
The Endophytic Fungi Diversity, Community Structure, and Ecological Function Prediction of Sophora alopecuroides in Ningxia, China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research District’s General Situation
2.2. Samples Collection
2.3. Sample Pretreatment
2.4. DNA Extraction, Amplicon of ITS rDNA Region, and High-Throughput Sequencing
2.5. Processing of Sequencing Data
2.6. Statistical Analysis
3. Results
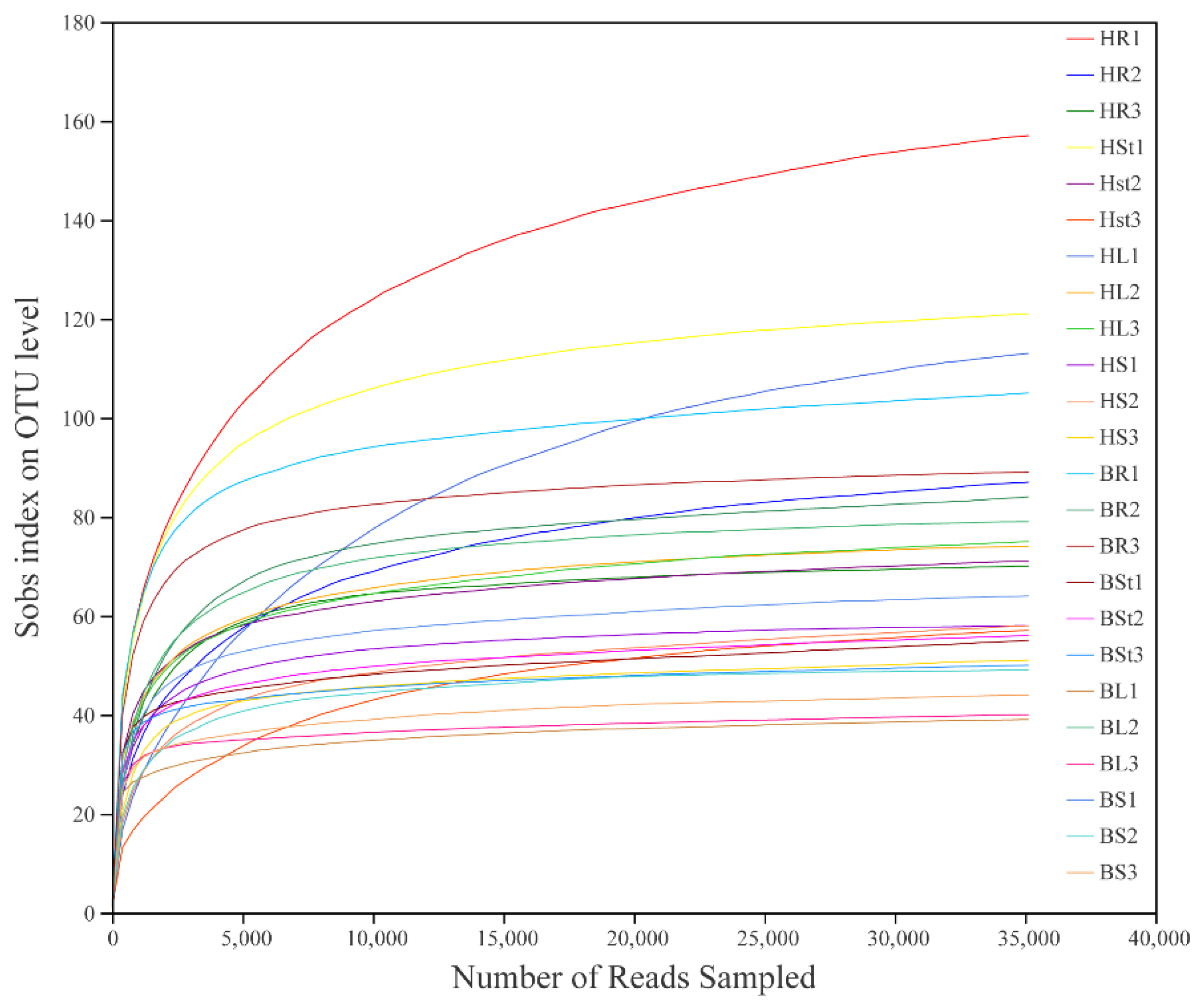
3.1. Overview of the Sequencing Data of Fungal Communities
3.2. Alpha Diversity of Endophytic Fungi of S. alopecuroides
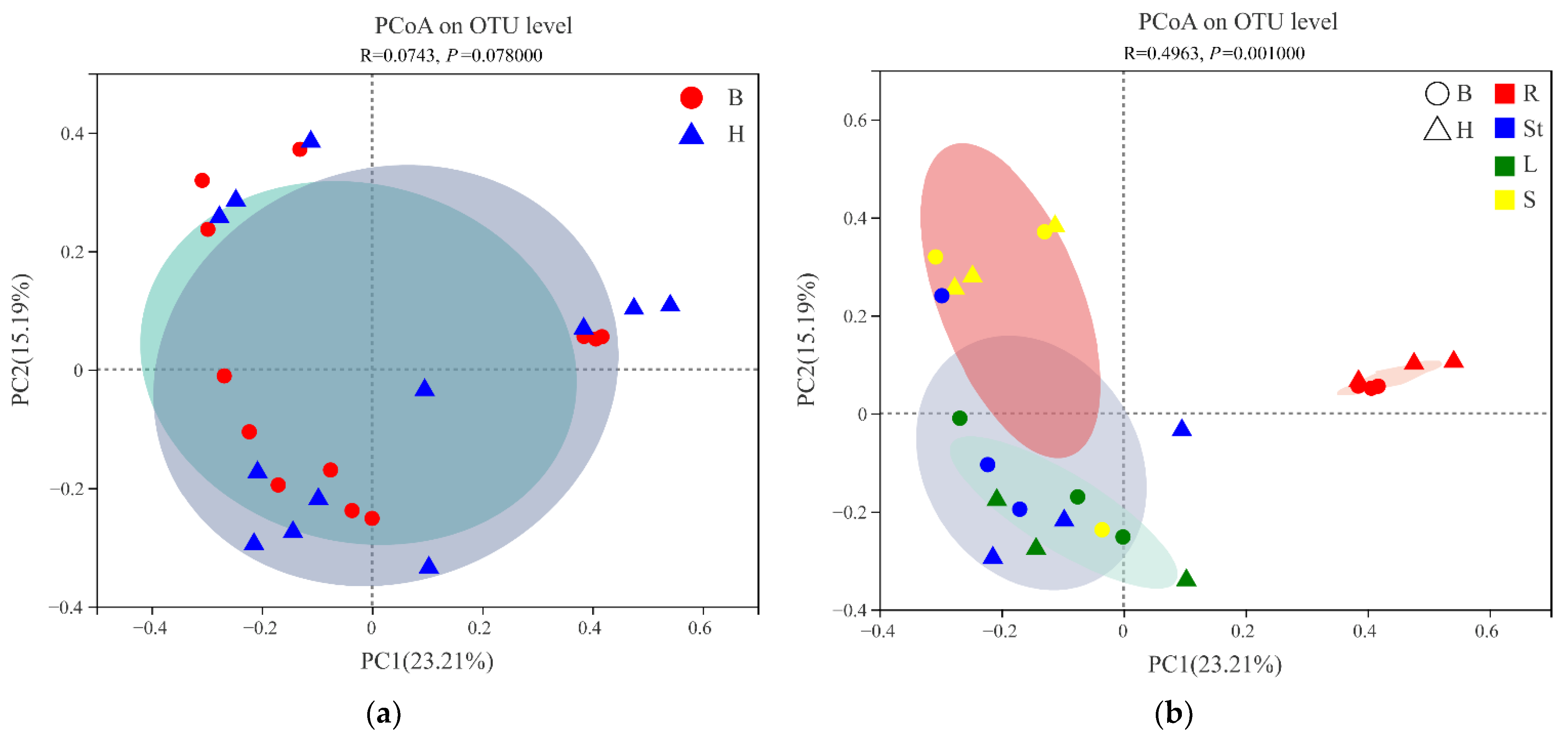
3.3. Beta Diversity of Endophytic Fungi of S. alopecuroides
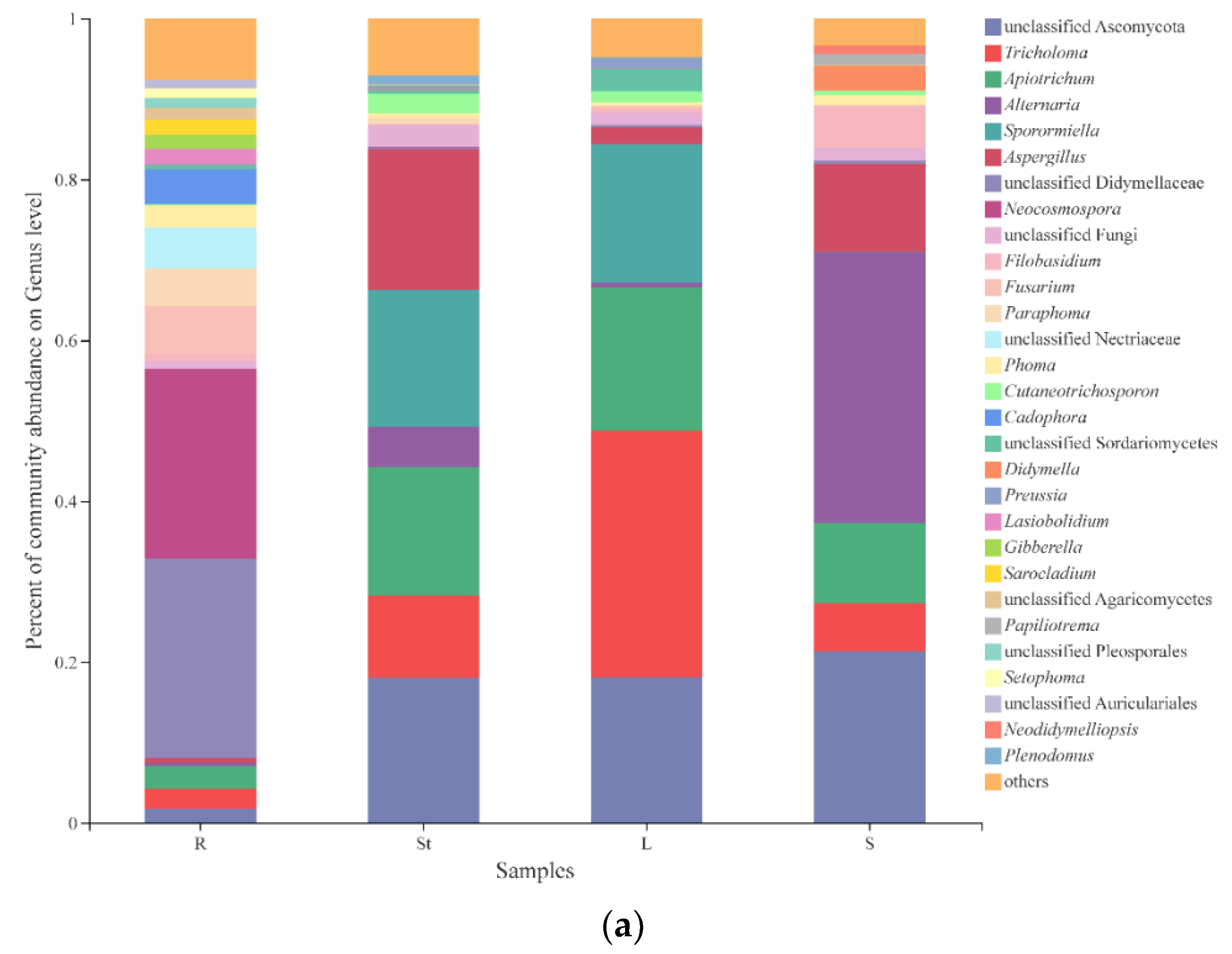
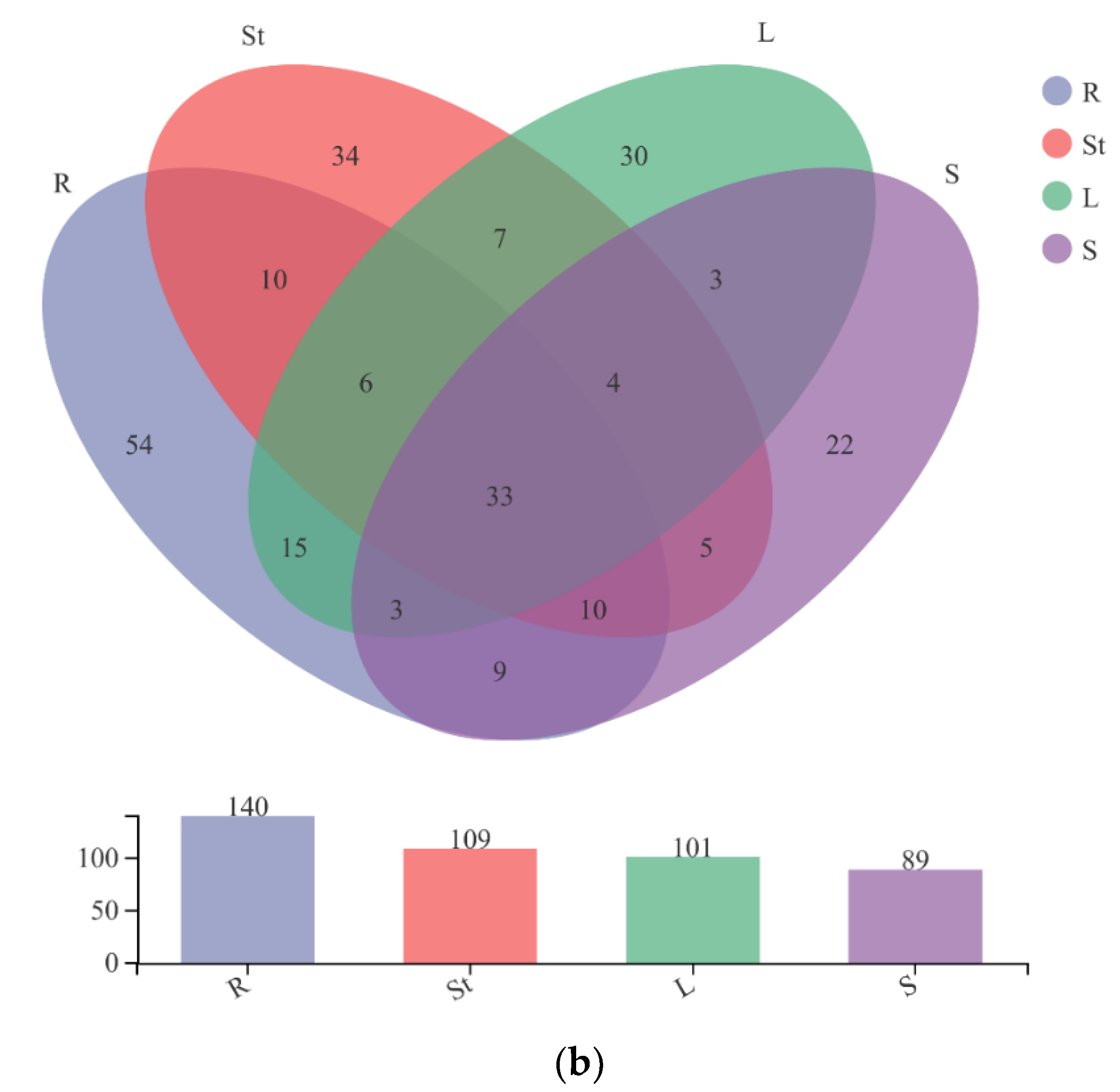
3.4. Fungal Composition and Relative Abundance among Different Organs of S. alopecuroides
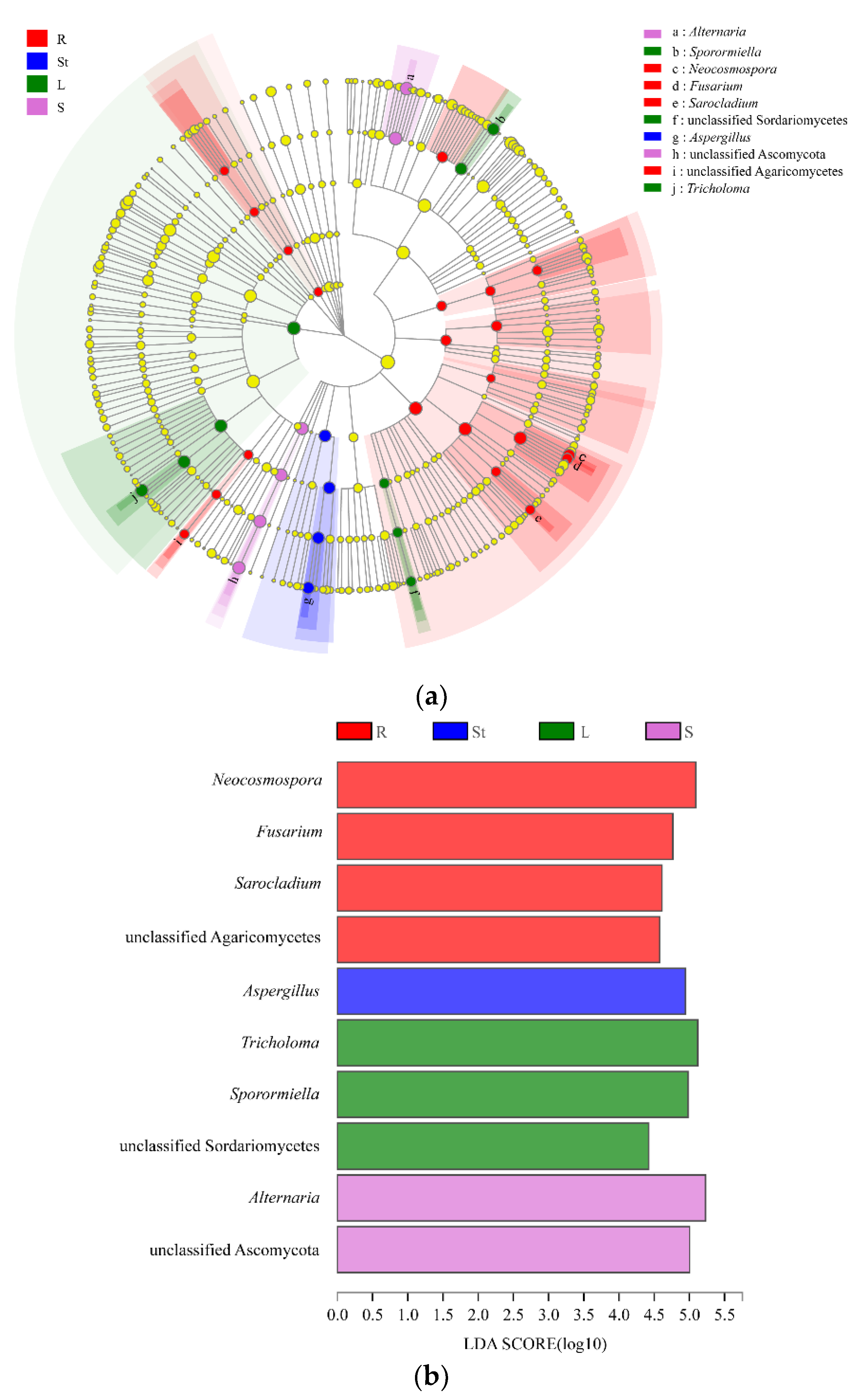
3.5. Analysis of Endophytic Fungal Biomarkers in Different Organs of S. alopecuroides
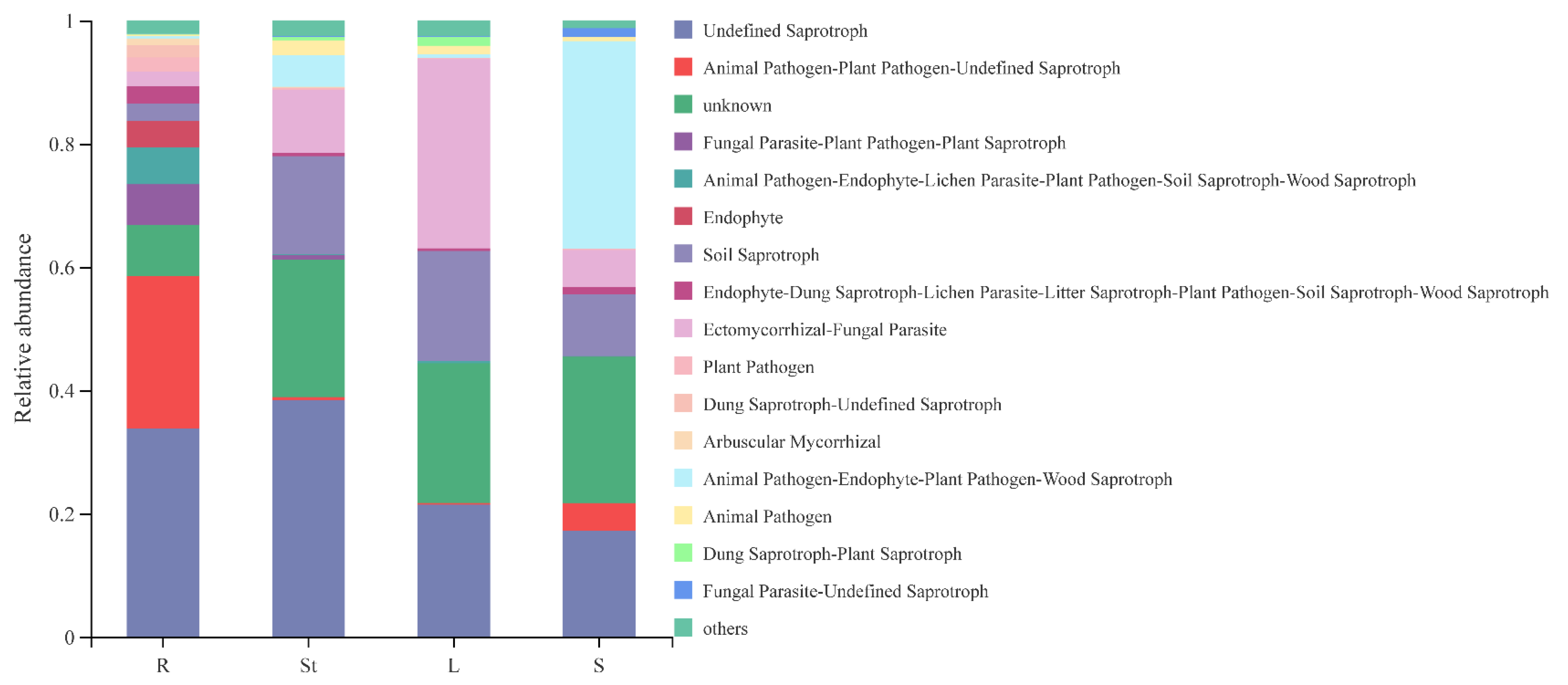
3.6. Functional Guild Function Prediction of Endophytic Fungi in Different Organs of S. alopecuroides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, S.; Chaturvedi, P.; Kulkarni, M.G.; Van Staden, J. A critical review on exploiting the pharmaceutical potential of plant endophytic fungi. Biotechnol. Adv. 2020, 39, 107462. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Ma, S.; Shi, X.; Xue, W.; Liu, C.; Ding, H. Diversity and Antimicrobial Activity of Endophytic Fungi Isolated from Chloranthus japonicus Sieb in Qinling Mountains, China. Int. J. Mol. Sci. 2020, 21, 5958. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Simon, J.C.; Marchesi, J.R.; Mougel, C.; Selosse, M.A. Host-microbiota interactions: From holobiont theory to analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Wang, Y.; Sun, X.; Tang, K. Bioactive natural products from endophytes: A review. Appl. Biochem. Microbiol. 2011, 44, 136–142. [Google Scholar] [CrossRef]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A Treasure House of Bioactive Compounds of Medicinal Importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef] [Green Version]
- Vandana, U.K.; Rajkumari, J.; Singha, L.P.; Satish, L.; Alavilli, H.; Sudheer, P.; Chauhan, S.; Ratnala, R.; Satturu, V.; Mazumder, P.B.; et al. The Endophytic Microbiome as a Hotspot of Synergistic Interactions, with Prospects of Plant Growth Promotion. Biology 2021, 10, 101. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes: Unique plant inhabitants with great promises. Appl. Microbiol. Biotechnol. 2011, 90, 1829–1845. [Google Scholar] [CrossRef]
- Hartley, S.E.; Eschen, R.; Horwood, J.M.; Gange, A.C.; Hill, E.M. Infection by a foliar endophyte elicits novel arabidopside-based plant defence reactions in its host, Cirsium arvense. New Phytol. 2015, 205, 816–827. [Google Scholar] [CrossRef] [Green Version]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Hückelhoven, R.; Neumann, C.; Wettstein, D.v.; et al. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhou, T.; Li, D.; Zhang, X.; Yu, W.; Cai, J.; Wang, G.; Guo, Q.; Yang, X.; Cao, F. The genetic diversity and population structure of Sophora alopecuroides (Faboideae) as determined by microsatellite markers developed from transcriptome. PLoS ONE 2019, 14, e0226100. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Deng, X.; Gao, Q.; Wu, X.; Han, L.; Gao, X.; Zhao, S.; Chen, W.; Zhou, R.; Li, Z.; et al. Sophora alopecuroides L.: An ethnopharmacological, phytochemical, and pharmacological review. J. Ethnopharmacol. 2020, 248, 112172. [Google Scholar] [CrossRef]
- Gu, P.; Hao, L.; Xu, R.; Hu, M.; Ma, H. Diversity and antimicrobial activity of endophytic fungi in Sophora alopecuroides L. from Baijitan National Nature Reserve of Ningxia. J. Northwest AF Univ. Nat. Sci. Ed. 2012, 40, 209–216. [Google Scholar]
- Gao, Y.; Zhou, X.; Sun, M.; Lv, M.; Gu, P. Diversity and Ecological Distribution of Endophytic Fungi of Sophora alopecuroides L. from Baijitan Nature Reserve of Ningxia. Acta Agrestia Sin. 2016, 24, 1309–1317. [Google Scholar] [CrossRef]
- Yan, K.; Pei, Z.; Meng, L.; Zheng, Y.; Wang, L.; Feng, R.; Li, Q.; Liu, Y.; Zhao, X.; Wei, Q.; et al. Determination of Community Structure and Diversity of Seed-Vectored Endophytic Fungi in Alpinia zerumbet. Front. Microbiol. 2022, 13, 814864. [Google Scholar] [CrossRef]
- Han, S.; Wang, Y.; Li, Y.; Shi, K. Investigation of bacterial diversity in Cajanus cajan-planted gangue soil via high-throughput sequencing. Bioengineered 2021, 12, 6981–6995. [Google Scholar] [CrossRef]
- Glynou, K.; Nam, B.; Thines, M.; Macia-Vicente, J.G. Facultative root-colonizing fungi dominate endophytic assemblages in roots of nonmycorrhizal Microthlaspi species. New Phytol. 2018, 217, 1190–1202. [Google Scholar] [CrossRef] [Green Version]
- Ren, F.; Dong, W.; Yan, D.H. Organs, Cultivars, Soil, and Fruit Properties Affect Structure of Endophytic Mycobiota of Pinggu Peach Trees. Microorganisms 2019, 7, 322. [Google Scholar] [CrossRef] [Green Version]
- Xue, F.; Liu, T. DNA sequence and community structure diversity of multi-year soil fungi in Grape of Xinjiang. Sci. Rep. 2021, 11, 16367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Guan, P.; Hao, C.; Yang, J.; Xie, Z.; Wu, D. Changes in assembly processes of soil microbial communities in forest-to-cropland conversion in Changbai Mountains, northeastern China. Sci. Total Environ. 2022, 818, 151738. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, B.; Zhang, C.; Fan, Z.; Chen, Y.; Xin, B.; Xie, Q. Current Progression: Application of High-Throughput Sequencing Technique in Space Microbiology. Biomed. Res. Int. 2020, 2020, 4094191. [Google Scholar] [CrossRef] [PubMed]
- Rego, S.M.; Snyder, M.P. High Throughput Sequencing and Assessing Disease Risk. Cold Spring Harb. Perspect. Med. 2019, 9, a026849. [Google Scholar] [CrossRef]
- Chan, A.W.; Naphtali, J.; Schellhorn, H.E. High-throughput DNA sequencing technologies for water and wastewater analysis. Sci. Prog. 2019, 102, 351–376. [Google Scholar] [CrossRef]
- Latz, M.A.C.; Kerrn, M.H.; Sorensen, H.; Collinge, D.B.; Jensen, B.; Brown, J.K.M.; Madsen, A.M.; Jorgensen, H.J.L. Succession of the fungal endophytic microbiome of wheat is dependent on tissue-specific interactions between host genotype and environment. Sci. Total Environ. 2021, 759, 143804. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Cacciola, S.O.; Mosca, S.; Zappia, R.; Schena, L. Analysis of the Fungal Diversity in Citrus Leaves with Greasy Spot Disease Symptoms. Microb. Ecol. 2017, 73, 739–749. [Google Scholar] [CrossRef]
- Alibrandi, P.; Schnell, S.; Perotto, S.; Cardinale, M. Diversity and Structure of the Endophytic Bacterial Communities Associated with Three Terrestrial Orchid Species as Revealed by 16S rRNA Gene Metabarcoding. Front. Microbiol. 2020, 11, 604964. [Google Scholar] [CrossRef]
- Hou, Q.Z.; Chen, D.W.; Wang, Y.P.; Ehmet, N.; Ma, J.; Sun, K. Analysis of endophyte diversity of two Gentiana plants species and the association with secondary metabolite. BMC Microbiol. 2022, 22, 90. [Google Scholar] [CrossRef]
- Cui, L.; Noushahi, H.A.; Zhang, Y.; Liu, J.; Cosoveanu, A.; Liu, Y.; Yan, L.; Zhang, J.; Shu, S. Endophytic Fungal Community of Huperzia serrata: Diversity and Relevance to the Production of Huperzine A by the Plant Host. Molecules 2021, 26, 892. [Google Scholar] [CrossRef]
- Liang, Y.; Wei, G.; Ning, K.; Zhang, G.; Liu, Y.; Dong, L.; Chen, S. Contents of lobetyolin, syringin, and atractylolide III in Codonopsis pilosula are related to dynamic changes of endophytes under drought stress. Chin. Med. 2021, 16, 122. [Google Scholar] [CrossRef]
- Ju, M.; Zhang, Q.; Wang, R.; Yan, S.; Li, Z.; Li, P.; Gu, P. Correlation in endophytic fungi community diversity and bioactive compounds of Sophora alopecuroides. Front. Microbiol. 2022, 13, 955647. [Google Scholar] [CrossRef]
- Houlden, A.; Timms-Wilson, T.M.; Day, M.J.; Bailey, M.J. Influence of plant developmental stage on microbial community structure and activity in the rhizosphere of three field crops. FEMS Microbiol. Ecol. 2008, 65, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; He, X.; Yuan, H.; Lv, G. Differed Growth Stage Dynamics of Root-Associated Bacterial and Fungal Community Structure Associated with Halophytic Plant Lycium ruthenicum. Microorganisms 2022, 10, 1644. [Google Scholar] [CrossRef]
- Ajilogba, C.F.; Olanrewaju, O.S.; Babalola, O.O. Plant Growth Stage Drives the Temporal and Spatial Dynamics of the Bacterial Microbiome in the Rhizosphere of Vigna subterranea. Front. Microbiol. 2022, 13, 825377. [Google Scholar] [CrossRef]
- Puri, S.C.; Nazir, A.; Chawla, R.; Arora, R.; Riyaz-ul-Hasan, S.; Amna, T.; Ahmed, B.; Verma, V.; Singh, S.; Sagar, R.; et al. The endophytic fungus Trametes hirsuta as a novel alternative source of podophyllotoxin and related aryl tetralin lignans. J. Biotechnol. 2006, 122, 494–510. [Google Scholar] [CrossRef]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Duan, Y.; Lian, J.; Wang, L.; Wang, X.; Luo, Y.; Wang, W.; Wu, F.; Zhao, J.; Ding, Y.; Ma, J.; et al. Variation in Soil Microbial Communities Along an Elevational Gradient in Alpine Meadows of the Qilian Mountains, China. Front. Microbiol. 2021, 12, 684386. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Alawneh, L.; Debbabi, M.; Hassaine, F.; Jarraya, Y.; Shahi, P.; Soeanu, A. Towards a unified paradigm for verification and validation of systems engineering design models. In Proceedings of the IASTED, Innsbruck(AT), Innsbruck, Austria, 15–17 February 2006. [Google Scholar]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Siddique, A.B.; Biella, P.; Unterseher, M.; Albrectsen, B.R. Mycobiomes of Young Beech Trees Are Distinguished by Organ Rather than by Habitat, and Community Analyses Suggest Competitive Interactions among Twig Fungi. Front. Microbiol. 2021, 12, 646302. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Wu, Q.; Wei, D.; Dong, L.; Liu, Y.; Ren, C.; Liu, Q.; Chen, C.; Chen, J.; Pei, J. Variation in the microbial community contributes to the improvement of the main active compounds of Magnolia officinalis Rehd. et Wils in the process of sweating. Chin. Med. 2019, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Ecology, P.G.K.A.J.F. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; He, S.; Zhao, Q. Isolation and Identification of Matrine-Producing Fungal Endophytes from Sophora alopecuroides in Ningxia. Sci. Agric. Sin. 2013, 46, 2643–2654. [Google Scholar] [CrossRef]
- Gamboa, M.A.; Laureano, S.; Bayman, P. Measuring diversity of endophytic fungi in leaf fragments: Does size matter? Mycopathologia 2002, 156, 41–45. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Bahram, M.; Sanchez-Castro, I.; Dai, D.Q.; Ariyawansa, K.; Jayalal, U.; Suwannarach, N.; Tedersoo, L. Current Insight into Culture-Dependent and Culture-Independent Methods in Discovering Ascomycetous Taxa. J. Fungi 2021, 7, 703. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [Green Version]
- Berg, G.; Rybakova, D.; Grube, M.; Köberl, M. The plant microbiome explored: Implications for experimental botany. J. Exp. Bot. 2016, 67, 995–1002. [Google Scholar] [CrossRef] [Green Version]
- Küngas, K.; Bahram, M.; Põldmaa, K. Host tree organ is the primary driver of endophytic fungal community structure in a hemiboreal forest. FEMS Microbiol. Ecol. 2020, 96, fiz199. [Google Scholar] [CrossRef]
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The Plant Microbiota: Systems-Level Insights and Perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Guo, L.; Zang, W.; Ping, W.; Chi, D. Diversity and ecological distribution of endophytic fungi associated with medicinal plants. Sci. China C Life Sci. 2008, 51, 751–759. [Google Scholar] [CrossRef]
- Su, Y.; Guo, L.; Hyde, K.D. Response of endophytic fungi of Stipa grandis to experimental plant function group removal in Inner Mongolia steppe, China. Fungal Divers. 2010, 43, 93–101. [Google Scholar] [CrossRef]
- Chen, M.; Chen, J.; Liu, J.; Wu, M.; Yan, Q.; Li, P.; Huang, L.; Xiao, X. Diversity analysis of rhizosphere soil fungi and endophytic fungi in Ampelocalamus luodianensis. Acta Ecol. Sin. 2021, 41, 4120–4130. [Google Scholar] [CrossRef]
- Yao, X.; Kang, Q.; Xiong, S.; Li, F.; Wang, Y.; Lin, S.; Bai, L.; Ma, W.; Deng, Z. Isolation and identification of endophytic actinomycetes from the seeds of Camptotheca acuminata Decne. and isolation of antimicrobial substances from those endophytic actinomycetes. Microbiol. China 2014, 41, 1109–1120. [Google Scholar] [CrossRef]
- Root, R. The niche exploitation pattern of the blue-grey gnatcatcher. Ecol. Monogr. 1967, 37, 317–350. [Google Scholar] [CrossRef]
- Zhou, J.; Xie, T.; Liu, J.; Lan, L.; Xu, Y.; Liu, Z.; Ai, Y.; Zhang, Q. Community structure and biological function of the root symbiotic fungi of wild Cymbidium ensifolium. Acta Microbiol. Sin. 2021, 61, 2136–2153. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, M.; Yu, L.; Liu, J.; Zou, X. The structure and function of endophytic fungal community in tobacco root. Mycosystema 2019, 38, 1610–1619. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Baarlen, P.V.; Belkum, A.V.; Summerbell, R.C.; Crous, P.W.; Thomma, B.P.H.J. Molecular mechanisms of pathogenicity: How do pathogenic microorganisms develop cross-kingdom host jumps? FEMS Microbiol. Rev. 2007, 31, 239–277. [Google Scholar] [CrossRef] [Green Version]
- Nagy, N.E.; Fossdal, C.G. Host responses in Norway spruce roots induced to the pathogen Ceratocystis polonica are evaded or suppressed by the ectomycorrhizal fungus Laccaria bicolor. Plant Biol. 2013, 15, 99–110. [Google Scholar] [CrossRef]
- Grau, O.; Geml, J.; Pérez-Haase, A.; Ninot, J.M.; Semenova-Nelsen, T.A.; Peñuelas, J. Abrupt changes in the composition and function of fungal communities along an environmental gradient in the high Arctic. Mol. Ecol. 2017, 26, 4798–4810. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Wang, C.; Wang, H.; Chen, H.; Tang, M. Pine wilt disease alters soil properties and root-associated fungal communities in Pinus tabulaeformis forest. Plant Soil 2016, 404, 237–249. [Google Scholar] [CrossRef]
- Millberg, H.; Boberg, J.; Stenlid, J. Changes in fungal community of Scots pine (Pinus sylvestris) needles along a latitudinal gradient in Sweden. Fungal Ecol. 2015, 17, 126–139. [Google Scholar] [CrossRef]
- Semchenko, M.; Leff, J.W.; Lozano, Y.M.; Saar, S.; Davison, J.; Wilkinson, A.; Jackson, B.G.; Pritchard, W.J.; Long, J.R.D.; Oakley, S.; et al. Fungal diversity regulates plant-soil feedbacks in temperate grassland. Sci. Adv. 2018, 4, eaau4578. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Guo, S.; Huang, M.; Thorsten, L.H.; Wei, J. Ascomycota has a faster evolutionary rate and higher species diversity than Basidiomycota. Sci. China Life Sci. 2010, 53, 1163–1169. [Google Scholar] [CrossRef]
- Elkady, W.M.; Raafat, M.M.; Abdel-Aziz, M.M.; Al-Huqail, A.A.; Ashour, M.L.; Fathallah, N. Endophytic Fungus from Opuntia ficus-indica: A Source of Potential Bioactive Antimicrobial Compounds against Multidrug-Resistant Bacteria. Plants 2022, 11, 1070. [Google Scholar] [CrossRef]
- Morales-Sanchez, V.; Diaz, C.E.; Trujillo, E.; Olmeda, S.A.; Valcarcel, F.; Munoz, R.; Andres, M.F.; Gonzalez-Coloma, A. Bioactive Metabolites from the Endophytic Fungus Aspergillus sp. SPH2. J. Fungi 2021, 7, 109. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Q.; Gao, Y.Q.; Shi, X.W.; Gao, J.M. Antifungal and antibacterial metabolites from an endophytic Aspergillus sp. associated with Melia azedarach. Nat. Prod. Res. 2014, 28, 1388–1392. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kim, Y.-H.; Kang, S.-M.; Lee, J.-H.; Lee, I.-J. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Process Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Mehmood, A.; Hussain, A.; Irshad, M.; Hamayun, M.; Iqbal, A.; Khan, N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis 2018, 77, 225–235. [Google Scholar] [CrossRef]
- Kwon, Y.-J.; Sohn, M.-J.; Zheng, C.-J.; Kim, W.-G. Fumimycin: A Peptide Deformylase Inhibitor with an Unusual Skeleton Produced by Aspergillus fumisynnematus. Org. Lett. 2007, 9, 2449–2451. [Google Scholar] [CrossRef]
- Woudenberg, J.H.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, H.X.; Chen, Y.C.; Sun, Z.H.; Li, H.H.; Li, S.N.; Yan, M.L.; Zhang, W.M. Two New Metabolites from the Endophytic Fungus Alternaria sp. A744 Derived from Morinda officinalis. Molecules 2017, 22, 765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, L.; Gong, S.; Yang, L.; Hao, J.; Xue, M.; Zeng, F.; Zhang, X.; Shi, W.; Wang, H.; Yu, D. Biocontrol potential of endophytic fungi in medicinal plants from Wuhan Botanical Garden in China. Biol. Control 2016, 94, 47–55. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, Q.; Hu, L.; Li, W.; Yan, S.; Lv, M.; Gu, P. Real-time fluorescent quantitative PCR detection of key enzyme genes expression of alkaloid biosynthesis promoted by endophytic fungal elicitor in Sophora alopecuroides. Chin. Tradit. Herb. Drugs 2018, 49, 4621–4627. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sequence | Ace | Simpson | Shannoneven | PD | |
|---|---|---|---|---|---|---|
| site | H | 58,139 ± 25,596 | 90.55 ± 32.86 a | 0.31 ± 0.24 | 0.44 ± 0.16 | 26.03 ± 5.42 |
| B | 53,271 ± 9435 | 64.25 ± 23.22 b | 0.27 ± 0.11 | 0.49 ± 0.09 | 29.12 ± 6.96 | |
| organ | R | 53,012 ± 3960 | 106.24 ± 34.43 a | 0.23 ± 0.14 | 0.49 ± 0.11 | 28.59 ± 5.02 |
| St | 53,832 ± 7679 | 71.31 ± 28.21 a | 0.26 ± 0.23 | 0.51 ± 0.16 | 29.27 ± 5.98 | |
| L | 67,764 ± 33,229 | 73.20 ± 28.86 a | 0.38 ± 0.26 | 0.43 ± 0.17 | 25.94 ± 7.16 | |
| S | 48,211 ± 15,229 | 58.85 ± 10.10 b | 0.29 ± 0.07 | 0.43 ± 0.06 | 26.48 ± 7.83 | |
| Comparison | R2 | p-Value | |
|---|---|---|---|
| Sites comparison | H vs. B | 0.0728 | 0.068 |
| Organ comparison | Roots vs. Stems | 0.3064 | 0.002 |
| Roots vs. Leaves | 0.2935 | 0.002 | |
| Roots vs. Seeds | 0.3585 | 0.002 | |
| Stems vs. Leaves | 0.0625 | 0.723 | |
| Stems vs. Seeds | 0.1713 | 0.048 | |
| Leaves vs. Seeds | 0.2198 | 0.015 | |
| Characteristics | SumsOfSqs | MeanSqs | F. Model | R2 | p. Value | P. Adjust |
|---|---|---|---|---|---|---|
| organ | 2.52902 | 0.84301 | 3.28379 | 0.33001 | 0.001 | 0.002 |
| site | 0.55774 | 0.55774 | 1.72685 | 0.07278 | 0.06 | 0.06 |
| Group Label | Number of Genera | Genus and Their Relative Abundance (>0.02) |
|---|---|---|
| Shared genera in 4 organs | 33 | unc. Ascomycota (18.42%), Tricholoma (15.31%), Apiotrichum (14.47%), Alternaria (12.35%), Aspergillus (9.65%), unc. Didymellaceae (7.96%), Neocosmospora (7.39%), unc. Fungi (2.09%), Filobasidium (2.06%), Fusarium (2.01%), others (8.29%) |
| Root unique genera | 54 | Cadophora (50.41%), Setophoma (14.10%), Phaeomycocentrospora (9.30%), Leohumicola (2.68%), Ophiosimulans (2.47%), unc. Sordariales (2.45%), unc. Ophiocordycipitaceae (2.44%), Setophaeosphaeria (2.41%), others (13.75%) |
| Stem unique genera | 34 | Plenodomus (38.25%), Mrakia (23.13%), Sphaerosporella (6.15%), Phaeotremella (4.74%), unc. Microascaceae (3.77%), unc. Lasiosphaeriaceae (2.65%), Entrophospora (2.39%), Dominikia (2.37%), Clavulina (2.13%), Biatriospora (2.01%), others (12.40%) |
| Leaf unique genera | 30 | Dioszegia (23.43%), Sistotrema (23.21%), Thanatephorus (11.10%), Hawksworthiomyces (8.41%), Saitozyma (5.61%), Gonytrichum (3.92%), unc. Xylariales (3.81%), Humaria (3.48%), Hydnellum (2.58%), unc. Metschnikowiaceae (2.24), unc. Mycosphaerellaceae (2.13%), others (10.09%) |
| Seed unique genera | 22 | Papiliotrema (69.88%), Curvularia (4.46%), Stereum (3.68%), unc. Pezizales (2.48%), others (19.50%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Zhang, Q.; Ju, M.; Yan, S.; Zhang, Q.; Gu, P. The Endophytic Fungi Diversity, Community Structure, and Ecological Function Prediction of Sophora alopecuroides in Ningxia, China. Microorganisms 2022, 10, 2099. https://doi.org/10.3390/microorganisms10112099
Wang R, Zhang Q, Ju M, Yan S, Zhang Q, Gu P. The Endophytic Fungi Diversity, Community Structure, and Ecological Function Prediction of Sophora alopecuroides in Ningxia, China. Microorganisms. 2022; 10(11):2099. https://doi.org/10.3390/microorganisms10112099
Chicago/Turabian StyleWang, Ruotong, Qingchen Zhang, Mingxiu Ju, Siyuan Yan, Qiangqiang Zhang, and Peiwen Gu. 2022. "The Endophytic Fungi Diversity, Community Structure, and Ecological Function Prediction of Sophora alopecuroides in Ningxia, China" Microorganisms 10, no. 11: 2099. https://doi.org/10.3390/microorganisms10112099