
Microbiome of Citrullus colocynthis (L.) Schrad. Reveals a Potential Association with Non-Photosynthetic Cyanobacteria
 , and
, and 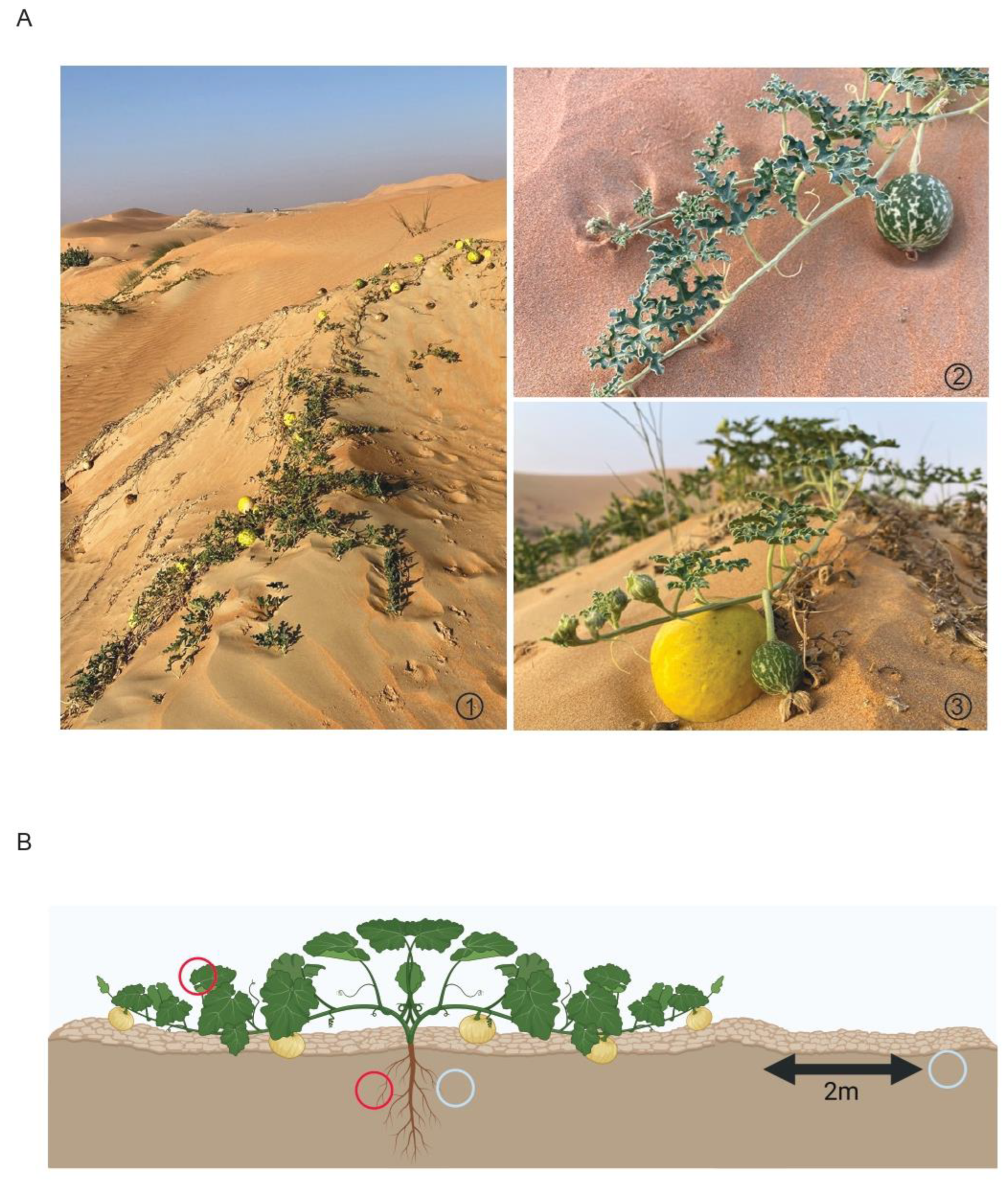{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Isolation and Sequencing
2.3. 16S rRNA Gene based Microbial Profiling
2.4. Soil Composition Analyses
3. Results
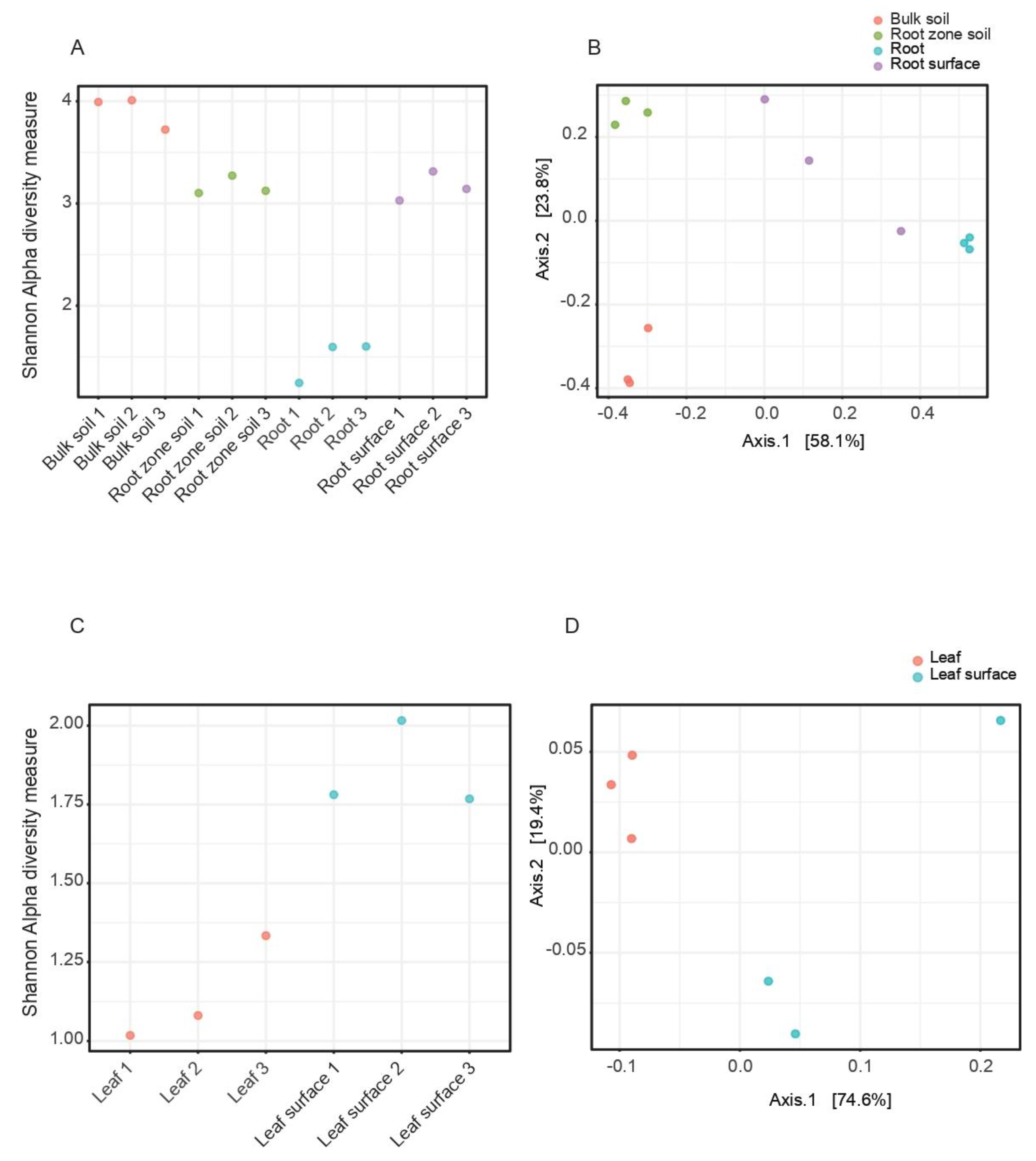
3.1. Species Diversity Indexes
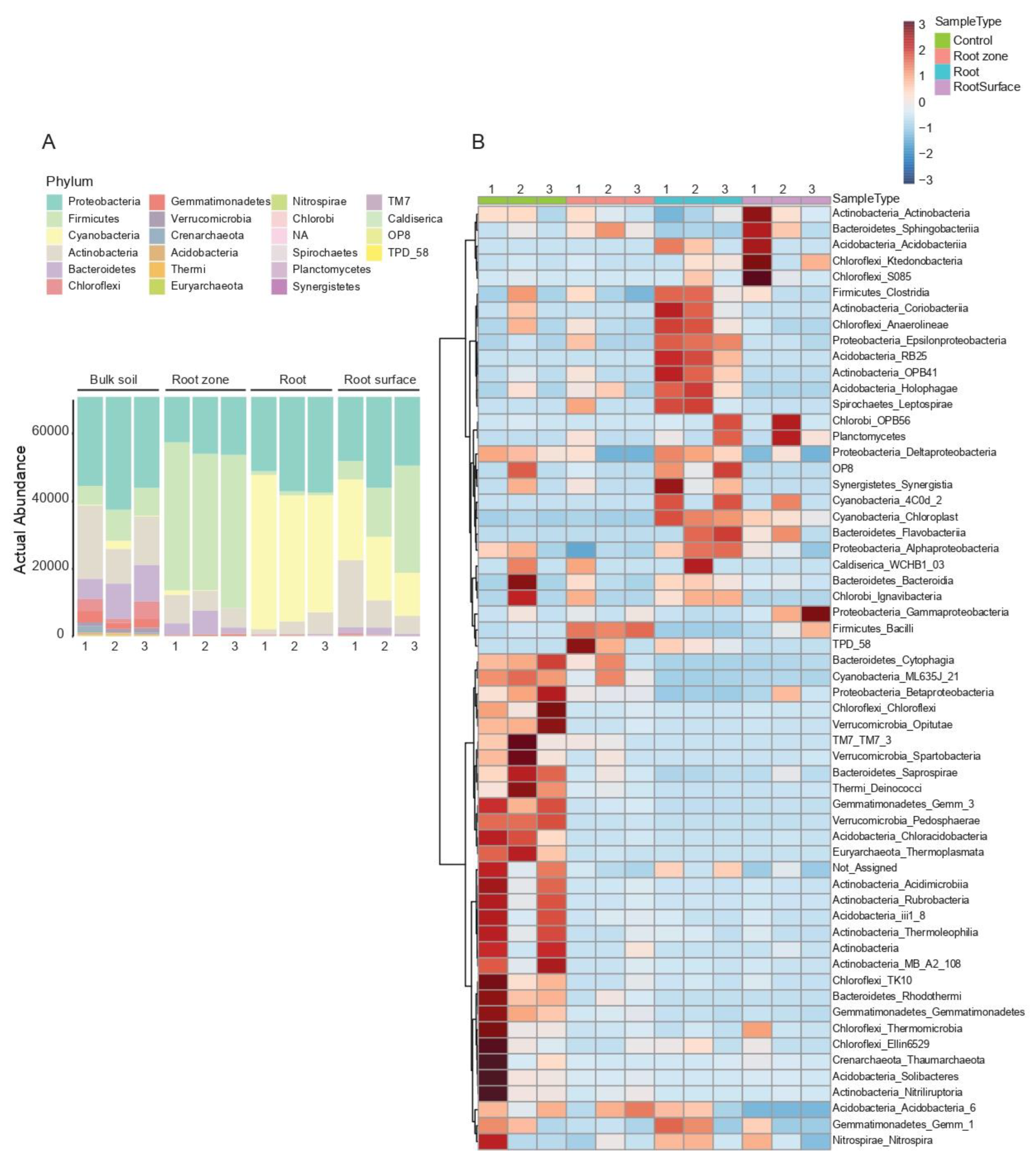
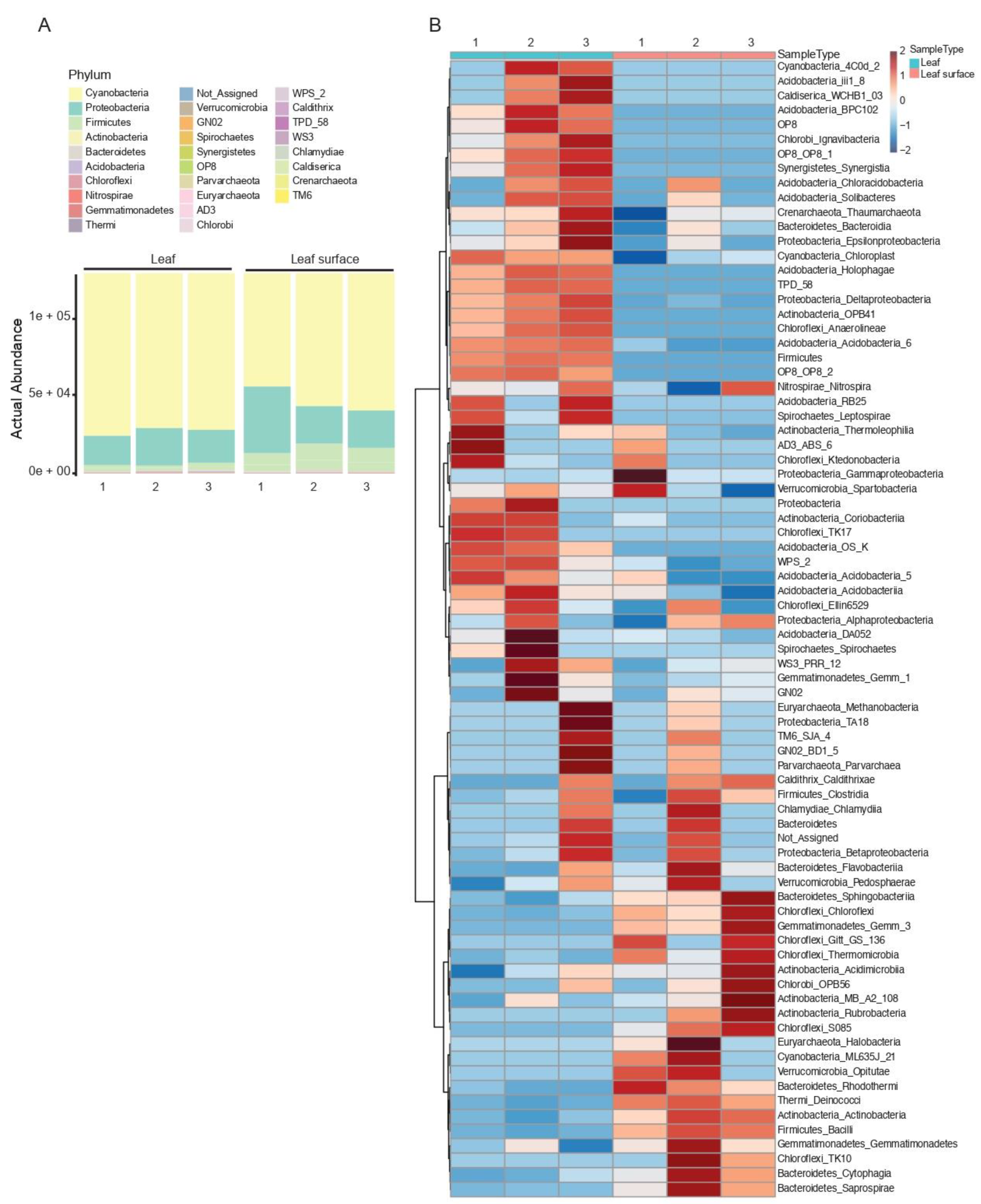
3.2. Taxonomic Composition and Abundance
3.3. Soil Composition Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hegazy, A.K.; Lovett Doust, J. Plant Ecology in the Middle East, 1st ed.; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Drezner, T.D. Desert Ecosystems. In The International Encyclopedia of Geography; Richardson, D., Castree, N., Goodchild, M.F., Kobayashi, A., Liu, W., Marston, R.A., Eds.; Wiley Blackwell: Malden, MA, USA; Oxford, UK; Chichester, UK, 2017; pp. 1–9. [Google Scholar]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makhalanyane, T.P.; Valverde, A.; Gunnigle, E.; Frossard, A.; Ramond, J.-B.; Cowan, D.A. Microbial ecology of hot desert edaphic systems. FEMS Microbiol. Rev. 2015, 39, 203–221. [Google Scholar] [CrossRef]
- Soussi, A.; Ferjani, R.; Marasco, R.; Guesmi, A.; Cherif, H.; Rolli, E.; Mapelli, F.; Ouzari, H.I.; Daffonchio, D.; Cherif, A. Plant-associated microbiomes in arid lands: Diversity, ecology and biotechnological potential. Plant Soil 2016, 405, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.M.; Balint-Kurti, P.; Bede, J.C.; Day, B.; Gold, S.; Goss, E.M.; Grenville-Briggs, L.J.; Jones, K.M.; Wang, A.; Wang, Y.; et al. What are the Top 10 Unanswered Questions in Molecular Plant-Microbe Interactions? Mol. Plant-Microbe Interact. MPMI 2020, 33, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Eida, A.A.; Ziegler, M.; Lafi, F.F.; Michell, C.T.; Voolstra, C.R.; Hirt, H.; Saad, M.M. Desert plant bacteria reveal host influence and beneficial plant growth properties. PLoS ONE 2018, 13, e0208223. [Google Scholar] [CrossRef]
- Alotaibi, M.O.; Sonbol, H.S.; Alwakeel, S.S.; Suliman, R.S.; Fodah, R.A.; Abu Jaffal, A.S.; AlOthman, N.I.; Mohammed, A.E. Microbial Diversity of Some Sabkha and Desert Sites in Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 2778–2789. [Google Scholar] [CrossRef]
- Alsharif, W.; Saad, M.M.; Hirt, H. Desert Microbes for Boosting Sustainable Agriculture in Extreme Environments. Front. Microbiol. 2020, 11, 1666. [Google Scholar] [CrossRef]
- Fuentes, A.; Herrera, H.; Charles, T.C.; Arriagada, C. Fungal and Bacterial Microbiome Associated with the Rhizosphere of Native Plants from the Atacama Desert. Microorganisms 2020, 8, 209. [Google Scholar] [CrossRef] [Green Version]
- Eshel, G.; Araus, V.; Undurraga, S.; Soto, D.C.; Moraga, C.; Montecinos, A.; Moyano, T.; Maldonado, J.; Díaz, F.P.; Varala, K.; et al. Plant ecological genomics at the limits of life in the Atacama Desert. Proc. Natl. Acad. Sci. USA 2021, 118, e2101177118. [Google Scholar] [CrossRef]
- Ha, J.; Gao, Y.; Zhang, R.; Li, K.; Zhang, Y.; Niu, X.; Chen, X.; Luo, K.; Chen, Y. Diversity of the Bacterial Microbiome Associated With the Endosphere and Rhizosphere of Different Cassava (Manihot esculenta Crantz) Genotypes. Front. Microbiol. 2021, 12, 729022. [Google Scholar] [CrossRef] [PubMed]
- Botanical Society of America. Carnivorous Plants/ Insectivorous Plants. Available online: https://botany.org/home/resources/carnivorous-plants-insectivorous-plants.html (accessed on 14 February 2022).
- Beltran-Garcia, M.J.; White, J.F.; Prado, F.M.; Prieto, K.R.; Yamaguchi, L.F.; Torres, M.S.; Kato, M.J.; Medeiros, M.H.G.; Di Mascio, P. Nitrogen acquisition in Agave tequilana from degradation of endophytic bacteria. Sci. Rep. 2014, 4, 6938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.F.; Kingsley, K.L.; Verma, S.K.; Kowalski, K.P. Rhizophagy Cycle: An Oxidative Process in Plants for Nutrient Extraction from Symbiotic Microbes. Microorganisms 2018, 6, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.F.; Torres, M.S.; Somu, M.P.; Johnson, H.; Irizarry, I.; Chen, Q.; Zhang, N.; Walsh, E.; Tadych, M.; Bergen, M. Hydrogen peroxide staining to visualize intracellular bacterial infections of seedling root cells. Microsc. Res. Tech. 2014, 77, 566–573. [Google Scholar] [CrossRef]
- White, J.F. Rhizophagy: Plants Farming Soil Microbes. In GreenCover Soil Resource Guide, 7th ed.; GreenCover, Ed.; Bladen, NE, USA, 2021; Volume 7, pp. 50–51. [Google Scholar]
- Hill, P.W.; Marsden, K.A.; Jones, D.L. How significant to plant N nutrition is the direct consumption of soil microbes by roots? New Phytol. 2013, 199, 948–955. [Google Scholar] [CrossRef] [Green Version]
- White, J.F.; Torres, M.S.; Verma, S.K.; Elmore, M.T.; Kowalski, K.P.; Kingsley, K.L. Evidence for Widespread Microbivory of Endophytic Bacteria in Roots of Vascular Plants Through Oxidative Degradation in Root Cell Periplasmic Spaces. In PGPR Amelioration in Sustainable Agriculture; Singh, A.K., Kumar, A., Singh, P.K., Eds.; Woodhead Publishing: Oxford, UK, 2018; pp. 167–193. [Google Scholar]
- White, J.F.; Torres, M.S. Is plant endophyte-mediated defensive mutualism the result of oxidative stress protection? Physiol. Plant. 2010, 138, 440–446. [Google Scholar] [CrossRef]
- Pravin, B.; Tushar, D.; Vijay, P.; Kishanchnad, K. Review on Citrullus colocynthis. Int. J. Res. Pharm. Chem. 2013, 3, 46–53. [Google Scholar]
- Mariod, A.A.; Saeed Mirghani, M.E.; Hussein, I. Citrullus colocynthis Colocynth, Bitter Apple, Bitter Gourd. In Unconventional Oilseeds and Oil Sources, Mirghani; Mirghani, M.S., Ed.; Elsevier: Boston, MA, USA, 2017; pp. 99–105. [Google Scholar]
- Jongbloed, M.V.D. Comprehensive Guide to the Wild Flowers of the United Arab Emirates, 1st ed.; Environmental Research and Wildlife Development Agency: Abu Dhabi, The United Arab Emirates, 2003; p. 576.
- Shahid, M.; Rao, N.K. Diversity of Citrullus colocynthis (L.) Schrad.(Cucurbitaceae) in the United Arab Emirates. J. New Biol. Rep. 2014, 3, 145–150. [Google Scholar]
- Giwa, S.; Abdullah, L.C.; Adam, N.M. Investigating “Egusi” (Citrullus Colocynthis L.) Seed Oil as Potential Biodiesel Feedstock. Energies 2010, 3, 607–618. [Google Scholar] [CrossRef]
- Greco, M.; Sáez, C.A.; Brown, M.T.; Bitonti, M.B. A simple and effective method for high quality co-extraction of genomic DNA and total RNA from low biomass Ectocarpus siliculosus, the model brown alga. PLoS ONE 2014, 9, e96470. [Google Scholar] [CrossRef] [Green Version]
- Kashi, F.J. An Improved Procedure of the Metagenomic DNA Extraction from Saline Soil, Sediment and Salt. Int. Lett. Nat. Sci. 2014, 60, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: From raw reads to community analyses. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 131. [Google Scholar]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume, F.B.; Roeland, K.; Legendre, P.; Peter, M.; O’Hara, R.B.; Gavin, S.; Peter, S.; Stevenes, M.H.H.; Helene, W. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 14 February 2022).
- Ameen, F.; Al, N.S.; Yassin, M.A.; Al-Sabri, A.; Almansob, A.; Alqahtani, N.; Stephenson, S.L. Desert soil fungi isolated from Saudi Arabia: Cultivable fungal community and biochemical production. Saudi. J. Biol. Sci. 2022, 29, 2409–2420. [Google Scholar] [CrossRef]
- Larbi, A.; Kchaou, H.; Gaaliche, B.; Gargouri, K.; Boulal, H.; Morales, F. Supplementary potassium and calcium improves salt tolerance in olive plants. Sci. Hortic. 2020, 260, 108912. [Google Scholar] [CrossRef]
- Greger, M.; Landberg, T.; Vaculík, M. Silicon Influences Soil Availability and Accumulation of Mineral Nutrients in Various Plant Species. Plants 2018, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The Ecology of Acidobacteria: Moving beyond Genes and Genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielak, A.M.; Cipriano, M.A.P.; Kuramae, E.E. Acidobacteria strains from subdivision 1 act as plant growth-promoting bacteria. Arch. Microbiol. 2016, 198, 987–993. [Google Scholar] [CrossRef]
- Yoneda, Y.; Yamamoto, K.; Makino, A.; Tanaka, Y.; Meng, X.-Y.; Hashimoto, J.; Shin-Ya, K.; Satoh, N.; Fujie, M.; Toyama, T.; et al. Novel Plant-Associated Acidobacteria Promotes Growth of Common Floating Aquatic Plants, Duckweeds. Microorganisms 2021, 9, 1133. [Google Scholar] [CrossRef] [PubMed]
- De Clerck, O.; Bogaert, K.A.; Leliaert, F. Chapter Two—Diversity and Evolution of Algae: Primary Endosymbiosis. In Advances in Botanical Research; Piganeau, G., Ed.; Academic Press: Cambridge, MA, USA, 2012; Volume 64, pp. 55–86. [Google Scholar]
- El-Tarabily, K.A.; Sham, A.; Elbadawi, A.A.; Hassan, A.H.; Alhosani, B.K.K.; El-Esawi, M.A.; AlKhajeh, A.S.; AbuQamar, S.F. A Consortium of Rhizosphere-Competent Actinobacteria Exhibiting Multiple Plant Growth-Promoting Traits Improves the Growth of Avicennia marina in the United Arab Emirates. Front. Mar. Sci. 2021, 8, 715123. [Google Scholar] [CrossRef]
- Soo, R.M.; Skennerton, C.T.; Sekiguchi, Y.; Imelfort, M.; Paech, S.J.; Dennis, P.G.; Steen, J.A.; Parks, D.H.; Tyson, G.W.; Hugenholtz, P. An expanded genomic representation of the phylum cyanobacteria. Genome Biol. Evol. 2014, 6, 1031–1045. [Google Scholar] [CrossRef] [Green Version]
- Soo, R.M.; Hemp, J.; Parks, D.H.; Fischer, W.W.; Hugenholtz, P. On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria. Science 2017, 355, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- McGorum, B.C.; Pirie, R.S.; Glendinning, L.; McLachlan, G.; Metcalf, J.S.; Banack, S.A.; Cox, P.A.; Codd, G.A. Grazing livestock are exposed to terrestrial cyanobacteria. Vet. Res. 2015, 46, 16. [Google Scholar] [CrossRef] [Green Version]
- Monchamp, M.-E.; Spaak, P.; Pomati, F. Long Term Diversity and Distribution of Non-photosynthetic Cyanobacteria in Peri-Alpine Lakes. Front. Microbiol. 2018, 9, 3344. [Google Scholar] [CrossRef] [Green Version]
- Cano-Díaz, C.; Maestre, F.T.; Eldridge, D.J.; Singh, B.K.; Bardgett, R.D.; Fierer, N.; Delgado-Baquerizo, M. Contrasting environmental preferences of photosynthetic and non-photosynthetic soil cyanobacteria across the globe. Glob. Ecol. Biogeogr. 2020, 29, 2025–2038. [Google Scholar] [CrossRef]
- Hu, C.; Rzymski, P. Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health. Life 2022, 12, 476. [Google Scholar] [CrossRef]
- Lee, S.-M.; Ryu, C.-M. Algae as New Kids in the Beneficial Plant Microbiome. Front. Plant Sci. 2021, 12, 599742. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzi, S.C.; Sharon, I.; Wrighton, K.C.; Koren, O.; Hug, L.A.; Thomas, B.C.; Goodrich, J.K.; Bell, J.T.; Spector, T.D.; Banfield, J.F.; et al. The human gut and groundwater harbor non-photosynthetic bacteria belonging to a new candidate phylum sibling to Cyanobacteria. eLife 2013, 2, e01102. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Procter, M.; Kundu, B.; Sudalaimuthuasari, N.; AlMaskari, R.S.; Saeed, E.E.; Hazzouri, K.M.; Amiri, K.M.A. Microbiome of Citrullus colocynthis (L.) Schrad. Reveals a Potential Association with Non-Photosynthetic Cyanobacteria. Microorganisms 2022, 10, 2083. https://doi.org/10.3390/microorganisms10102083
Procter M, Kundu B, Sudalaimuthuasari N, AlMaskari RS, Saeed EE, Hazzouri KM, Amiri KMA. Microbiome of Citrullus colocynthis (L.) Schrad. Reveals a Potential Association with Non-Photosynthetic Cyanobacteria. Microorganisms. 2022; 10(10):2083. https://doi.org/10.3390/microorganisms10102083
Chicago/Turabian StyleProcter, Miranda, Biduth Kundu, Naganeeswaran Sudalaimuthuasari, Raja S. AlMaskari, Esam E. Saeed, Khaled M. Hazzouri, and Khaled M. A. Amiri. 2022. "Microbiome of Citrullus colocynthis (L.) Schrad. Reveals a Potential Association with Non-Photosynthetic Cyanobacteria" Microorganisms 10, no. 10: 2083. https://doi.org/10.3390/microorganisms10102083