796. [Google Scholar] [CrossRef]
- Lodeiros, C.; Bolinches, J.; Dopazo, C.P.; Toranzo, A.E. Bacillary necrosis in hatcheries of Ostrea edulis in Spain. Aquaculture 1987, 65, 15–29. [Google Scholar] [CrossRef]
- Dubert, J.; da Costa, F.; Aranda-Burgos, J.A.; Martínez-Patiño, D.; Prado, S.; Barja, J.L. Beneficial effects of carpet shell clam (Ruditapes decussatus) depuration during short periods of conditioning in shellfish hatchery: Role of the temperature and phytoplankton on reduction and diversity of vibrios. Aquaculture 2016, 459, 65–72. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.A. Virulence factors and their mechanisms of action: The view from a damage-response framework. J. Water Health 2009, 7, S2–S18. [Google Scholar] [CrossRef]
- Spinard, E.J.; Dubert, J.; Nelson, D.R.; Gomez-Chiarri, M.; Barja, J.L. Draft genome sequence of the emerging bivalve pathogen Vibrio tubiashii subsp. europaeus. Genome Announc. 2016, 4, e00625-16. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, H.; Lind, E.J.; Boin, M.A.; Häse, C.C. The extracellular metalloprotease of Vibrio tubiashii is a major virulence factor for Pacific Oyster (Crassostrea gigas) larvae. Appl. Environ. Microbiol. 2008, 74, 4101–4110. [Google Scholar] [CrossRef] [Green Version]
- Sussman, M.; Mieog, J.C.; Doyle, J.; Victor, S.; Willis, B.L.; Bourne, D.G. Vibrio zinc-metalloprotease causes photoinactivation of coral endosymbionts and coral tissue lesions. PLoS ONE 2009, 4, e4511. [Google Scholar] [CrossRef] [Green Version]
- Labreuche, Y.; Le Roux, F.; Henry, J.; Zatylny, C.; Huvet, A.; Lambert, C.; Soudant, P.; Mazel, D.; Nicolas, J.L. Vibrio aestuarianus zinc metalloprotease causes lethality in the Pacific oyster Ostrea edulis and impairs the host cellular immune defenses. Fish Shellfish Immunol. 2010, 29, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Galvis, F.; Barja, J.L.; Lemos, M.L.; Balado, M. The vibriolysin-like protease VnpA and the vollagenase ColA are required for full virulence of the bivalve mollusks pathogen Vibrio neptunius. Antibiotics 2021, 10, 391. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, F.; Binesse, J.; Saulnier, D.; Mazel, D. Construction of a Vibrio splendidus mutant lacking the metalloprotease gene vsm by use of a novel counterselectable suicide vector. Appl. Environ. Microbiol. 2006, 73, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaz-Hidalgo, R.; Cleenwerck, I.; Balboa, S.; Prado, S.; De Vos, P.; Romalde, J.L. Vibrio breoganii sp. nov., a non-motile, alginolytic, marine bacterium within the Vibrio halioticoli clade. Int. J. Syst. Evol. Microbiol. 2009, 59, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Val, M.E.; Skovgaard, O.; Ducos-Galand, M.; Bland, M.J.; Mazel, D. Genome engineering in Vibrio cholerae: A feasible approach to address biological issues. PLoS Genet. 2012, 8, e1002472. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Hussain, F.A.; Dubert, J.; Elsherbini, J.; Murphy, M.; Vaninsberghe, D.; Arevalo, P.; Kauffman, K.; Rodino-Janeiro, B.K.; Gavin, H.; Gomez, A.; et al. Rapid evolutionary turnover of mobile genetic elements drives bacterial resistance to phages. Science 2021, 374, 488–492. [Google Scholar] [CrossRef]
- Dubert, J.; Romalde, J.L.; Prado, S.; Barja, J.L. Vibrio bivalvicida sp. nov., a novel larval pathogen for bivalve molluscs reared in a hatchery. Syst. Appl. Microbiol. 2015, 39, 8–13. [Google Scholar] [CrossRef]
- Terceti, M.S.; Vences, A.; Matanza, X.M.; Barca, A.V.; Noia, M.; Lisboa, J.; Dos Santos, N.M.S.; do Vale, A.; Osorio, C.R. The RstAB system impacts virulence, motility, cell morphology, penicillin tolerance and production of type II secretion system-dependent factors in the fish and human pathogen Photobacterium damselae subsp. damselae. Front. Microbiol. 2019, 10, 897. [Google Scholar] [CrossRef] [PubMed]
- Valiente, E.; Lee, C.T.; Hor, L.I.; Fouz, B.; Amaro, C. Role of the metalloprotease Vvp and the virulence plasmid pR99 of Vibrio vulnificus serovar E in surface colonization and fish virulence. Environ. Microbiol. 2008, 10, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.F.; Zeng, B.Q.; Liu, D.; Wu, R.B.; Zhang, J.; Liao, B.Q.; He, H.L.; Bian, F. Classification and structural insight into vibriolysin-like proteases of Vibrio pathogenicity. Microb. Pathog. 2018, 117, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.K.; Kirn, H.Y.; Park, J.E.; Acharya, P.; Park, I.S.; Yoon, S.M.; You, H.J.; Hahm, K.S.; Park, J.K.; Lee, J.S. Vibrio vulnificus secretes a broad-specificity metalloprotease capable of interfering with blood homeostasis through prothrombin activation and fibrinolysis. J. Bacteriol. 2005, 187, 6909–6916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Liao, F.; Pang, B.; Fu, X.; Xu, W.; Kan, B.; Jing, H.; Gu, W. The complete genomic analysis of an imported Vibrio cholerae from Myanmar in southwest China. Infect. Genet. Evol. 2016, 44, 272–277. [Google Scholar] [CrossRef]
- Binesse, J.; Delsert, C.; Saulnier, D.; Champomier-Vergès, M.-C.; Zagorec, M.; Munier-Lehmann, H.; Mazel, D.; Le Roux, F. Metalloprotease Vsm is the major determinant of toxicity for extracellular products of Vibrio splendidus. Appl. Environ. Microbiol. 2008, 74, 7108–7117. [Google Scholar] [CrossRef] [Green Version]
- De O Santos, E.; Alves, N.; Dias, G.M.; Mazotto, A.M.; Vermelho, A.; Vora, G.J.; Wilson, B.; Beltran, V.H.; Bourne, D.G.; Le Roux, F.; et al. Genomic and proteomic analyses of the coral pathogen Vibrio coralliilyticus reveal a diverse virulence repertoire. ISME J. 2011, 5, 1471–1483. [Google Scholar] [CrossRef]
- Dubert, J.; Nelson, D.R.; Spinard, E.J.; Kessner, L.; Gomez-Chiarri, M.; DaCosta, F.; Prado, S.; Barja, J.L. Following the infection process of vibriosis in Manila clam (Ruditapes philippinarum) larvae through GFP-tagged pathogenic Vibrio species. J. Invertebr. Pathol. 2016, 133, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Denkin, S.M.; Nelson, D.R. Induction of protease activity in Vibrio anguillarum by gastrointestinal mucus. Appl. Environ. Microbiol. 1999, 65, 3555–3560. [Google Scholar] [CrossRef] [Green Version]
- Booth, B.A.; Boesman-Finkelstein, M.; Finkelstein, R.A. Vibrio cholerae soluble hemagglutinin/protease is a metalloenzyme. Infect. Immun. 1983, 42, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Hase, C.; Finkelstein, R.A. Bacterial extracellular zinc-containing metalloproteases. Microbiol. Rev. 1993, 57, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Benitez, J.A.; Silva, A.J. Vibrio cholerae hemagglutinin (HA)/protease: An extracellular metalloprotease with multiple pathogenic activities. Toxicon 2016, 115, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.; Garren, M.; Penn, K.; Fernandez, V.I.; Seymour, J.R.; Thompson, J.R.; Raina, J.B.; Stocker, R. Coral mucus rapidly induces chemokinesis and genome-wide transcriptional shifts toward early pathogenesis in a bacterial coral pathogen. ISME J. 2021, 15, 3668–3682. [Google Scholar] [CrossRef]
- Bisharat, N.; Cohen, D.I.; Maiden, M.C.; Crook, D.W.; Peto, T.; Harding, R.M. The evolution of genetic structure in the marine pathogen, Vibrio vulnificus. Infect. Genet. Evol. 2007, 7, 685–693. [Google Scholar] [CrossRef]
- Hasan, N.A.; Grim, C.J.; Lipp, E.K.; Rivera, I.N.G.; Chun, J.; Haley, B.J.; Taviani, E.; Choi, S.Y.; Hoq, M.; Munk, C.; et al. Deep-sea hydrothermal vent bacteria related to human pathogenic Vibrio species. Proc. Natl. Acad. Sci. USA 2015, 112, E2813–E2819. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wu, C.; Liu, D.; Yang, X.; Wu, R.; Zhang, J.; Ma, C.; He, H. C-terminal domains of bacterial proteases: Structure, function and the biotechnological applications. J. Appl. Microbiol. 2017, 122, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.F.; Wu, R.B.; Wu, C.L.; Liu, D.; Zhang, J.; Liao, B.Q.; Lei, M.; Xiao, X.; Ma, C.B.; He, H.L. New method of detecting hydrophobic interaction between C-terminal binding domain and bio-macromolecules. J. Biotechnol. 2018, 265, 101–108. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains and Plasmids | Description | Reference |
|---|---|---|
| CECT 8136 | V. europaeus wildtype strain | Dubert et al. [14] |
| PC1-11 | CECT 8136 ΔvemA | This study |
| C5.5 | Vibrio breoganii | Beaz-Hidalgo et al. [24] |
| E. coli Π3813 | B462 ΔthyA::(erm-pir-116) (EryR) | Le Roux et al. [23] |
| E. coli β3914 | β2163 gyrA462 zei-298::Tn10 (KnR EryR) | Le Roux et al. [23] |
| pSW7848T | pSW23T::araC-PBADccdB (CmR) | Val et al. [25] |
| pPC1 | pSW7848T flanked by two homologous regions upstream/downstream vemA | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez, C.; Rodriguez, S.; Vences, A.; Barja, J.L.; Toranzo, A.E.; Dubert, J. Role of the Vibriolysin VemA Secreted by the Emergent Pathogen Vibrio europaeus in the Colonization of Manila Clam Mucus. Microorganisms 2022, 10, 2475. https://doi.org/10.3390/microorganisms10122475
Martinez C, Rodriguez S, Vences A, Barja JL, Toranzo AE, Dubert J. Role of the Vibriolysin VemA Secreted by the Emergent Pathogen Vibrio europaeus in the Colonization of Manila Clam Mucus. Microorganisms. 2022; 10(12):2475. https://doi.org/10.3390/microorganisms10122475
Chicago/Turabian StyleMartinez, Clara, Sergio Rodriguez, Ana Vences, Juan L. Barja, Alicia E. Toranzo, and Javier Dubert. 2022. "Role of the Vibriolysin VemA Secreted by the Emergent Pathogen Vibrio europaeus in the Colonization of Manila Clam Mucus" Microorganisms 10, no. 12: 2475. https://doi.org/10.3390/microorganisms10122475