
Colonization Ability of Bacillus subtilis NCD-2 in Different Crops and Its Effect on Rhizosphere Microorganisms
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Culture Conditions
2.2. Plant Growth Experiment
2.3. Assays of Growth Indices
2.4. Soil Sample Collection and DNA Extraction
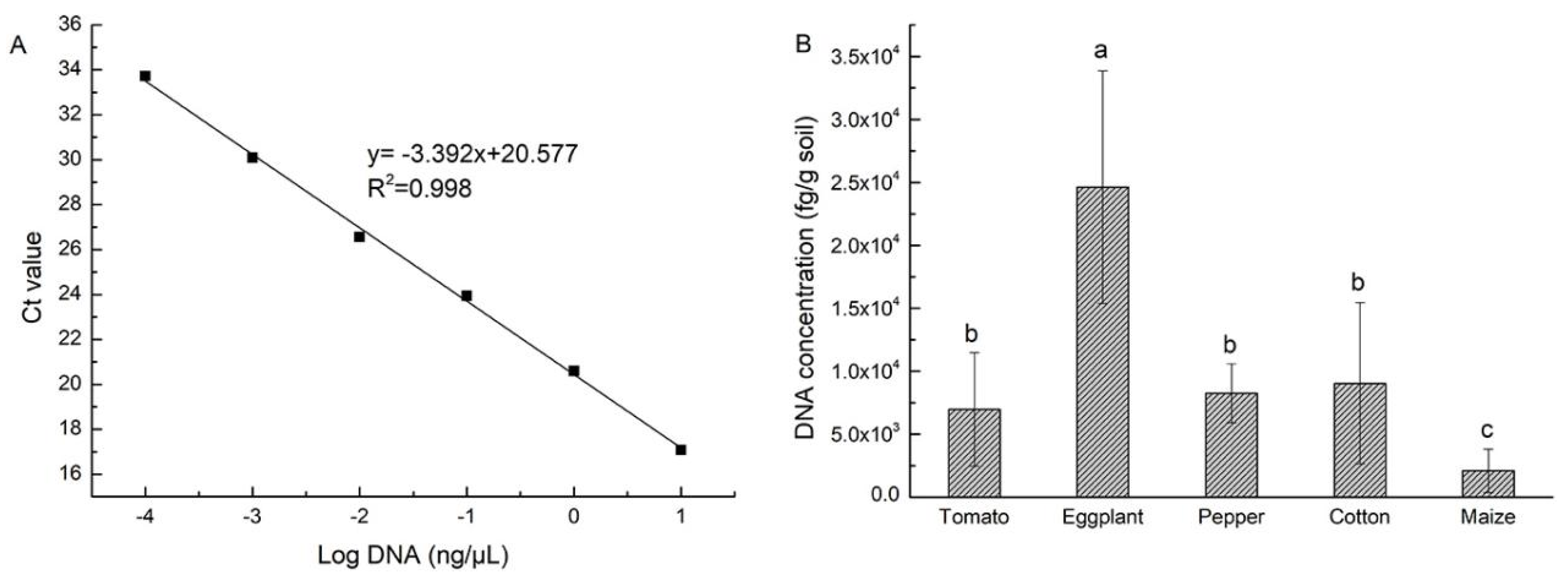
2.5. Quantitative Detection of Strain NCD-2 in the Rhizosphere of Five Crops
2.6. PCR Amplification, Illumina Sequencing and Data Processing
2.7. Statistical Analysis
3. Results
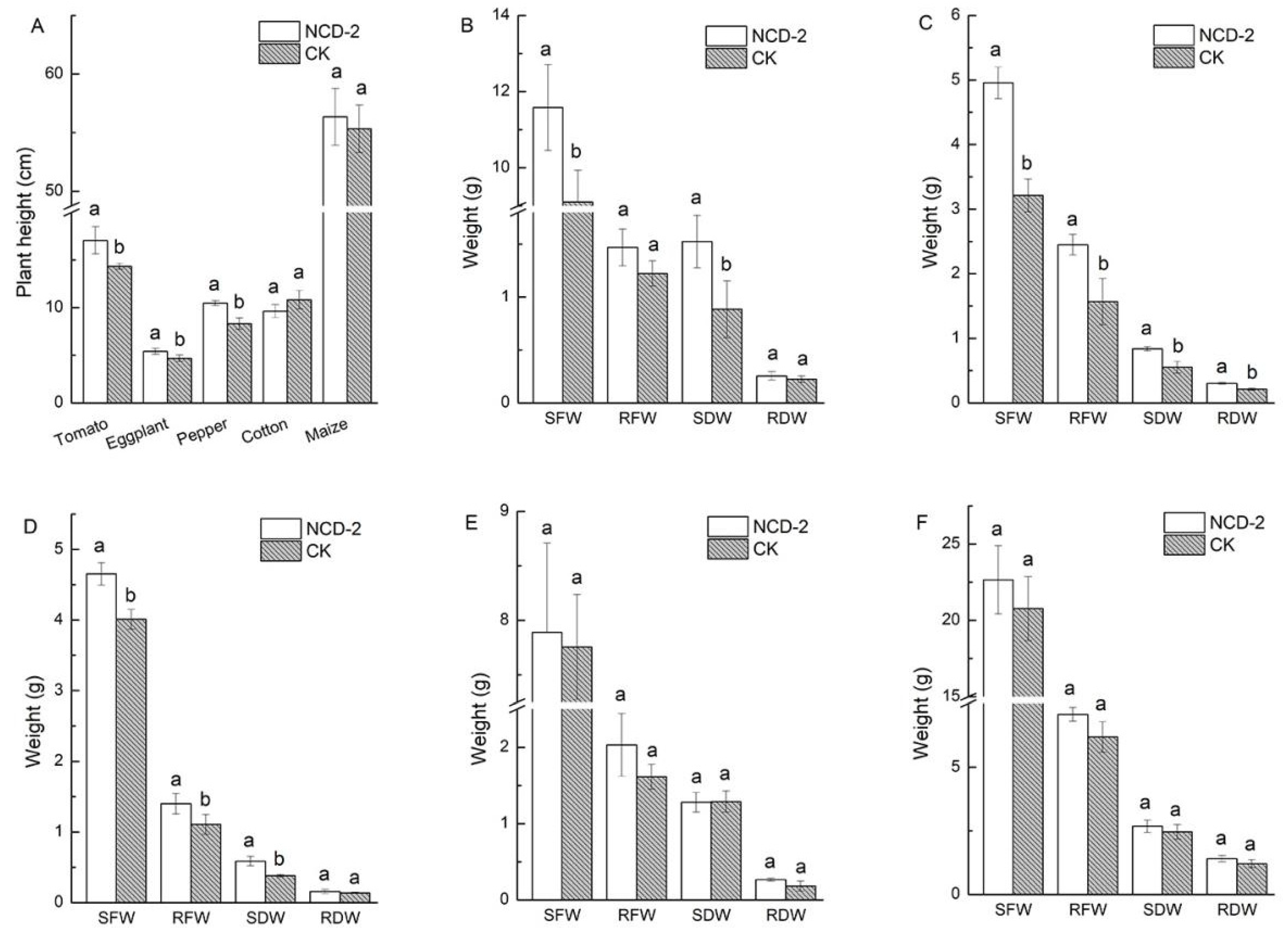
3.1. Effect of Strain NCD-2 on the Biomass for Different Crops
3.2. Populations of Strain NCD-2 in Rhizosphere
3.3. Sequencing Data Summary and Microbial Community Diversity
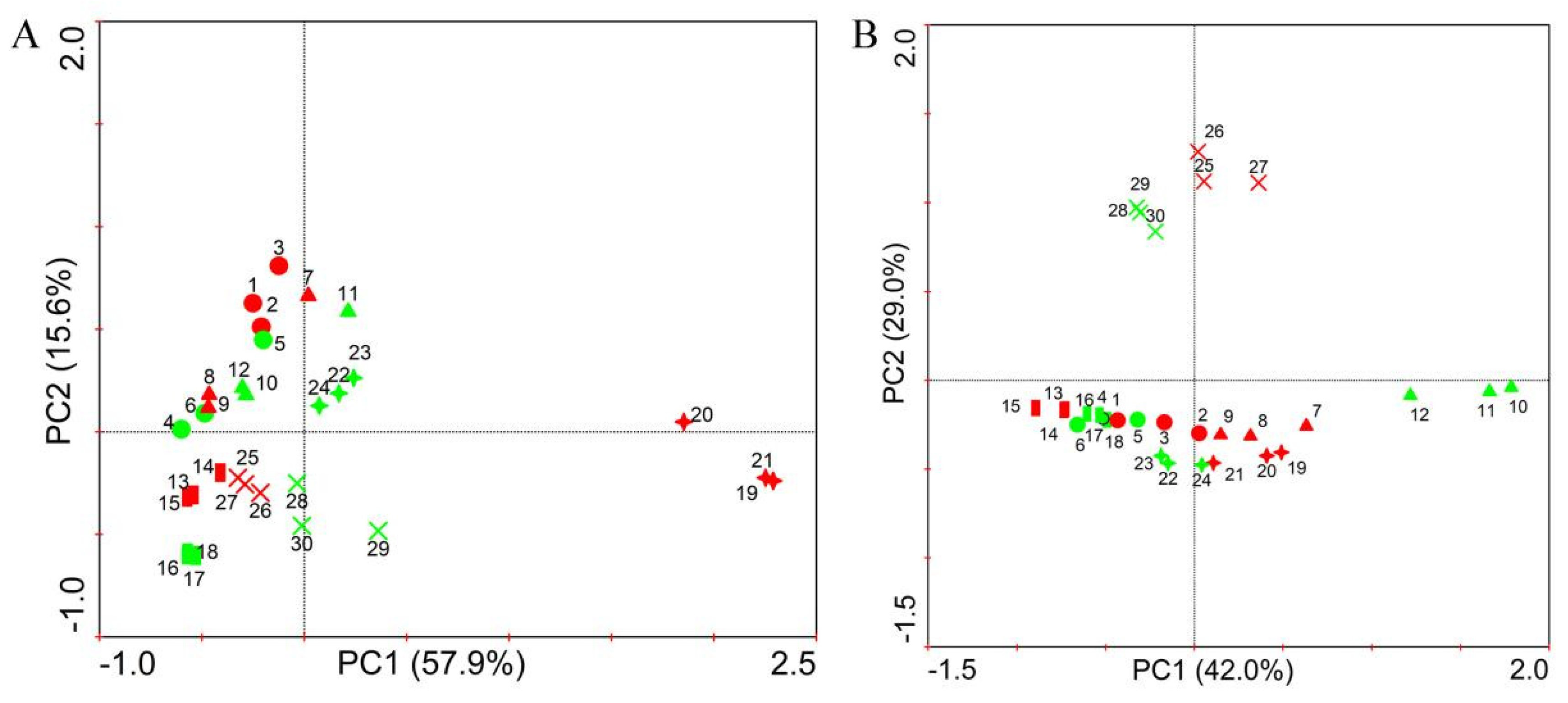
3.4. Effect of Strain NCD-2 on the Microbial Community Structure in the Rhizosphere of Different Crops
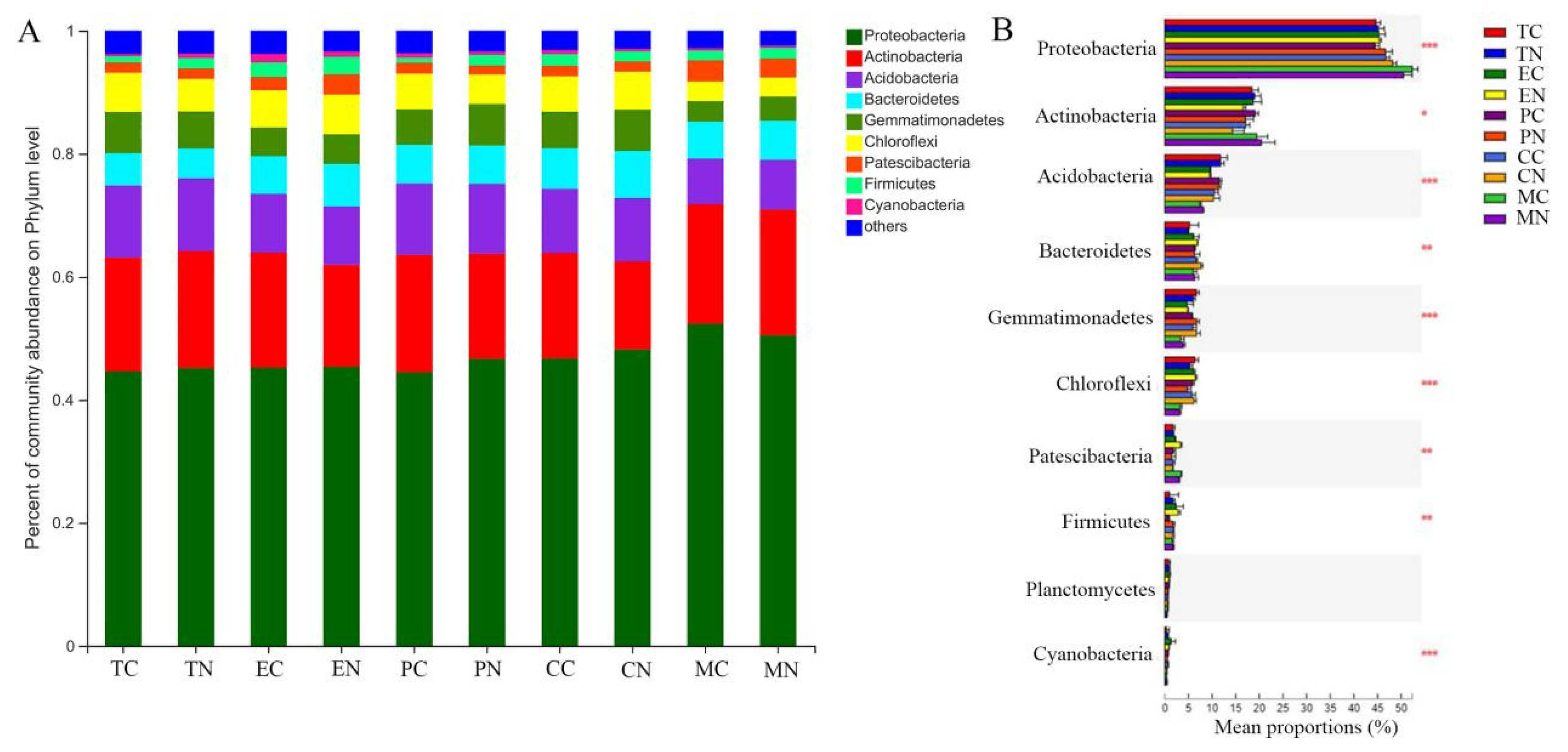
3.5. Effect of Strain NCD-2 on the Bacterial Taxonomic Community Composition in the Rhizosphere of Different Crops
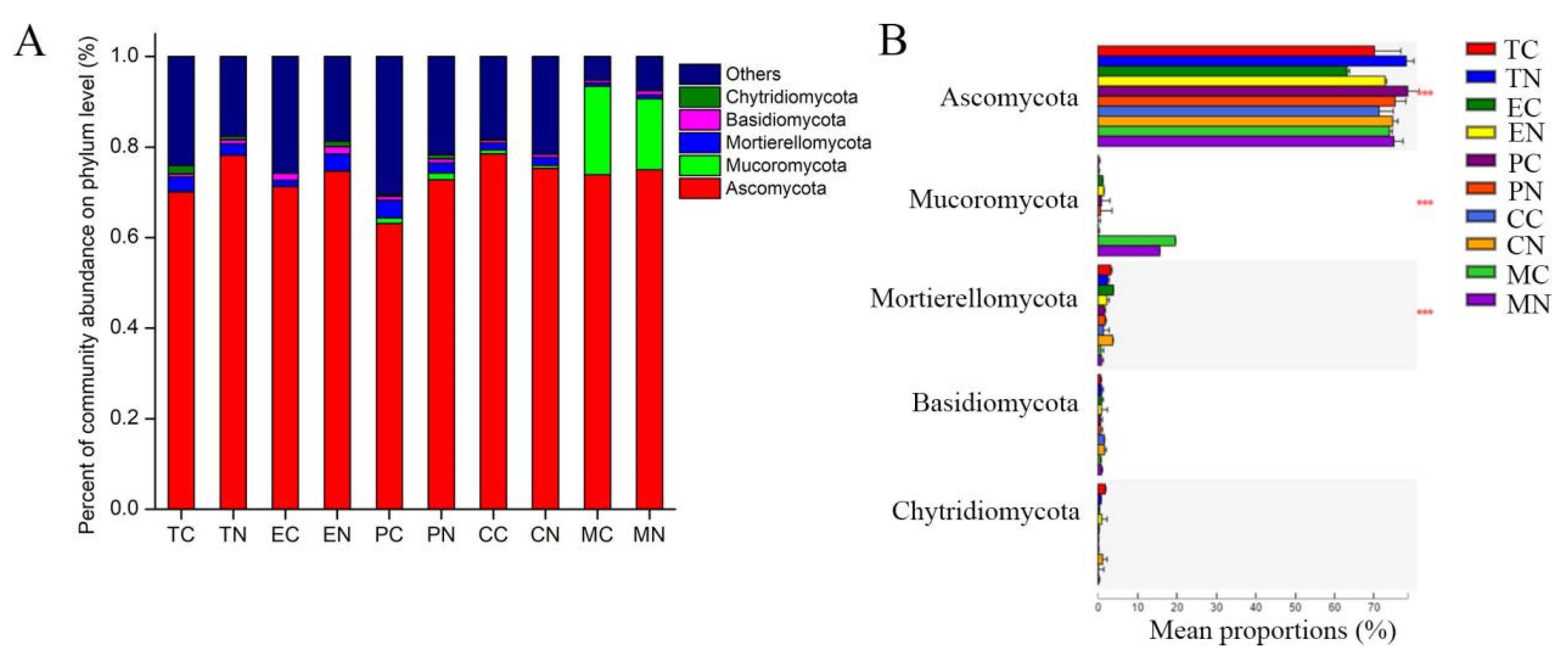
3.6. Effect of Strain NCD-2 on the Fungal Taxonomic Community Composition in the Rhizosphere of Different Crops
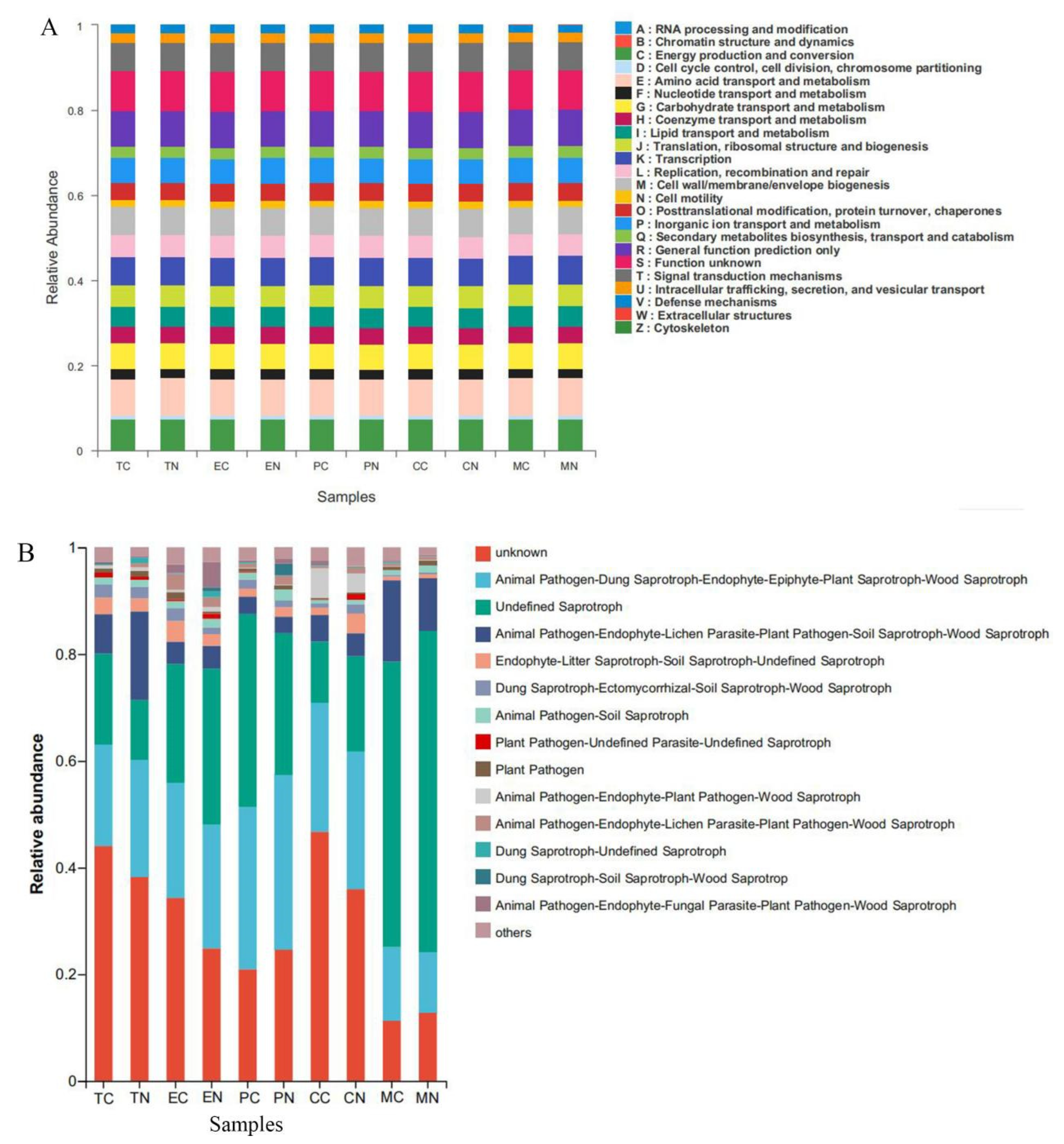
3.7. Predicted Functional Analysis of the Microbial Community
4. Discussion
4.1. The Colonization and Growth-Promoting Ability of Strain NCD-2
4.2. The Influence of Strain NCD-2 on Microbial Communities Varied among Plant Species
4.3. Variations in Microbial Functional Metabolic Genes for Five Crops by Strain NCD-2
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, S.; Pandey, P.; Maheshwari, D.K. Reduction in dose of chemical fertilizers and growth enhancement of sesame (Sesamum indicum L.) with application of rhizospheric competent Pseudomonas aeruginosa LES4. Eur. J. Soil Biol. 2009, 45, 334–340. [Google Scholar] [CrossRef]
- Ayala, S.; Rao, E. Perspectives of soil fertility management with a focus on fertilizer use for crop productivity. Curr. Sci. 2002, 82, 797–807. [Google Scholar]
- Li, F.; Zhang, S.Q.; Wang, Y.; Li, Y.; Li, P.P.; Chen, L.; Jie, X.L.; Hu, D.S.; Feng, B.; Yue, K.; et al. Rare fungus, Mortierella capitata, promotes crop growth by stimulating primary metabolisms related genes and reshaping rhizosphere bacterial community. Soil Biol. Biochem. 2020, 151, 108017. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.J.G.; Biology, M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Thakur, S.; Dhingra, G.K.; Singh, A.; Pal, M.K.; Harshvardhan, K.; Dubey, R.C.; Maheshwari, D.K. Inoculation of siderophore producing rhizobacteria and their consortium for growth enhancement of wheat plant. Biocatal. Agric. Biotechnol. 2018, 15, 264–269. [Google Scholar] [CrossRef]
- Dimkpa, C.; Weinand, T.; Asch, F. Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef]
- Bukhat, S.; Imran, A.; Javaid, S.; Shahid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defense signaling. Microbiol. Res. 2020, 238, 126486. [Google Scholar] [CrossRef]
- Qin, Y.; Druzhinina, I.S.; Pan, X.Y.; Yuan, Z.L. Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef]
- Uroz, S.; Oger, P.; Tisserand, E.; Cébron, A.; Turpault, M.P.; Buée, M.; De Boer, W.; Leveau, J.H.J.; Frey-Klett, P. Specific impacts of beech and Norway spruce on the structure and diversity of the rhizosphere and soil microbial communities. Sci. Rep. 2016, 6, 27756. [Google Scholar] [CrossRef] [Green Version]
- Kusstatscher, P.; Wicaksono, W.A.; Thenappan, D.P.; Adam, E.; Berg, G.J.M. Microbiome management by biological and chemical treatments in maize is linked to plant health. Microorganisms 2020, 8, 1506. [Google Scholar] [CrossRef]
- Mathur, P.; Roy, S. Insights into the plant responses to drought and decoding the potential of root associated microbiome for inducing drought tolerance. Physiol. Plantarum. 2021, 172, 1016–1029. [Google Scholar] [CrossRef] [PubMed]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial interactions in the rhizosphere: Beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Merbach, W.; Mirus, E.; Knof, G.; Remus, R.; Ruppel, S.; Russow, R.; Gransee, A.; Schulze, J. Release of carbon and nitrogen compounds by plant roots and their possible ecological importance. J. Plant Nutr. Soil Sci. 1999, 162, 373–383. [Google Scholar] [CrossRef]
- Pii, Y.; Borruso, L.; Brusetti, L.; Crecchio, C.; Cesco, S.; Mimmo, T. The interaction between iron nutrition, plant species and soil type shapes the rhizosphere microbiome. Plant Physiol. Biochem. 2016, 99, 39–48. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Kupper, K.C.; Moretto, R.K.; Fujimoto, A. Production of antifungal compounds by Bacillus spp. isolates and its capacity for controlling citrus black spot under field conditions. World J. Microb. Biot. 2020, 36, 7. [Google Scholar] [CrossRef]
- Mazzoli, A.; Donadio, G.; Lanzilli, M.; Saggese, A.; Isticato, R. Bacillus megaterium SF185 spores exert protective effects against oxidative stress in vivo and in vitro. Sci. Rep. 2019, 9, 12082. [Google Scholar] [CrossRef] [Green Version]
- Li, S.Z.; Ma, P.; Gao, S.G.; Liu, X.Z.; Liu, G. Evaluation of biocontrol potential of a bacterial strain NCD-2 against cotton verticillium wilt in field trials. Acta Phytopathol. Sin. 2005, 35, 451–455. [Google Scholar]
- Zhao, W.S.; Guo, Q.G.; Li, S.Z.; Wang, P.P.; Dong, L.H.; Su, Z.H.; Zhang, X.Y.; Lu, X.Y.; Ma, P. Effects of Bacillus subtilis NCD-2 and broccoli residues return on potato verticillium wilt and soil fungal community structure. Biol. Control 2021, 159, 104628. [Google Scholar] [CrossRef]
- Huang, Z.Q.; Liu, B.W.; Yin, Y.; Liang, F.; Xie, D.S.; Han, T.T.; Liu, Y.Z.; Yan, B.; Li, Q.; Huang, Y.; et al. Impact of biocontrol microbes on soil microbial diversity in ginger (Zingiber officinale Roscoe). Pest Manag. Sci. 2021, 77, 5537–5546. [Google Scholar] [CrossRef]
- Sylla, J.; Alsanius, B.W.; Krüger, E.; Reineke, A.; Strohmeier, S.; Wohanka, W. Leaf microbiota of strawberries as affected by biological control agents. Phytopathology 2013, 103, 1001–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.J.; Wang, Z.Y.; Li, N.; Wang, Y.H.; Feng, J.T.; Zhang, X. Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community. Appl. Soil Ecol. 2019, 136, 55–66. [Google Scholar] [CrossRef]
- Abdallah, D.B.; Frikha-Gargouri, O.; Tounsi, S. Rizhospheric competence, plant growth promotion and biocontrol efficacy of Bacillus amyloliquefaciens subsp. plantarum strain 32a. Biol. Control 2018, 124, 61–67. [Google Scholar]
- Wang, C.; Zheng, M.M.; Song, W.F.; Wen, S.L.; Wang, B.R.; Zhu, C.Q.; Shen, R.F. Impact of 25 years of inorganic fertilization on diazotrophic abundance and community structure in an acidic soil in southern China. Soil Biol. Biochem. 2017, 113, 240–249. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, J.W.; Dong, L.H.; Li, X.; Zhang, L.Q. Fengycin produced by Bacillus subtilis NCD-2 is involved in suppression of clubroot on Chinese cabbage. Biol. Control 2019, 136, 104001. [Google Scholar] [CrossRef]
- Lu, R.; Li, Y.P.; Li, W.X.; Xie, Z.S.; Fan, C.L.; Liu, P.X.; Deng, S.X. Bacterial community structure in atmospheric particulate matters of different sizes during the haze days in Xi’an, China. Sci. Total Environ. 2018, 637–638, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.G.; Li, S.Z.; Lu, X.Y.; Li, B.Q.; Ma, P. PhoR/PhoP two component regulatory system affects biocontrol capability of Bacillus subtilis NCD-2. Genet. Mol. Biol. 2010, 33, 333–340. [Google Scholar] [CrossRef]
- Guo, Q.G.; Dong, W.X.; Li, S.Z.; Lu, X.Y.; Wang, P.P.; Zhang, X.Y.; Wang, Y.; Ma, P. Fengycin produced by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling damping-off disease. Microbiol. Res. 2014, 169, 533–540. [Google Scholar] [CrossRef]
- Zhang, Q.X.; Stummer, B.E.; Guo, Q.G.; Zhang, W.; Zhang, X.J.; Zhang, L.Q.; Harvey, P.R. Quantification of Pseudomonas protegens FD6 and Bacillus subtilis NCD-2 in soil and the wheat rhizosphere and suppression of root pathogenic Rhizoctonia solani AG-8. Biol. Control 2020, 154, 104504. [Google Scholar] [CrossRef]
- Shao, M.Q.; Zhao, W.S.; Su, Z.H.; Dong, L.H.; Guo, Q.G.; Ma, P. Effect of Bacillus subtilis NCD-2 on the growth of tomato and the microbial community structure of rhizosphere soil under salt stress. Sci. Agric. Sin. 2021, 54, 4573–4584. [Google Scholar]
- Zhang, N.; Wu, K.; He, X.; Li, S.Q.; Zhang, Z.H.; Shen, B.; Yang, X.M.; Zhang, R.F.; Huang, Q.W.; Shen, Q.R. A new bioorganic fertilizer can effectively control banana wilt by strong colonization with Bacillus subtilis N11. Plant Soil 2011, 344, 87–97. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, X.; Shen, Z.Z.; Zhu, C.Z.; Jiao, Z.X.; Li, R.; Shen, Q.R. Pre-colonization of PGPR triggers rhizosphere microbiota succession associated with crop yield enhancement. Plant Soil 2019, 439, 553–567. [Google Scholar] [CrossRef]
- Wen, T.; Zhao, M.L.; Yuan, J.; Kowalchuk, G.; Shen, Q.R. Root exudates mediate plant defense against foliar pathogens by recruiting beneficial microbes. Soil Ecol. Lett. 2021, 3, 42–51. [Google Scholar] [CrossRef]
- Singh, B.K.; Trivedi, P.; Egidi, E.; Macdonald, C.A.; Baquerizo, M.D. Crop microbiome and sustainable agriculture. Nat. Rev. Microbiol. 2020, 18, 601–602. [Google Scholar] [CrossRef]
- Stephen, A.R.; Joseph, G.; Jurriaan, T. Crying out for help with root exudates: Adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar]
- Zhou, Z.K.; Yu, J.; Wang, Z.; Gou, Z.H.; Lyu, J.; Li, W.Y.; Yu, Y.J.; Shu, L.P.; Zhao, Y.J.; Ma, Y.M.; et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 2015, 33, 408–414. [Google Scholar] [CrossRef] [Green Version]
- Wieland, G.; Neumann, R.; Backhaus, H. Variation of microbial communities in soil, rhizosphere, and rhizoplane in response to crop species, soil type, and crop development. Appl. Environ. Microb. 2001, 67, 5849–5854. [Google Scholar] [CrossRef] [Green Version]
- Goodrich, J.K.; Davenport, E.R.; Waters, J.L.; Clark, A.G.; Ley, R.E. Cross-species comparisons of host genetic associations with the microbiome. Science 2016, 352, 532–535. [Google Scholar] [CrossRef] [Green Version]
- Shade, A.; Stopnisek, N. Abundance-occupancy distributions to prioritize plant core microbiome membership. Curr. Opin. Microbiol. 2019, 49, 50–58. [Google Scholar] [CrossRef]
- Cipriano, M.A.P.; Lupatini, M.; Lopes-Santos, L.; da Silva, M.J.; Roesch, L.F.W.; Destéfano, S.A.L.; Freitas, S.S.; Kuramae, E.E. Lettuce and rhizosphere microbiome responses to growth promoting Pseudomonas species under field conditions. FEMS Microbiol. Ecol. 2016, 92, fiw197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhauer, N.; Schulz, W.; Scheu, S.; Jousset, A. Niche dimensionality links biodiversity and invasibility of microbial communities. Funct. Ecol. 2013, 27, 282–288. [Google Scholar] [CrossRef]
- Griffiths, B.; Ritz, K.; Wheatley, R.; Kuan, H.; Boag, B.; Christensen, S.; Ekelund, F.; Sorensen, S.J.; Muller, S.; Bloem, J. An examination of the biodiversity-ecosystem function relationship in arable soil microbial communities. Soil Biol. Biochem. 2001, 33, 1713–1722. [Google Scholar] [CrossRef]
- Chen, L.; Wang, X.H.; Ma, Q.H.; Bian, L.S.; Liu, X.; Xu, Y.; Zhang, H.H.; Shao, J.H.; Liu, Y.P. Bacillus velezensis CLA178-induced systemic resistance of Rosa multiflora against crown gall disease. Front. Microbiol. 2020, 11, 587667. [Google Scholar] [CrossRef]
- Blake, C.; Nordgaard Christensen, M.; Kovács, Á.T. Molecular aspects of plant growth promotion and protection by Bacillus subtilis. Mol. Plant-Microbe Interact. 2021, 34, 15–25. [Google Scholar] [CrossRef]
- Shao, J.H.; Li, Y.C.; Li, Z.F.; Xu, Z.H.; Xun, W.B.; Zhang, N.; Feng, H.C.; Miao, Y.Z.; Shen, Q.R.; Zhang, R.F. Participating mechanism of a major contributing gene ysnE for auxin biosynthesis in Bacillus amyloliquefaciens SQR9. J. Basic Microb. 2021, 61, 569–575. [Google Scholar] [CrossRef]
- Ali, A.; Mohanta, T.K.; Asaf, S.; Rehman, N.; Al-Housni, S.; Al-Harrasi, A.; Khan, A.L.; Al-Rawahi, A. Biotransformation of benzoin by Sphingomonas sp. LK11 and ameliorative effects on growth of Cucumis sativus. Arch. Microbiol. 2019, 201, 591–601. [Google Scholar] [CrossRef]
- Halo, B.A.; Khan, A.L.; Waqas, M.; Al-Harrasi, A.; Hussain, J.; Ali, L.; Adnan, M.; Lee, I. Endophytic bacteria (Sphingomonas sp. LK11) and gibberellin can improve Solanum lycopersicum growth and oxidative stress under salinity. J. Plant Interact. 2015, 10, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Puopolo, G.; Raio, A.; Zoina, A. Identification and charactherization of Lysobacter capsici strain pg4: A new plant health-promoting rhizobacterium. J. Plant Pathol. 2010, 92, 157–164. [Google Scholar]
- Wang, Y.; Ohara, Y.; Nakayashiki, H.; Tosa, Y.; Mayama, S. Microarray analysis of the gene expression profile induced by the endophytic plant growth-promoting rhizobacteria, Pseudomonas fluorescens FPT9601-T5 in Arabidopsis. Mol. Plant-Microbe Interact. 2005, 5, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinayarani, G.; Madhusudhan, K.N.; Prakash, H.S. Induction of systemic resistance in turmeric by rhizospheric isolate Trichoderma asperellum against rhizome rot disease. J. Plant Pathol. 2019, 101, 965–980. [Google Scholar] [CrossRef]
- Masunaka, A.; Hyakumachi, M.; Takenaka, S. Plant growth-promoting fungus, Trichoderma koningi suppresses isoflavonoid phytoalexin vestitol production for colonization on/in the roots of Lotus japonicus. Microbes Environ. 2011, 26, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 2020, 11, 7. [Google Scholar] [CrossRef]
- Al-Askar, A.A.; Saber, W.; Ghoneem, K.M.; Hafez, E.E.; Ibrahim, A.A. Crude citric acid of Trichoderma asperellum: Tomato growth promotor and suppressor of Fusarium oxysporum f. sp. Lycopersici. Plants 2021, 10, 222. [Google Scholar] [CrossRef] [PubMed]
- Musyoki, M.K.; Cadisch, G.; Zimmermann, J.; Wainwright, H.; Beed, F.; Rasche, F. Soil properties, seasonality and crop growth stage exert a stronger effect on rhizosphere prokaryotes than the fungal biocontrol agent Fusarium oxysporum f. sp. Strigae. Appl. Soil Ecol. 2016, 105, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Fravel, D.R.; Deahl, K.L.; Stommel, J.R. Compatibility of the biocontrol fungus Fusarium oxysporum strain CS-20 with selected fungicides. Biol. Control 2005, 34, 165–169. [Google Scholar] [CrossRef]
- Su, Z.H.; Chen, X.Y.; Liu, X.M.; Guo, Q.G.; Li, S.Z.; Lu, X.Y.; Zhang, X.Y.; Wang, P.P.; Dong, L.H.; Zhao, W.S.; et al. Genome mining and UHPLC-MS/MS illuminate the specificity of secondary metabolite synthetic gene clusters in Bacillus subtilis NCD-2. BMC Genom. 2020, 21, 767. [Google Scholar] [CrossRef]
- Ogundeji, A.O.; Li, Y.; Liu, X.J.; Meng, L.B.; Sang, P.; Mu, Y.; Wu, H.L.; Ma, Z.N.; Hou, J.; Li, S.M. Eggplant by grafting enhanced with biochar recruits specific microbes for disease suppression of Verticillium wilt. Appl. Soil Ecol. 2021, 163, 103912. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; van Elsas, J.D.; Olff, H.; Salles, J.F. Dispersal-competition tradeoff in microbiomes in the quest for land colonization. Sci. Rep. 2018, 8, 9451. [Google Scholar] [CrossRef] [Green Version]
- Berg, N.V.D.; Berger, D.K.; Hein, I.; Birch, P.R.J.; Wingfield, M.J.; Viljoen, A. Tolerance in banana to Fusarium wilt is associated with early up-regulation of cell wall-strengthening genes in the roots. Mol. Plant Pathol. 2007, 8, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.P.; Xue, Z.Q.; Yang, Z.Y.; Chai, Z.; Niu, J.P.; Shi, Z.Y. Effects of microbial organic fertilizers on Astragalus membranaceus growth and rhizosphere microbial community. Ann. Microbiol. 2021, 71, 11. [Google Scholar] [CrossRef]
- Rais, A.; Jabeen, Z.; Shair, F.; Hafeez, F.Y.; Hassan, M.N. Bacillus spp., a bio-control agent enhances the activity of antioxidant defense enzymes in rice against Pyricularia oryzae. PLoS ONE 2017, 12, e0187412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.P.; Gaur, R. Endophytic Streptomyces spp. underscore induction of defense regulatory genes and confers resistance against Sclerotium rolfsii in chickpea. Biol. Control 2017, 104, 44–56. [Google Scholar] [CrossRef]
- Dukare, A.; Paul, S. Biological control of Fusarium wilt and growth promotion in pigeon pea (Cajanus cajan) by antagonistic rhizobacteria, displaying multiple modes of pathogen inhibition. Rhizosphere 2021, 17, 100278. [Google Scholar] [CrossRef]
- Jayapala, N.; Mallikarjunaiah, N.; Puttaswamy, H.; Gavirangappa, H.; Ramachandrappa, S. Rhizobacteria Bacillus spp. induce resistance against anthracnose disease in chili (Capsicum annuum L.) through activating host defense response. Egypt J. Biol. Pest Control 2019, 29, 45. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, S.; Santamaria, O.; Halecker, S.; Lledó, S.; Stadler, M. Antagonism between Byssochlamys spectabilis (anamorph Paecilomyces variotii ) and plant pathogens: Involvement of the bioactive compounds produced by the endophyte. Ann. Appl. Biol. 2017, 171, 464–476. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W.H.; Zhou, Y.H.; Yao, S.X.; Deng, L.L.; Zeng, K.F. Isolation, identification and in vitro, screening of chongqing orangery yeasts for the biocontrol of Penicillium digitatum, on citrus fruit. Biol. Control 2017, 110, 18–24. [Google Scholar] [CrossRef]
- Nong, Q.; Zhang, W.L.; Lan, T.j.; Su, Q.; Chen, Y.L.; Zhang, Y.; Qin, L.P.; Xie, L. Screening and identification of dark septate endophyte strain L-14 and its mechanism of banana Fusarium wilt disease resistance. Chin. J. Trop. Crops 2017, 38, 559–564. [Google Scholar]
- Sánchez-Montesinos, B.; Santos, M.; Moreno-Gavíra, A.; Marín-Rodulfo, T.; Gea, F.J.; Diánez, F. Biological control of fungal diseases by Trichoderma aggressivum f. europaeum and its compatibility with fungicides. J. Fungi 2021, 7, 598. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Treatment | Bacteria | Fungi |
|---|---|---|---|
| Tomato | CK | 5317.60 ± 110.67 a | 752.42 ± 37.72 a |
| NCD-2 | 5086.30 ± 115.61 a | 648.89 ± 44.84 b | |
| Eggplant | CK | 5536.00 ± 357.83 a | 787.24 ± 88.72 a |
| NCD-2 | 5287.20 ± 109.10 a | 684.16 ± 55.44 a | |
| Pepper | CK | 4996.70 ± 277.47 a | 724.17 ± 97.82 a |
| NCD-2 | 4922.60 ± 243.39 a | 704.29 ± 18.04 a | |
| Cotton | CK | 5141.90 ± 309.11 a | 565.92 ± 45.41 a |
| NCD-2 | 4792.20 ± 129.30 a | 531.79 ± 25.70 a | |
| Maize | CK | 4292.30 ± 440.07 a | 506.52 ± 72.85 a |
| NCD-2 | 4465.20 ± 45.48 a | 561.22 ± 16.13 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Ban, Y.; Su, Z.; Li, S.; Liu, X.; Guo, Q.; Ma, P. Colonization Ability of Bacillus subtilis NCD-2 in Different Crops and Its Effect on Rhizosphere Microorganisms. Microorganisms 2023, 11, 776. https://doi.org/10.3390/microorganisms11030776
Zhao W, Ban Y, Su Z, Li S, Liu X, Guo Q, Ma P. Colonization Ability of Bacillus subtilis NCD-2 in Different Crops and Its Effect on Rhizosphere Microorganisms. Microorganisms. 2023; 11(3):776. https://doi.org/10.3390/microorganisms11030776
Chicago/Turabian StyleZhao, Weisong, Yiyun Ban, Zhenhe Su, Shezeng Li, Xiaomeng Liu, Qinggang Guo, and Ping Ma. 2023. "Colonization Ability of Bacillus subtilis NCD-2 in Different Crops and Its Effect on Rhizosphere Microorganisms" Microorganisms 11, no. 3: 776. https://doi.org/10.3390/microorganisms11030776