
Dynamics of Microbial Community during the Co-Composting of Swine and Poultry Manure with Spent Mushroom Substrates at an Industrial Scale

Abstract
:1. Introduction
2. Materials and Methods
2.1. Composting Materials and Processing
2.2. Sampling
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Physiochemical Analysis
2.5. Statistical Analyses
3. Results and Discussion
3.1. Physiochemical Characteristics of Composts
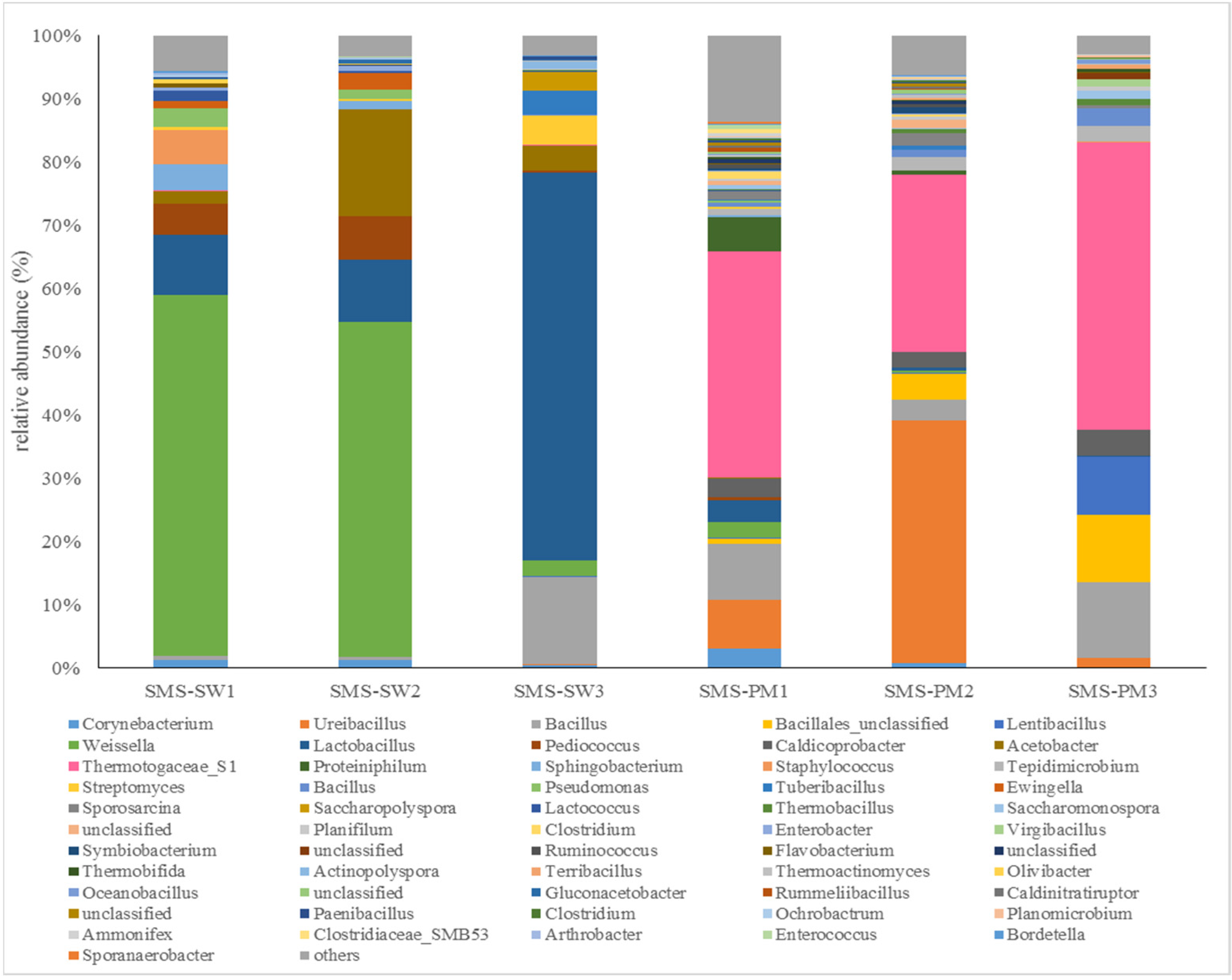
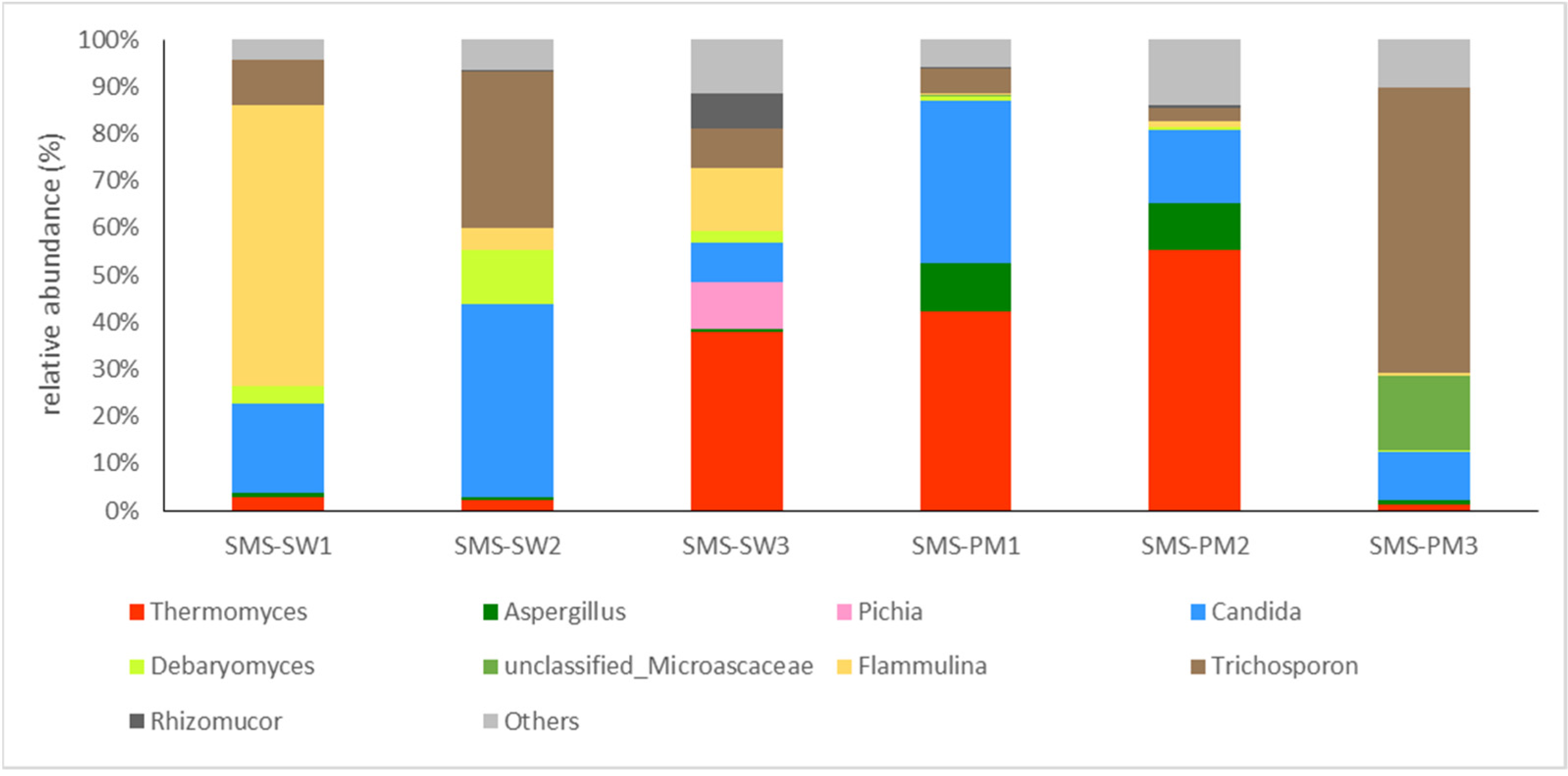
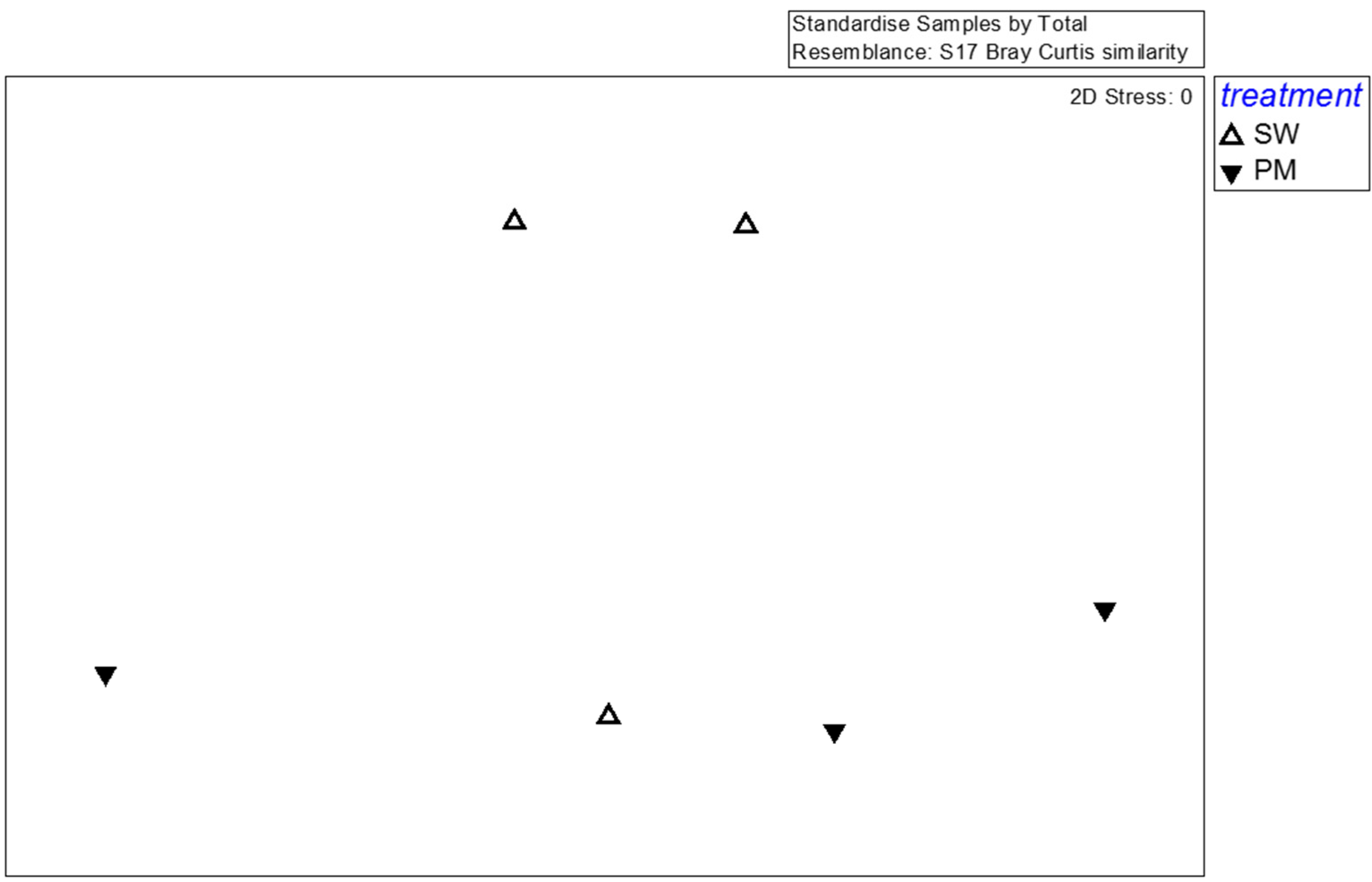
3.2. Bacterial and Fungal Diversity
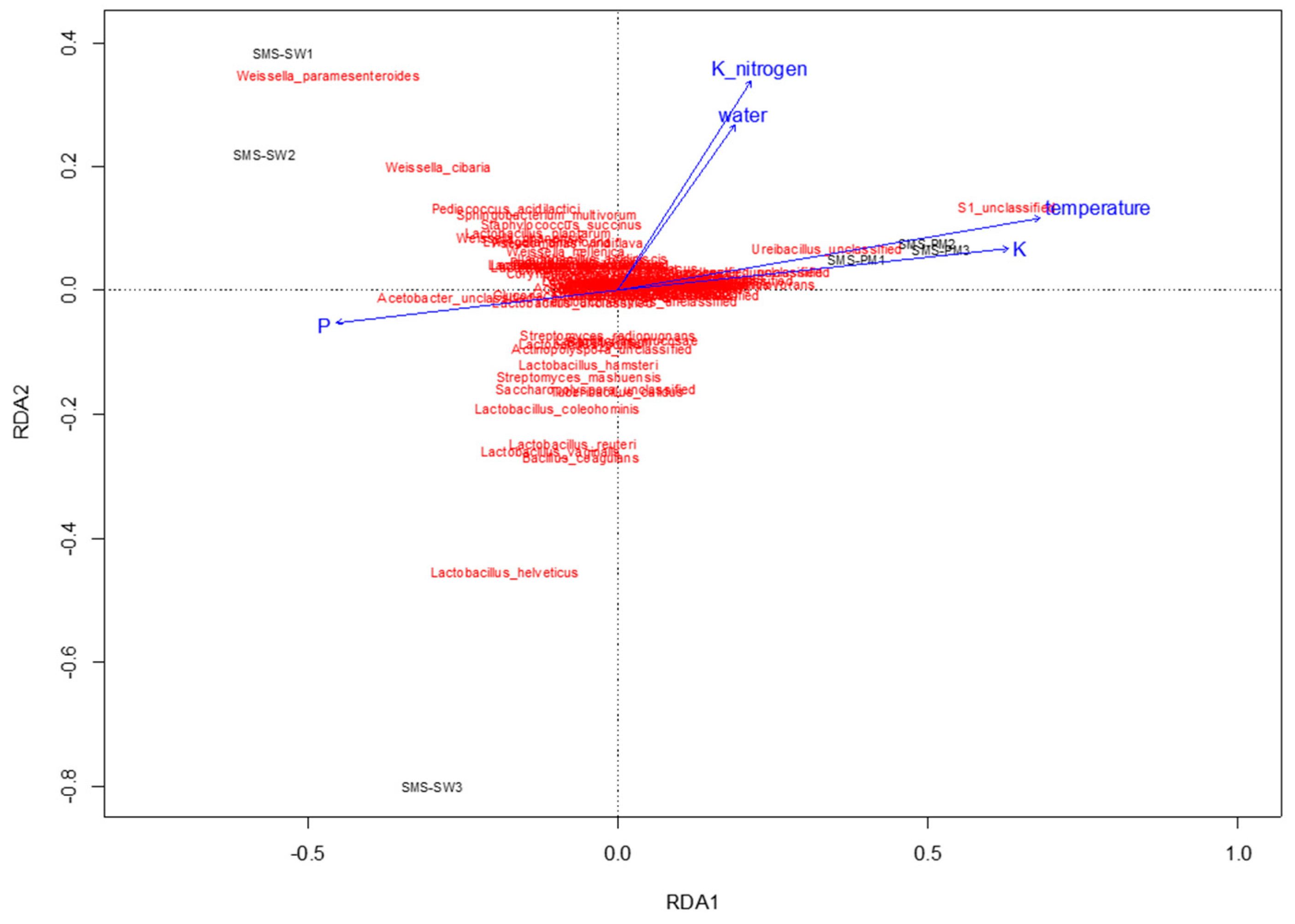
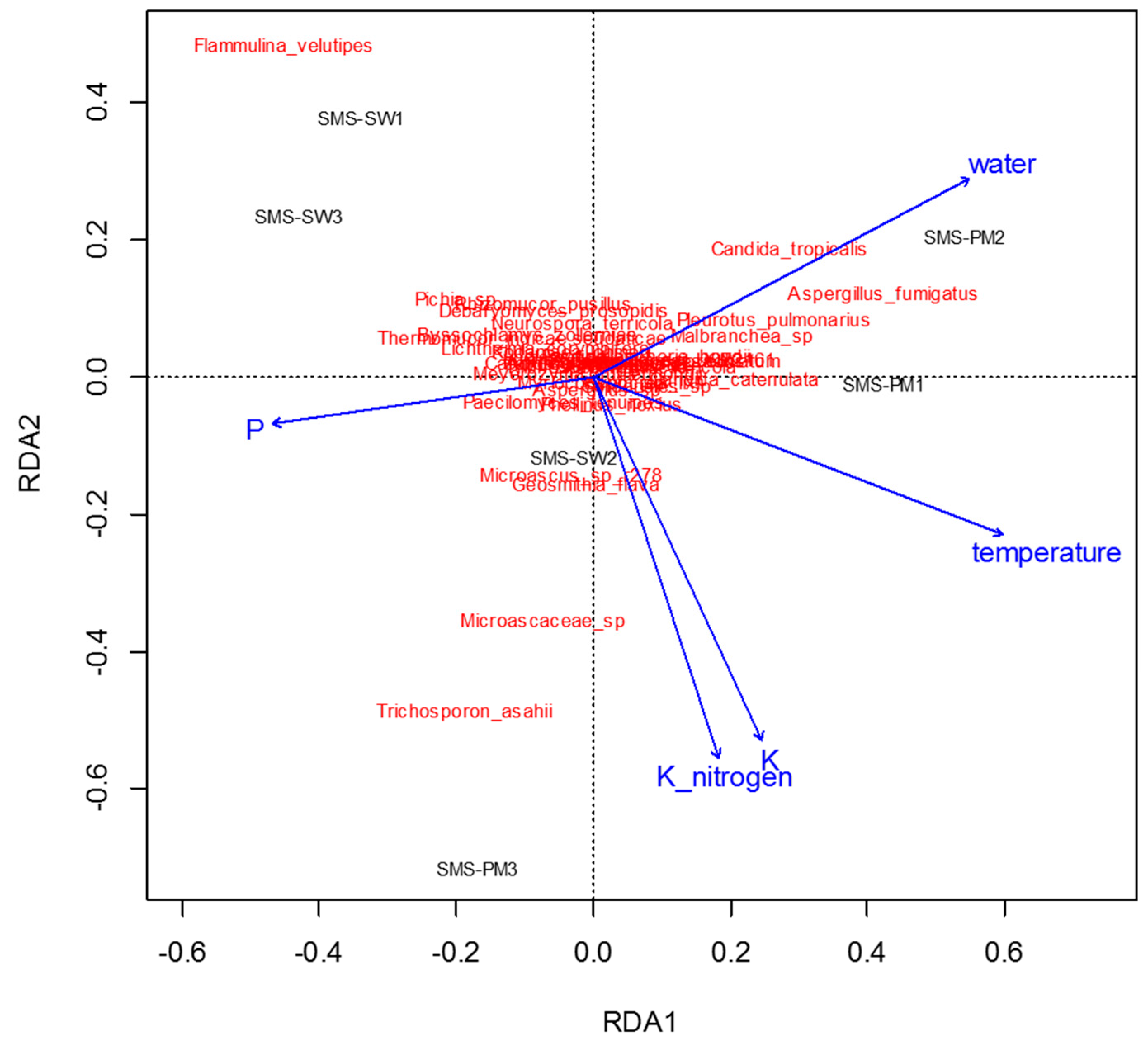
3.3. Correlations between Microbiota and Physiochemical Parameters during Composting
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finney, K.N.; Ryu, C.; Sharifi, V.N.; Swithenbank, J. The reuse of spent mushroom compost and coal tailings for energy recovery: Comparison of thermal treatment technologies. Bioresour. Technol. 2009, 100, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Polat, E.; Uzun, H.; Topçuo, B.; Önal, K.; Onus, A.N. Effects of spent mushroom compost on quality and productivity of cucumber (Cucumis sativus L.) grown in greenhouses. Afr. J. Biotechnol. 2009, 8, 176–180. [Google Scholar]
- Uzun, I. Use of spent mushroom compost in sustainable fruit production. J. Fruit Ornam. Plant Res. 2004, 12, 157–165. [Google Scholar]
- Ayilara, M.S.; Olanrewaju, O.S.; Babalola, O.O.; Odeyemi, O. Waste Management through Composting: Challenges and Potentials. Sustainability 2020, 12, 4456. [Google Scholar] [CrossRef]
- Nakasaki, K.; Hirai, H.; Mimoto, H.; Quyen, T.N.M.; Koyama, M.; Takeda, K. Succession of microbial community during vigorous organic matter degradation in the primary fermentation stage of food waste composting. Sci. Total Environ. 2019, 671, 1237–1244. [Google Scholar] [CrossRef]
- Antunes, L.P.; Martins, L.F.; Pereira, R.V.; Thomas, A.M.; Barbosa, D.; Lemos, L.N.; Silva, G.M.M.; Moura, L.M.S.; Epamino, G.W.C.; Digiampietri, L.; et al. Microbial community structure and dynamics in thermophilic composting viewed through metagenomics and metatranscriptomics. Sci. Rep. 2016, 6, 38915. [Google Scholar] [CrossRef] [Green Version]
- Varma, V.S.; Dhamodharan, K.; Kalamdhad, A.S. Characterization of bacterial community structure during in-vessel composting of agricultural waste by 16S rRNA sequencing. 3 Biotech 2018, 8, 301. [Google Scholar] [CrossRef]
- Xie, X.; Guo, X.; Zhou, L.; Yao, Q.; Zhu, H. Study of Biochemical and Microbiological Properties During Co-composting of Spent Mushroom Substrates and Chicken Feather. Waste Biomass-Valorization 2017, 10, 23–32. [Google Scholar] [CrossRef]
- Herlemann, D.P.R.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Motooka, D.; Fujimoto, K.; Tanaka, R.; Yaguchi, T.; Gotoh, K.; Maeda, Y.; Furuta, Y.; Kurakawa, T.; Goto, N.; Yasunaga, T.; et al. Fungal ITS1 Deep-Sequencing Strategies to Reconstruct the Composition of a 26-Species Community and Evaluation of the Gut Mycobiota of Healthy Japanese Individuals. Front. Microbiol. 2017, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Kalamdhad, A.S.; Singh, Y.K.; Ali, M.; Khwairakpam, M.; Kazmi, A. Rotary drum composting of vegetable waste and tree leaves. Bioresour. Technol. 2009, 100, 6442–6450. [Google Scholar] [CrossRef]
- MacDonald, D.C. Methods of Soil and Tissue Analysis Used in the Analytical Laboratory; Canadian Forestry Service Information Report 1977, MM-X-78; Maritimes Forest Research Centre, Canadian Forestry Service, Department of the Environment: Fredericton, NB, Canada, 1972. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 17th ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Peter Solymos, M.; Stevens, H.H.; Wagner, H. Package ‘Vegan’. Community Ecol. Package 2013, 2, 1–295. [Google Scholar]
- Hu, T.-J.; Zeng, G.-M.; Huang, D.-L.; Yu, H.-Y.; Jiang, X.-Y.; Dai, F.; Huang, G.-H. Use of potassium dihydrogen phosphate and sawdust as adsorbents of ammoniacal nitrogen in aerobic composting process. J. Hazard. Mater. 2007, 141, 736–744. [Google Scholar] [CrossRef]
- Meng, X.; Liu, B.; Xi, C.; Luo, X.; Yuan, X.; Wang, X.; Zhu, W.; Wang, H.; Cui, Z. Effect of pig manure on the chemical composition and microbial diversity during co-composting with spent mushroom substrate and rice husks. Bioresour. Technol. 2017, 251, 22–30. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, X. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar. Bioresour. Technol. 2014, 171, 274–284. [Google Scholar] [CrossRef]
- Garvie, E.I. Genus Leuconostoc. In Bergey’s Manual of Determinative Bacteriology, 8th ed.; Buchanan, R.E., Gibbons, N.E., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1974; pp. 529–550. [Google Scholar]
- Fusco, V.; Quero, G.M.; Cho, G.S.; Kabisch, J.; Meske, D.; Neve, H.; Bockelmann, W.; Franz, C.M. The genus Weissella: Taxonomy, ecology and biotechnological potential. Front. Microbiol. 2015, 6, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qian, M.; Shen, Y.; Qin, X.; Huang, H.; Yang, H.; He, Y.; Bai, W. An high-throughput sequencing approach to the preliminary analysis of bacterial communities associated with changes in amino acid nitrogen, organic acid and reducing sugar contents during soy sauce fermentation. Food Chem. 2021, 349, 129131. [Google Scholar] [CrossRef] [PubMed]
- Tran, Q.N.M.; Mimoto, H.; Koyama, M.; Nakasaki, K. Lactic acid bacteria modulate organic acid production during early stages of food waste composting. Sci. Total Environ. 2019, 687, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, C.; Donders, G.G.; Palmeira-De-Oliveira, R.; Queiroz, J.A.; Tomaz, C.; Martinez-De-Oliveira, J. Bacteriocin production of the probiotic Lactobacillus acidophilus KS400. AMB Express 2018, 8, 153. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, Y.; Hou, Q.; Huang, W.; Zheng, H.; Gao, X.; Yu, J.; Kwok, L.-Y.; Zhang, H.; Sun, Z. Lactobacillus plantarum improves the efficiency of sheep manure composting and the quality of the final product. Bioresour. Technol. 2019, 297, 122456. [Google Scholar] [CrossRef] [PubMed]
- Dahle, H.; Hannisdal, B.; Steinsbu, B.O.; Ommedal, H.; Einen, J.; Jensen, S.; Larsen, O.; Øvreås, L.; Norland, S. Evolution of temperature optimum in Thermotogaceae and the prediction of trait values of uncultured organisms. Extremophiles 2011, 15, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Huber, R.; Langworthy, T.A.; König, H.; Thomm, M.; Woese, C.R.; Sleytr, U.B.; Stetter, K.O. Thermotoga maritima sp. nov. represents a new genus of unique extremely thermophilic eubacteria growing up to 90 ℃. Arch. Microbiol. 1986, 144, 324–333. [Google Scholar] [CrossRef]
- Andersson, M.; Laukkanen, M.; Nurmiaho-Lassila, E.L.; Rainey, F.A.; Niemela, S.I.; Salkinoja-Salonen, M. Bacillus thermosphaericus sp. nov.: A new thermophilic ureolytic bacillus isolated from air. Syst. Appl. Microbiol. 1995, 18, 203–220. [Google Scholar] [CrossRef]
- Fortina, M.G.; Pukall, R.; Schumann, P.; Mora, D.; Parini, C.; Manachini, P.L.; Stackebrandt, E. Ureibacillus gen. nov., a new genus to accommodate Bacillus thermosphaericus (Anderson et al. 1995), emendation of Ureibacillus thermosphaericus and description of Ureibacillus terrenus sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 5, 447–455. [Google Scholar] [CrossRef]
- Kim, B.-Y.; Lee, S.-Y.; Weon, H.-Y.; Kwon, S.-W.; Go, S.-J.; Park, Y.-K.; Schumann, P.; Fritze, D. Ureibacillus suwonensis sp. nov., isolated from cotton waste composts. Int. J. Syst. Evol. Microbiol. 2006, 56, 663–666. [Google Scholar] [CrossRef]
- Wan, L.; Wang, X.; Cong, C.; Li, J.; Xu, Y.; Li, X.; Hou, F.; Wu, Y.; Wang, L. Effect of inoculating microorganisms in chicken manure composting with maize straw. Bioresour. Technol. 2020, 301, 122730. [Google Scholar] [CrossRef]
- Asano, R.; Otawa, K.; Ozutsumi, Y.; Yamamoto, N.; Abdel-Mohsein, H.S.; Nakai, Y. Development and analysis of microbial characteristics of an acidulocomposting system for the treatment of garbage and cattle manure. J. Biosci. Bioeng. 2010, 110, 419–425. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, Y.; Yang, S.; Zhang, W.; Xu, M.; Ma, A.; Zhuang, G.; Chen, G.; Liu, W. Diversity and dynamics of the microbial community on decomposing wheat straw during mushroom compost production. Bioresour. Technol. 2014, 170, 183–195. [Google Scholar] [CrossRef]
- Singh, S.; Madlala, A.M.; Prior, B.A. Thermomyces lanuginosus: Properties of strains and their hemicellulases. FEMS Microbiol. Rev. 2003, 27, 3–16. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Moisture Content (%) | Total Kjeldahl Nitrogen (mg/L) | Total Phosphorus (mg/L) | Total Potassium (mg/L) | Total Organic Carbon (%) | Ammonium Nitrogen (%) | pH | EC (dS/m) | |
|---|---|---|---|---|---|---|---|---|---|
| SMS-SW1 | 38.33 | 48.15 | 46.59 | 28,802.24 | 2.42 | 96.83 | 0.01 | 6.30 | 2.11 |
| SMS-SW2 | 43.50 | 44.38 | 82.85 | 28,969.66 | 2.48 | 95.13 | 0.01 | 6.23 | 2.22 |
| SMS-SW3 | 39.33 | 38.30 | 28.70 | 28,467.42 | 2.49 | 93.40 | 0.10 | 5.93 | 3.04 |
| SMS-PM1 | 56.67 | 56.33 | 64.20 | 27,714.06 | 3.57 | 85.95 | 0.20 | 5.90 | 4.86 |
| SMS-PM2 | 64.50 | 52.63 | 54.82 | 14,823.30 | 2.93 | 87.73 | 0.20 | 6.82 | 2.91 |
| SMS-PM3 | 55.33 | 37.73 | 87.26 | 26,123.64 | 3.99 | 92.35 | 0.01 | 6.01 | 4.79 |
| Species | Average of Relative Abundance (%) | Contribution (%) | Cumulative (%) | ||
|---|---|---|---|---|---|
| SW | PM | ||||
| Bacteria | Thermotogaceae sp. | 0.06 | 41 | 21.38 | 21.38 |
| Weissella paramesenteroides | 26.75 | 0.9 | 13.61 | 34.99 | |
| Ureibacillus sp. | 0.08 | 17.58 | 9.14 | 44.13 | |
| Lactobacillus helveticus | 11.57 | 0.18 | 5.97 | 50.1 | |
| Weissella cibaria | 9.84 | 0.16 | 5.06 | 55.15 | |
| Fungi | Flammulina velutipes | 29.97 | 1.44 | 21.52 | 21.52 |
| Trichosporon asahii | 19.18 | 26.41 | 16.75 | 38.27 | |
| Candida catenulata | 18.15 | 25.58 | 12.9 | 51.17 | |
| Aspergillus fumigatus | 0.65 | 12.34 | 8.95 | 60.13 | |
| Candida tropicalis | 6.56 | 11.5 | 7.47 | 67.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.-R.; Li, H.-Y.; Lin, L.-C.; Hsieh, S.-Y. Dynamics of Microbial Community during the Co-Composting of Swine and Poultry Manure with Spent Mushroom Substrates at an Industrial Scale. Microorganisms 2022, 10, 2064. https://doi.org/10.3390/microorganisms10102064
Lin W-R, Li H-Y, Lin L-C, Hsieh S-Y. Dynamics of Microbial Community during the Co-Composting of Swine and Poultry Manure with Spent Mushroom Substrates at an Industrial Scale. Microorganisms. 2022; 10(10):2064. https://doi.org/10.3390/microorganisms10102064
Chicago/Turabian StyleLin, Wan-Rou, Han-Yun Li, Lei-Chen Lin, and Sung-Yuan Hsieh. 2022. "Dynamics of Microbial Community during the Co-Composting of Swine and Poultry Manure with Spent Mushroom Substrates at an Industrial Scale" Microorganisms 10, no. 10: 2064. https://doi.org/10.3390/microorganisms10102064