
The Changes in Bacterial Microbiome Associated with Immune Disorder in Allergic Respiratory Disease

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
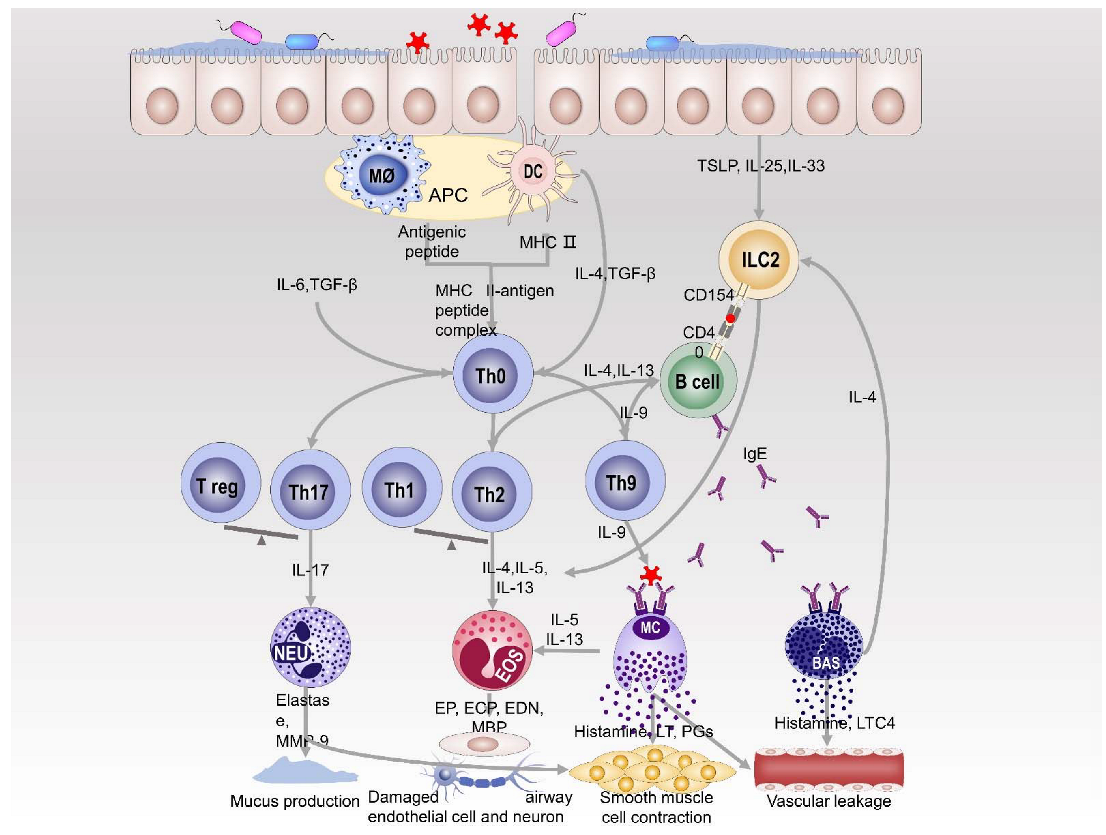
2. Immune Mechanism of Allergic Respiratory Diseases
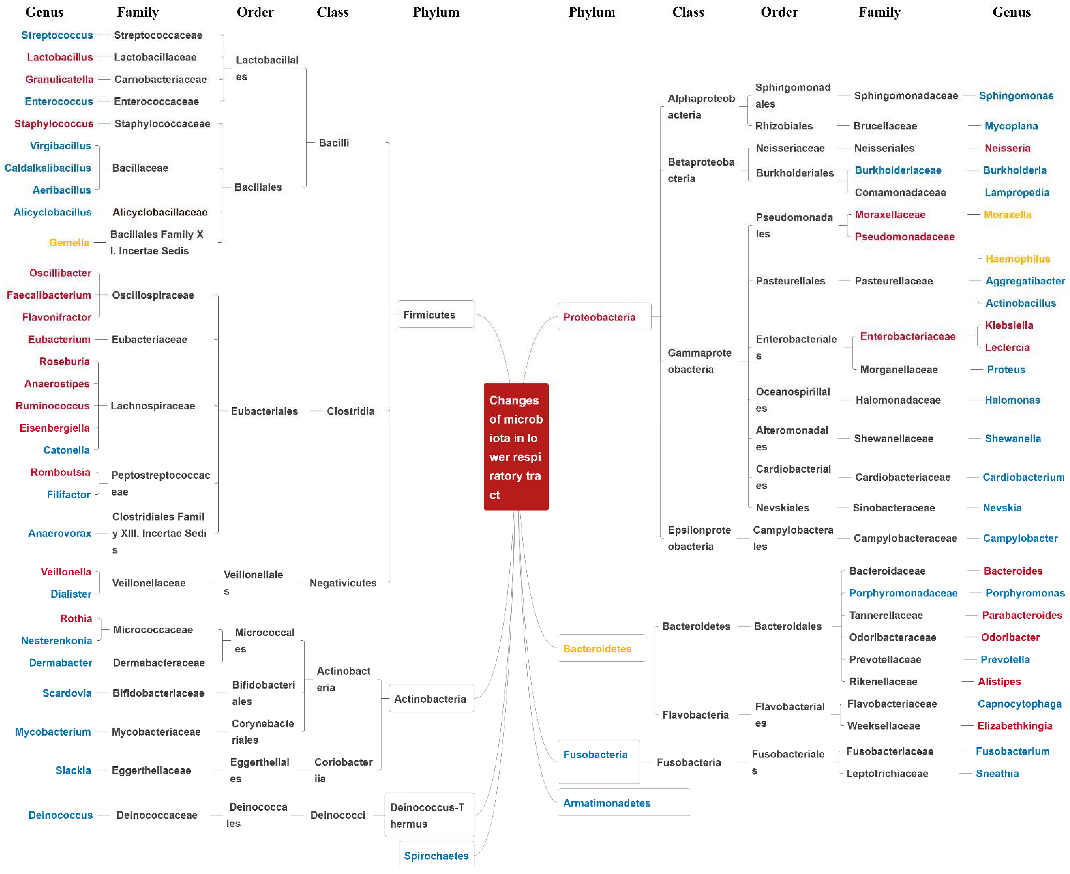
3. Characteristics of Bacterial Microbiome Changes in the Respiratory tract and Intestine
3.1. Bacteria Characteristics of Allergic Respiratory Diseases in the Upper Respiratory Tract and Intestine
3.1.1. Changes in Upper Respiratory Tract Bacteria in Allergic Respiratory Diseases
3.1.2. Changes of Intestinal Bacteria in Allergic Respiratory Diseases
3.2. Changes in the Bacteria of the Lower Respiratory Tract in Asthma
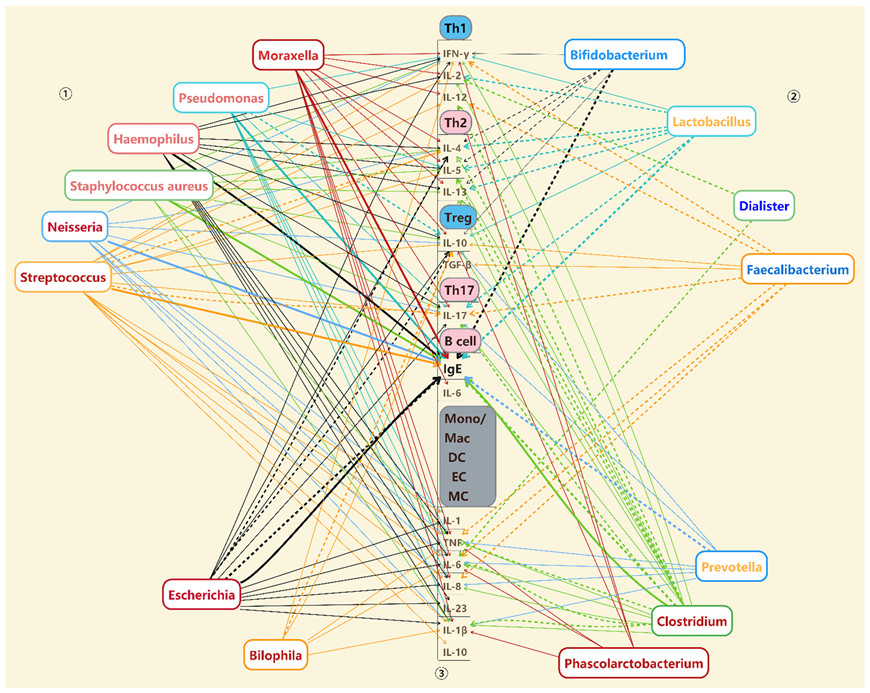
4. The Association between Changed Bacteria and Immune Disorders
5. Treatment
6. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fazlollahi, M.; Lee, T.D.; Andrade, J.; Oguntuyo, K.; Chun, Y.; Grishina, G.; Grishin, A.; Bunyavanich, S. The nasal microbiome in asthma. J. Allergy Clin. Immun. 2018, 142, 834–843.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boeck, I.; Wittouck, S.; Martens, K.; Claes, J.; Jorissen, M.; Steelant, B.; van den Broek, M.F.L.; Seys, S.F.; Hellings, P.W.; Vanderveken, O.M.; et al. Anterior Nares Diversity and Pathobionts Represent Sinus Microbiome in Chronic Rhinosinusitis. Msphere 2019, 4, e00532-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marazzato, M.; Zicari, A.M.; Aleandri, M.; Conte, A.L.; Longhi, C.; Vitanza, L.; Bolognino, V.; Zagaglia, C.; De Castro, G.; Brindisi, G.; et al. 16S Metagenomics Reveals Dysbiosis of Nasal Core Microbiota in Children With Chronic Nasal Inflammation: Role of Adenoid Hypertrophy and Allergic Rhinitis. Front. Cell. Infect. Microbiol. 2020, 10, 458. [Google Scholar] [CrossRef] [PubMed]
- Rup, L. The Human Microbiome Project. Indian J. Microbiol. 2012, 52, 315. [Google Scholar] [CrossRef] [Green Version]
- Eifan, A.O.; Durham, S.R. Pathogenesis of rhinitis. Clin. Exp. Allergy 2016, 46, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Schleimer, R.P. Immunopathogenesis of Chronic Rhinosinusitis and Nasal Polyposis. Annu. Rev. Pathol. Mech. 2017, 12, 331–357. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.D.; Vercelli, D. Asthma. Lancet 2013, 382, 1360–1372. [Google Scholar] [CrossRef]
- Koch, S.; Sopel, N.; Finotto, S. Th9 and other IL-9-producing cells in allergic asthma. Semin. Immunopathol. 2017, 39, 55–68. [Google Scholar] [CrossRef]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4+T Cells: Differentiation and Functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [Green Version]
- Scadding, G.K.; Kariyawasam, H.H.; Scadding, G.; Mirakian, R.; Buckley, R.J.; Dixon, T.; Durham, S.R.; Farooque, S.; Jones, N.; Leech, S.; et al. BSACI guideline for the diagnosis and management of allergic and non-allergic rhinitis (Revised Edition 2017; First edition 2007). Clin. Exp. Allergy 2017, 47, 856–889. [Google Scholar] [CrossRef]
- Bloemen, K.; Verstraelen, S.; Van Den Heuvel, R.; Witters, H.; Nelissen, I.; Schoeters, G. The allergic cascade: Review of the most important molecules in the asthmatic lung. Immunol. Lett. 2007, 113, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Barcik, W.; Boutin, R.C.T.; Sokolowska, M.; Finlay, B.B. The Role of Lung and Gut Microbiota in the Pathology of Asthma. Immunity 2020, 52, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef] [PubMed]
- Maggi, L.; Montaini, G.; Mazzoni, A.; Rossettini, B.; Capone, M.; Rossi, M.C.; Santarlasci, V.; Liotta, F.; Rossi, O.; Gallo, O.; et al. Human circulating group 2 innate lymphoid cells can express CD154 and promote IgE production. J. Allergy Clin. Immunol. 2017, 139, 964–976.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alnahas, S.; Hagner, S.; Raifer, H.; Kilic, A.; Gasteiger, G.; Mutters, R.; Hellhund, A.; Prinz, I.; Pinkenburg, O.; Visekruna, A.; et al. IL-17 and TNF-α Are Key Mediators of Moraxella catarrhalis Triggered Exacerbation of Allergic Airway Inflammation. Front. Immunol. 2017, 8, 1562. [Google Scholar] [CrossRef] [Green Version]
- Larsen, J.M.; Musavian, H.S.; Butt, T.M.; Ingvorsen, C.; Thysen, A.H.; Brix, S. Chronic obstructive pulmonary disease and asthma-associated Proteobacteria, but not commensal Prevotella spp., promote Toll-like receptor 2-independent lung inflammation and pathology. Immunology 2015, 144, 333–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ta, L.D.H.; Yap, G.C.; Tay, C.J.X.; Lim, A.S.M.; Huang, C.H.; Chu, C.W.; De Sessions, P.F.; Shek, L.P.; Goh, A.; Van Bever, H.P.S.; et al. Establishment of the nasal microbiota in the first 18 months of life: Correlation with early-onset rhinitis and wheezing. J. Allergy Clin. Immunol. 2018, 142, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Copeland, E.; Leonard, K.; Carney, R.; Kong, J.; Forer, M.; Naidoo, Y.; Oliver, B.G.G.; Seymour, J.R.; Woodcock, S.; Burke, C.M.; et al. Chronic Rhinosinusitis: Potential Role of Microbial Dysbiosis and Recommendations for Sampling Sites. Front. Cell. Infect. Microbiol. 2018, 8, 57. [Google Scholar] [CrossRef]
- Hyun, D.W.; Min, H.J.; Kim, M.S.; Whon, T.W.; Shin, N.R.; Kim, P.S.; Kim, H.S.; Lee, J.Y.; Kang, W.; Choi, A.M.K.; et al. Dysbiosis of Inferior Turbinate Microbiota Is Associated with High Total IgE Levels in Patients with Allergic Rhinitis. Infect. Immun. 2018, 86, e00934-17. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.B.; Hong, S.W.; Kim, D.K.; Jeon, S.G.; Kim, K.R.; Cho, S.H.; Gho, Y.S.; Jee, Y.K.; Kim, Y.K. Decreased diversity of nasal microbiota and their secreted extracellular vesicles in patients with chronic rhinosinusitis based on a metagenomic analysis. Allergy 2014, 69, 517–526. [Google Scholar] [CrossRef]
- Kim, B.S.; Lee, E.; Lee, M.J.; Kang, M.J.; Yoon, J.; Cho, H.J.; Park, J.; Won, S.; Lee, S.Y.; Hong, S.J. Different functional genes of upper airway microbiome associated with natural course of childhood asthma. Allergy 2018, 73, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Gan, W.G.; Yang, F.J.; Tang, Y.P.; Zhou, D.M.; Qing, D.; Hu, J.J.; Liu, S.X.; Liu, F.; Meng, J. The difference in nasal bacterial microbiome diversity between chronic rhinosinusitis patients with polyps and a control population. Int. Forum Allergy Rhinol. 2019, 9, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Gan, W.G.; Zhang, H.T.; Yang, F.J.; Liu, S.X.; Liu, F.; Meng, J. The influence of nasal microbiome diversity and inflammatory patterns on the prognosis of nasal polyps. Sci. Rep. 2021, 11, 6364. [Google Scholar] [CrossRef] [PubMed]
- Gan, W.G.; Zhang, H.T.; Yang, F.J.; Liu, S.X.; Liu, F.; Meng, J. The influence of nasal bacterial microbiome diversity on the pathogenesis and prognosis of chronic rhinosinusitis patients with polyps. Eur. Arch. Oto-Rhino-Laryngol. 2021, 278, 1075–1088. [Google Scholar] [CrossRef]
- Cleland, E.J.; Bassiouni, A.; Vreugde, S.; Wormald, P.J. The bacterial microbiome in chronic rhinosinusitis: Richness, diversity, postoperative changes, and patient outcomes. Am. J. Rhinol. Allergy 2016, 30, 37–43. [Google Scholar] [CrossRef]
- Gan, W.G.; Yang, F.J.; Meng, J.; Liu, F.; Liu, S.X.; Xian, J.M. Comparing the nasal bacterial microbiome diversity of allergic rhinitis, chronic rhinosinusitis and control subjects. Eur. Arch. Oto-Rhino-Laryngol. 2021, 278, 711–718. [Google Scholar] [CrossRef]
- Park, S.C.; Park, I.; Lee, J.S.; Park, S.M.; Kang, S.H.; Hong, S.M.; Byun, S.H.; Jung, Y.G.; Hong, S.J. Microbiome of Unilateral Chronic Rhinosinusitis: A Controlled Paired Analysis. Int. J. Environ. Res. Public Health 2021, 18, 9878. [Google Scholar] [CrossRef]
- Biswas, K.; Mackenzie, B.W.; Waldvogel-Thurlow, S.; Middleditch, M.; Jullig, M.; Zoing, M.; Taylor, M.W.; Douglas, R.G. Differentially Regulated Host Proteins Associated with Chronic Rhinosinusitis Are Correlated with the Sinonasal Microbiome. Front. Cell. Infect. Microbiol. 2017, 7, 504. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, K.; Negm, H.; Ralli, M.; Passali, D. The theory of a “staphylococcus superantigen” in chronic rhinosinusitis with nasal polyps: Myth or reality? Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 48–54. [Google Scholar]
- Kim, J.H.; Kim, S.H.; Lim, J.Y.; Kim, D.; Jeong, I.S.; Lee, D.K.; Jang, Y.J. Association between the sinus microbiota with eosinophilic inflammation and prognosis in chronic rhinosinusitis with nasal polyps. Exp. Mol. Med. 2020, 52, 978–987. [Google Scholar] [CrossRef]
- Abbas, E.E.; Li, C.; Xie, A.; Lu, S.; Tang, L.; Liu, Y.H.; Elfadil, A.; Wen, S. Distinct Clinical Pathology and Microbiota in Chronic Rhinosinusitis With Nasal Polyps Endotypes. Laryngoscope 2021, 131, E34–E44. [Google Scholar] [CrossRef] [PubMed]
- Aurora, R.; Chatterjee, D.; Hentzleman, J.; Prasad, G.; Sindwani, R.; Sanford, T. Contrasting the Microbiomes From Healthy Volunteers and Patients With Chronic Rhinosinusitis. JAMA Otolaryngol. 2013, 139, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Kim, D.Y.; Choi, S.; Won, S.; Kang, H.R.; Yi, H. Microbiome profiling of uncinate tissue and nasal polyps in patients with chronic rhinosinusitis using swab and tissue biopsy. PLoS ONE 2021, 16, e0249688. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chan, Y.L.; Tsai, Y.S.; Chen, S.A.; Wang, C.J.; Chen, K.F.; Chung, I.F. Airway Microbial Diversity is Inversely Associated with Mite-Sensitized Rhinitis and Asthma in Early Childhood. Sci. Rep. 2017, 7, 1820. [Google Scholar] [CrossRef] [Green Version]
- Hoggard, M.; Biswas, K.; Zoing, M.; Mackenzie, B.W.; Taylor, M.W.; Douglas, R.G. Evidence of microbiota dysbiosis in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2017, 7, 230–239. [Google Scholar] [CrossRef]
- Mansbach, J.M.; Luna, P.N.; Shaw, C.A.; Hasegawa, K.; Petrosino, J.F.; Piedra, P.A.; Sullivan, A.F.; Espinola, J.A.; Stewart, C.J.; Camargo, C.A. Increased Moraxella and Streptococcus species abundance after severe bronchiolitis is associated with recurrent wheezing. J. Allergy Clin. Immunol. 2020, 145, 518–527.e8. [Google Scholar] [CrossRef] [Green Version]
- Depner, M.; Ege, M.J.; Cox, M.J.; Dwyer, S.; Walker, A.W.; Birzele, L.T.; Genuneit, J.; Horak, E.; Braun-Fahrlander, C.; Danielewicz, H.; et al. Bacterial microbiota of the upper respiratory tract and childhood asthma. J. Allergy Clin. Immunol. 2017, 139, 826–834.e13. [Google Scholar] [CrossRef] [Green Version]
- Maniakas, A.; Asmar, M.H.; Flores, A.E.R.; Nayan, S.; Alromaih, S.; Endam, L.M.; Desrosiers, M.Y. Staphylococcus aureus on Sinus Culture Is Associated With Recurrence of Chronic Rhinosinusitis After Endoscopic Sinus Surgery. Front. Cell. Infect. Microbiol. 2018, 8, 150. [Google Scholar] [CrossRef] [Green Version]
- Lal, D.; Keim, P.; Delisle, J.; Barker, B.; Rank, M.A.; Chia, N.; Schupp, J.M.; Gillece, J.D.; Cope, E.K. Mapping and comparing bacterial microbiota in the sinonasal cavity of healthy, allergic rhinitis, and chronic rhinosinusitis subjects. Int. Forum Allergy Rhinol. 2017, 7, 561–569. [Google Scholar] [CrossRef]
- Wei, H.Z.; Li, Y.C.; Wang, X.D.; Lu, X.X.; Hu, C.H.; He, S.; Liu, X. The microbiology of chronic rhinosinusitis with and without nasal polyps. Eur. Arch. Oto-Rhino-Laryngol. 2018, 275, 1439–1447. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chan, Y.L.; Tsai, M.H.; Wang, C.J.; Chiang, M.H.; Chiu, C.C.; Su, S.C. Cross-talk between airway and gut microbiome links to IgE responses to house dust mites in childhood airway allergies. Sci. Rep. 2020, 10, 13449. [Google Scholar] [CrossRef]
- Morin, A.; McKennan, C.G.; Pedersen, C.E.T.; Stokholm, J.; Chawes, B.L.; Schoos, A.M.M.; Naughton, K.A.; Thorsen, J.; Mortensen, M.S.; Vercelli, D.; et al. Epigenetic landscape links upper airway microbiota in infancy with allergic rhinitis at 6 years of age. J. Allergy Clin. Immunol. 2020, 146, 1358–1366. [Google Scholar] [CrossRef]
- Chun, Y.; Do, A.; Grishina, G.; Grishin, A.; Fang, G.; Rose, S.; Spencer, C.; Vicencio, A.; Schadt, E.; Bunyavanich, S. Integrative study of the upper and lower airway microbiome and transcriptome in asthma. JCI Insight 2020, 5, e133707. [Google Scholar] [CrossRef]
- Feng, T.T.; Miao, P.; Liu, B.; Liu, Y.; Bao, X.M.; Xu, J.; Ren, N.; Li, Y.; Shi, J.L.; Cao, W.X.; et al. Sinus Microbiota in Patients With Eosinophilic and Non-Eosinophilic Chronic Rhinosinusitis With Nasal Polyps. Front. Cell. Infect. Microbiol. 2021, 11, 672355. [Google Scholar] [CrossRef]
- Hoggard, M.; Nocera, A.; Biswas, K.; Taylor, M.W.; Douglas, R.G.; Bleier, B.S. The sinonasal microbiota, neural signaling, and depression in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2018, 8, 394–405. [Google Scholar] [CrossRef]
- Park, I.H.; Lee, J.S.; Park, J.H.; Kang, S.H.; Hong, S.M.; Park, I.S.; Yoon, J.H.; Hong, S.J. Comparison of the human microbiome in adults and children with chronic rhinosinusitis. PLoS ONE 2020, 15, e0242770. [Google Scholar] [CrossRef] [PubMed]
- Bachert, C.; Holtappels, G.; Merabishvili, M.; Meyer, T.; Murr, A.; Zhang, N.; Van Crombruggen, K.; Gevaert, E.; Volker, U.; Broker, B.M.; et al. Staphylococcus aureus controls interleukin-5 release in upper airway inflammation. J. Proteom. 2018, 180, 53–60. [Google Scholar] [CrossRef]
- Chalermwatanachai, T.; Vilchez-Vargas, R.; Holtappels, G.; Lacoere, T.; Jauregui, R.; Kerckhof, F.M.; Pieper, D.H.; Van de Wiele, T.; Vaneechoutte, M.; Van Zele, T.; et al. Chronic rhinosinusitis with nasal polyps is characterized by dysbacteriosis of the nasal microbiota. Sci. Rep. 2018, 8, 7926. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.B.; Xu, F.; Wan, W.R.; Yu, B.; Tang, L.; Yang, Y.M.; Du, Y.L.; Chen, Z.R.; Xu, H.Z. Gut microbial characteristics of adult patients with allergy rhinitis. Microb. Cell Fact. 2020, 19, 171. [Google Scholar] [CrossRef]
- Liu, X.; Tao, J.; Li, J.; Cao, X.L.; Li, Y.; Gao, X.F.; Fu, Y. Dysbiosis of Fecal Microbiota in Allergic Rhinitis Patients. Am. J. Rhinol. Allergy 2020, 34, 650–660. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chan, Y.L.; Tsai, M.H.; Wang, C.J.; Chiang, M.H.; Chiu, C.C. Gut microbial dysbiosis is associated with allergen-specific IgE responses in young children with airway allergies. World Allergy Organ. J. 2019, 12, 100021. [Google Scholar] [CrossRef]
- Wang, Q.; Li, F.; Liang, B.S.; Liang, Y.H.; Chen, S.J.; Mo, X.D.; Ju, Y.M.; Zhao, H.; Jia, H.J.; Spector, T.D.; et al. A metagenome-wide association study of gut microbiota in asthma in UK adults. BMC Microbiol. 2018, 18, 114. [Google Scholar] [CrossRef] [Green Version]
- Pang, W.H.; Jiang, Y.; Li, A.F.; Zhang, J.S.; Chen, M.; Hu, L.; Li, Z.Y.; Wang, D.H. Bacteroides thetaiotaomicron Ameliorates Experimental Allergic Airway Inflammation via Activation of ICOS(+)Tregs and Inhibition of Th2 Response. Front. Immunol. 2021, 12, 620943. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Arevalo, A.; Stiemsma, L.; Dimitriu, P.; Chico, M.E.; Loor, S.; Vaca, M.; Boutin, R.C.T.; Morien, E.; Jin, M.L.; et al. Associations between infant fungal and bacterial dysbiosis and childhood atopic wheeze in a nonindustrialized setting. J. Allergy Clin. Immunol. 2018, 142, 424–434.e10. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.Y.; Cheng, M.L.; Chiang, M.H.; Kuo, Y.L.; Tsai, M.H.; Chiu, C.C.; Lin, G. Gut microbial-derived butyrate is inversely associated with IgE responses to allergens in childhood asthma. Pediatr. Allergy Immunol. 2019, 30, 689–697. [Google Scholar] [CrossRef]
- Zhang, L.W.; Wan, Y.; Ma, L.; Xu, K.H.; Cheng, B.J. A low abundance of Bifidobacterium but not Lactobacillius in the feces of Chinese children with wheezing diseases. Medicine 2018, 97, e12745. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Lai, Z.D.; Zhang, X.X.; Huang, P.K.; Xie, J.X.; Jiang, Q.; Zhang, Q.L.; Chung, K.F. Altered gut microbiome compositions are associated with the severity of asthma. J. Thorac. Dis. 2021, 13, 4322–4338. [Google Scholar] [CrossRef]
- Watts, A.M.; West, N.P.; Zhang, P.; Smith, P.K.; Cripps, A.W.; Cox, A.J. The Gut Microbiome of Adults with Allergic Rhinitis Is Characterised by Reduced Diversity and an Altered Abundance of Key Microbial Taxa Compared to Controls. Int. Arch. Allergy Immunol. 2021, 182, 94–105. [Google Scholar] [CrossRef]
- Lee-Sarwar, K.A.; Kelly, R.S.; Lasky-Su, J.; Zeiger, R.S.; O’Connor, G.T.; Sandel, M.T.; Bacharier, L.B.; Beigelman, A.; Laranjo, N.; Gold, D.R.; et al. Integrative analysis of the intestinal metabolome of childhood asthma. J. Allergy Clin. Immunol. 2019, 144, 442–454. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.X.; Li, T.; Yuan, H.Q.; Pan, W.; Dai, Q.G. Correlations of Inflammatory Factors with Intestinal Flora and Gastrointestinal Incommensurate Symptoms in Children with Asthma. Med. Sci. Monitor 2018, 24, 7975–7979. [Google Scholar] [CrossRef]
- Stiemsma, L.T.; Arrieta, M.C.; Dimitriu, P.A.; Cheng, J.; Thorson, L.; Lefebvre, D.L.; Azad, M.B.; Subbarao, P.; Mandhane, P.; Becker, A.; et al. Shifts in Lachnospira and Clostridium sp in the 3-month stool microbiome are associated with preschool age asthma. Clin. Sci. 2016, 130, 2199–2207. [Google Scholar] [CrossRef]
- Demirci, M.; Tokman, H.B.; Uysal, H.K.; Demiryas, S.; Karakullukcu, A.; Saribas, S.; Cokugras, H.; Kocazeybek, B.S. Reduced Akkermansia muciniphila and Faecalibacterium prausnitzii levels in the gut microbiota of children with allergic asthma. Allergol. Immunopathol. 2019, 47, 365–371. [Google Scholar] [CrossRef]
- Zou, X.L.; Wu, J.J.; Ye, H.X.; Feng, D.Y.; Meng, P.; Yang, H.L.; Wu, W.B.; Li, H.T.; He, Z.; Zhang, T.T. Associations Between Gut Microbiota and Asthma Endotypes: A Cross-Sectional Study in South China Based on Patients with Newly Diagnosed Asthma. J. Asthma Allergy 2021, 14, 981–992. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Cox, M.; Liang, Z.K.; Brinkmann, F.; Cardenas, P.A.; Duff, R.; Bhavsar, P.; Cookson, W.; Moffatt, M.; Chung, K.F. Airway Microbiota in Severe Asthma and Relationship to Asthma Severity and Phenotypes. PLoS ONE 2016, 11, e0152724. [Google Scholar] [CrossRef] [Green Version]
- Hilty, M.; Burke, C.; Pedro, H.; Cardenas, P.; Bush, A.; Bossley, C.; Davies, J.; Ervine, A.; Poulter, L.; Pachter, L.; et al. Disordered Microbial Communities in Asthmatic Airways. PLoS ONE 2010, 5, e8578. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.Y.; Li, C.X.; Liang, Z.Y.; Zhang, S.Y.; Yang, W.Y.; Ye, Y.M.; Lin, Y.X.; Chen, R.C.; Zhou, H.W.; Su, J. The Interactions of Airway Bacterial and Fungal Communities in Clinically Stable Asthma. Front. Microbiol. 2020, 11, 1647. [Google Scholar] [CrossRef]
- Goldman, D.L.; Chen, Z.G.; Shankar, V.; Tyberg, M.; Vicencio, A.; Burk, R. Lower airway microbiota and mycobiota in children with severe asthma. J. Allergy Clin. Immunol. 2018, 141, 808–811. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.Q.; Wang, G.Q.; Gibson, P.; Guan, X.W.; Zhang, W.J.; Zheng, R.P.; Chen, F.; Wang, Z.Y.; Wang, F. Airway Microbiome in Different Inflammatory Phenotypes of Asthma: A Cross-Sectional Study in Northeast China. Int. J. Med. Sci. 2019, 16, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Li, N.J.; Qiu, R.H.; Yang, Z.W.; Li, J.; Chung, K.F.; Zhong, N.S.; Zhang, Q.L. Sputum microbiota in severe asthma patients: Relationship to eosinophilic inflammation. Respir. Med. 2017, 131, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Sverrild, A.; Kiilerich, P.; Brejnrod, A.; Pedersen, R.; Porsbjerg, C.; Bergqvist, A.; Erjefalt, J.S.; Kristiansen, K.; Backer, V. Eosinophilic airway inflammation in asthmatic patients is associated with an altered airway microbiome. J. Allergy Clin. Immunol. 2017, 140, 407–417.e11. [Google Scholar] [CrossRef]
- Taylor, S.L.; Leong, L.E.X.; Choo, J.M.; Wesselingh, S.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Jenkins, C.; et al. Inflammatory phenotypes in patients with severe asthma are associated with distinct airway microbiology. J. Allergy Clin. Immunol. 2018, 141, 94–103.e15. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, M.I.; Brinkman, P.; Vijverberg, S.J.H.; Neerincx, A.H.; Riley, J.H.; Bates, S.; Hashimoto, S.; Kermani, N.Z.; Chung, K.F.; Djukanovic, R.; et al. Sputum microbiome profiles identify severe asthma phenotypes of relative stability at 12 to 18 months. J. Allergy Clin. Immunol. 2021, 147, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Shen, C.; Chen, Y.; Yang, X.; Luo, X.; Hang, C.; Yan, L.; Xu, X. Follow-up study of airway microbiota in children with persistent wheezing. Respir Res 2021, 22, 213. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Nariya, S.; Harris, J.M.; Lynch, S.V.; Choy, D.F.; Arron, J.R.; Boushey, H. The airway microbiome in patients with severe asthma: Associations with disease features and severity. J. Allergy Clin. Immunol. 2015, 136, 874–884. [Google Scholar] [CrossRef] [Green Version]
- Durack, J.; Christian, L.S.; Nariya, S.; Gonzalez, J.; Bhakta, N.R.; Ansel, K.M.; Beigelman, A.; Castro, M.; Dyer, A.M.; Israel, E.; et al. Distinct associations of sputum and oral microbiota with atopic, immunologic, and clinical features in mild asthma. J. Allergy Clin. Immunol. 2020, 146, 1016–1026. [Google Scholar] [CrossRef]
- Ren, D.; Xie, H.; Zhang, W.; Hassan, F.; Petralia, R.S.; Yu, S.; Lim, D.J.; Gu, X.X. Intranasal immunization of the combined lipooligosaccharide conjugates protects mice from the challenges with three serotypes of Moraxella catarrhalis. PLoS ONE 2011, 6, e29553. [Google Scholar] [CrossRef] [Green Version]
- Vissing, N.H.; Larsen, J.M.; Rasmussen, M.A.; Chawes, B.L.; Thysen, A.H.; Bønnelykke, K.; Brix, S.; Bisgaard, H. Susceptibility to Lower Respiratory Infections in Childhood is Associated with Perturbation of the Cytokine Response to Pathogenic Airway Bacteria. Pediatr. Infect. Dis. J. 2016, 35, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Schoos, A.M.; Larsen, J.M.; Brix, S.; Thysen, A.H.; Rasmussen, M.A.; Stokholm, J.; Bønnelykke, K.; Bisgaard, H.; Chawes, B.L. Reduced IL-2 response from peripheral blood mononuclear cells exposed to bacteria at 6 months of age is associated with elevated total-IgE and allergic rhinitis during the first 7 years of life. EBioMedicine 2019, 43, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Nordström, T.; Jendholm, J.; Samuelsson, M.; Forsgren, A.; Riesbeck, K. The IgD-binding domain of the Moraxella IgD-binding protein MID (MID962-1200) activates human B cells in the presence of T cell cytokines. J. Leukoc. Biol. 2006, 79, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Gu, X.X. Moraxella catarrhalis lipooligosaccharide selectively upregulates ICAM-1 expression on human monocytes and stimulates adjacent naïve monocytes to produce TNF-alpha through cellular cross-talk. Cell Microbiol. 2008, 10, 1453–1467. [Google Scholar] [CrossRef]
- Slevogt, H.; Schmeck, B.; Jonatat, C.; Zahlten, J.; Beermann, W.; van Laak, V.; Opitz, B.; Dietel, S.; N’Guessan, P.D.; Hippenstiel, S.; et al. Moraxella catarrhalis induces inflammatory response of bronchial epithelial cells via MAPK and NF-kappaB activation and histone deacetylase activity reduction. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L818–L826. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Zimmermann, C.; Mausberg, A.K.; Dehmel, T.; Kieseier, B.C.; Hartung, H.P.; Hofstetter, H.H. Pseudomonas aeruginosa and Its Bacterial Components Influence the Cytokine Response in Thymocytes and Splenocytes. Infect. Immun. 2016, 84, 1413–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiringer, K.; Treis, A.; Fucik, P.; Gona, M.; Gruber, S.; Renner, S.; Dehlink, E.; Nachbaur, E.; Horak, F.; Jaksch, P.; et al. A Th17- and Th2-skewed cytokine profile in cystic fibrosis lungs represents a potential risk factor for Pseudomonas aeruginosa infection. Am. J. Respir. Crit. Care Med. 2013, 187, 621–629. [Google Scholar] [CrossRef]
- Weaver, A.J., Jr.; Brandenburg, K.S.; Smith, B.W.; Leung, K.P. Comparative Analysis of the Host Response in a Rat Model of Deep-Partial and Full-Thickness Burn Wounds With Pseudomonas aeruginosa Infection. Front. Cell. Infect. Microbiol. 2019, 9, 466. [Google Scholar] [CrossRef] [PubMed]
- Murali, P.S.; Pathial, K.; Saff, R.H.; Splaingard, M.L.; Atluru, D.; Kurup, V.P.; Fink, J.N. Immune responses to Aspergillus fumigatus and Pseudomonas aeruginosa antigens in cystic fibrosis and allergic bronchopulmonary aspergillosis. Chest 1994, 106, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, S.; Hayes, D., Jr.; Wozniak, D.J. Mucoid Pseudomonas aeruginosa and regional inflammation in the cystic fibrosis lung. J. Cyst. Fibros. 2019, 18, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Pickering, J.L.; Prosser, A.; Corscadden, K.J.; de Gier, C.; Richmond, P.C.; Zhang, G.; Thornton, R.B.; Kirkham, L.A. Haemophilus haemolyticus Interaction with Host Cells Is Different to Nontypeable Haemophilus influenzae and Prevents NTHi Association with Epithelial Cells. Front. Cell. Infect. Microbiol. 2016, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cai, R.J.; Song, S.; Jiang, Z.Y.; Li, Y.; Gou, H.C.; Chu, P.P.; Li, C.L.; Qiu, H.J. Evaluation of immunogenicity and protective efficacy of recombinant outer membrane proteins of Haemophilus parasuis serovar 5 in a murine model. PLoS ONE 2017, 12, e0176537. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Feng, S.; He, X.; Zhou, Q.; Wang, Y.; Yue, H.; Tang, C.; Zhang, B. Surface-exposed loops L7 and L8 of Haemophilus (Glaesserella) parasuis OmpP2 contribute to the expression of proinflammatory cytokines in porcine alveolar macrophages. Vet. Res. 2019, 50, 105. [Google Scholar] [CrossRef] [Green Version]
- Otczyk, D.C.; Clancy, R.L.; Cripps, A.W. Haemophilus influenzae and smoking-related obstructive airways disease. Int. J. Chronic Obstr. Pulm. Dis. 2011, 6, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Stentzel, S.; Teufelberger, A.; Nordengrün, M.; Kolata, J.; Schmidt, F.; van Crombruggen, K.; Michalik, S.; Kumpfmüller, J.; Tischer, S.; Schweder, T.; et al. Staphylococcal serine protease-like proteins are pacemakers of allergic airway reactions to Staphylococcus aureus. J. Allergy Clin. Immunol. 2017, 139, 492–500.e498. [Google Scholar] [CrossRef] [PubMed]
- Kolata, J.B.; Kühbandner, I.; Link, C.; Normann, N.; Vu, C.H.; Steil, L.; Weidenmaier, C.; Bröker, B.M. The Fall of a Dogma? Unexpected High T-Cell Memory Response to Staphylococcus aureus in Humans. J. Infect. Dis. 2015, 212, 830–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.F.; Murphy, A.G.; Lalor, S.J.; Leech, J.M.; O’Keeffe, K.M.; Mac Aogáin, M.; O’Halloran, D.P.; Lacey, K.A.; Tavakol, M.; Hearnden, C.H.; et al. Memory Th1 Cells Are Protective in Invasive Staphylococcus aureus Infection. PLoS Pathog. 2015, 11, e1005226. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Hattori, H.; Yoshino, T.; Sugata, Y.; Yamamoto, M.; Fujiwara, T.; Satoskar, A.A.; Satoskar, A.R.; Nishizaki, K. Nasal exposure to Staphylococcal enterotoxin enhances the development of allergic rhinitis in mice. Clin. Exp. Allergy 2005, 35, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Oftung, F.; Korsvold, G.E.; Aase, A.; Næss, L.M. Cellular Immune Responses in Humans Induced by Two Serogroup B Meningococcal Outer Membrane Vesicle Vaccines Given Separately and in Combination. Clin. Vaccine Immunol. 2016, 23, 353–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Hammer, L.A.; Liu, W.; Hobbs, M.M.; Zielke, R.A.; Sikora, A.E.; Jerse, A.E.; Egilmez, N.K.; Russell, M.W. Experimental vaccine induces Th1-driven immune responses and resistance to Neisseria gonorrhoeae infection in a murine model. Mucosal Immunol. 2017, 10, 1594–1608. [Google Scholar] [CrossRef] [Green Version]
- Ashton, F.E.; Vijay, H.M.; Lavergne, G.; Brodeur, B.R.; Diena, B.B. Induction of reaginic (IgE) gonococcal antibodies in the rat by a common antigen of Neisseria gonorrhoeae. Can. J. Microbiol. 1979, 25, 138–145. [Google Scholar] [CrossRef]
- Johswich, K. Innate immune recognition and inflammation in Neisseria meningitidis infection. Pathog. Dis. 2017, 75, ftx022. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.; Shen, C.; Ding, M.; Reinhart, T.A.; Chen, Y.; Sankapal, S.; Gupta, P. Neisseria gonorrhoeae uses cellular proteins CXCL10 and IL8 to enhance HIV-1 transmission across cervical mucosa. Am. J. Reprod. Immunol. 2019, 81, e13111. [Google Scholar] [CrossRef] [Green Version]
- Zughaier, S.M. Neisseria meningitidis capsular polysaccharides induce inflammatory responses via TLR2 and TLR4-MD-2. J. Leukoc. Biol. 2011, 89, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Botelho, R.M.; Tenorio, L.P.G.; Silva, A.L.M.; Tanabe, E.L.L.; Pires, K.S.N.; Gonçalves, C.M.; Santos, J.C.; Marques, A.L.X.; Allard, M.J.; Bergeron, J.D.; et al. Biomechanical and functional properties of trophoblast cells exposed to Group B Streptococcus in vitro and the beneficial effects of uvaol treatment. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1417–1428. [Google Scholar] [CrossRef]
- Santos, M.; Castro, H.H.O.; Magalhôes, L.M.D.; Dutra, W.O.; Gollob, K.J.; Guimarães, N.R.; Magalhães, P.P.; de Macêdo Farias, L.; Horta, M.C.R.; Souza, P.E.A. Resinous adhesive systems differentially affect the expression of cytokines by human monocytes stimulated or not with Streptococcus mutans in vitro. Arch. Oral Biol. 2020, 111, 104641. [Google Scholar] [CrossRef]
- Dargahi, N.; Matsoukas, J.; Apostolopoulos, V. Streptococcus thermophilus ST285 Alters Pro-Inflammatory to Anti-Inflammatory Cytokine Secretion against Multiple Sclerosis Peptide in Mice. Brain Sci. 2020, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Engen, S.A.; Valen Rukke, H.; Becattini, S.; Jarrossay, D.; Blix, I.J.; Petersen, F.C.; Sallusto, F.; Schenck, K. The oral commensal Streptococcus mitis shows a mixed memory Th cell signature that is similar to and cross-reactive with Streptococcus pneumoniae. PLoS ONE 2014, 9, e104306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.Y.; Fu, F.; Kong, W.N.; Xuan, Q.K.; Wen, D.H.; Chen, X.Q.; He, Y.M.; He, L.H.; Guo, J.; Zhou, A.P.; et al. Streptococcus agalactiae Inhibits Candida albicans Hyphal Development and Diminishes Host Vaginal Mucosal TH17 Response. Front. Microbiol. 2018, 9, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjaergard, L.L.; Larsen, F.O.; Norn, S.; Clementsen, P.; Skov, P.S.; Permin, H. Basophil-bound IgE and serum IgE directed against Haemophilus influenzae and Streptococcus pneumoniae in patients with chronic bronchitis during acute exacerbations. Apmis 1996, 104, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Almada, G.; Haro, C.; Vintiñi, E.; Medina, M. Safety of a nasal vaccine against Streptococcus pneumoniae using heat-killed Lactobacillus casei as adjuvant. Immunobiology 2015, 220, 109–116. [Google Scholar] [CrossRef]
- van den Bogert, B.; Meijerink, M.; Zoetendal, E.G.; Wells, J.M.; Kleerebezem, M. Immunomodulatory properties of Streptococcus and Veillonella isolates from the human small intestine microbiota. PLoS ONE 2014, 9, e114277. [Google Scholar] [CrossRef]
- Kaushal, N.; Kumari, S.; Jhelum, H.; Sehgal, D. In vitro and in vivo characterization of the interaction, proinflammatory, immunomodulatory and antigenic properties of capsular polysaccharide from Streptococcus pneumoniae serotype 1. Int. J. Biol. Macromol. 2020, 143, 521–532. [Google Scholar] [CrossRef]
- Iwabuchi, N.; Takahashi, N.; Xiao, J.Z.; Miyaji, K.; Iwatsuki, K. In vitro Th1 cytokine-independent Th2 suppressive effects of bifidobacteria. Microbiol. Immunol. 2007, 51, 649–660. [Google Scholar] [CrossRef]
- López, P.; González-Rodríguez, I.; Sánchez, B.; Gueimonde, M.; Margolles, A.; Suárez, A. Treg-inducing membrane vesicles from Bifidobacterium bifidum LMG13195 as potential adjuvants in immunotherapy. Vaccine 2012, 30, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.G.; Kang, G.D.; Kim, H.I.; Han, M.J.; Kim, D.H. Bifidobacterium longum IM55 and Lactobacillus plantarum IM76 alleviate allergic rhinitis in mice by restoring Th2/Treg imbalance and gut microbiota disturbance. Benef. Microbes 2019, 10, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Hacini-Rachinel, F.; Gosoniu, M.L.; Bourdeau, T.; Holvoet, S.; Doucet-Ladeveze, R.; Beaumont, M.; Mercenier, A.; Nutten, S. Immune-modulatory effect of probiotic Bifidobacterium lactis NCC2818 in individuals suffering from seasonal allergic rhinitis to grass pollen: An exploratory, randomized, placebo-controlled clinical trial. Eur. J. Clin. Nutr. 2013, 67, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Kim, W.K.; Han, D.H.; Lee, K.; Ko, G. Lactobacillus fermentum species ameliorate dextran sulfate sodium-induced colitis by regulating the immune response and altering gut microbiota. Gut Microbes 2019, 10, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, L.; Sun, T.; Guo, K.; Geng, S. Dysbiosis of gut microbiota and its correlation with dysregulation of cytokines in psoriasis patients. BMC Microbiol. 2021, 21, 78. [Google Scholar] [CrossRef] [PubMed]
- Martínez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A.; et al. Gut microbiome composition is linked to whole grain-induced immunological improvements. ISME J. 2013, 7, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Jeong, Y.; Kang, S.; You, H.J.; Ji, G.E. Co-Culture with Bifidobacterium catenulatum Improves the Growth, Gut Colonization, and Butyrate Production of Faecalibacterium prausnitzii: In Vitro and In Vivo Studies. Microorganisms 2020, 8, 788. [Google Scholar] [CrossRef]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.L.; Rainteau, D.; Bridonneau, C.; da Costa, G.; van Hylckama Vlieg, J.; Sovran, B.; Chamignon, C.; et al. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat. Commun. 2018, 9, 2802. [Google Scholar] [CrossRef] [Green Version]
- Pang, B.; Jin, H.; Liao, N.; Li, J.; Jiang, C.; Shao, D.; Shi, J. Lactobacillus rhamnosus from human breast milk ameliorates ulcerative colitis in mice via gut microbiota modulation. Food Funct. 2021, 12, 5171–5186. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Wei, H.; Xu, C.; Xie, X.; Jiang, S.; Peng, J. Maternal Soluble Fiber Diet during Pregnancy Changes the Intestinal Microbiota, Improves Growth Performance, and Reduces Intestinal Permeability in Piglets. Appl. Environ. Microbiol. 2018, 84, e01047-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Han, Y.; Cai, X.; Gu, M.; Sun, J.; Qi, C.; Goulette, T.; Song, M.; Li, Z.; Xiao, H. Dietary resveratrol attenuated colitis and modulated gut microbiota in dextran sulfate sodium-treated mice. Food Funct. 2020, 11, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bian, G.; Sun, D.; Zhu, W.; Mao, S. Starter Feeding Supplementation Alters Colonic Mucosal Bacterial Communities and Modulates Mucosal Immune Homeostasis in Newborn Lambs. Front. Microbiol. 2017, 8, 429. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Jiang, Y.; Zhang, B.; Pang, J.; Xu, X.; Sun, J.; Lv, X.; Cai, Q. Comparison of the Modulatory Effect on Intestinal Microbiota between Raw and Bran-Fried Atractylodis Rhizoma in the Rat Model of Spleen-Deficiency Syndrome. Int. J. Environ. Res. Public Health 2019, 16, 3183. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Cao, J.; Yu, L.; Ma, H. Dehydroepiandrosterone resisted E. coli O157:H7-induced inflammation via blocking the activation of p38 MAPK and NF-κB pathways in mice. Cytokine 2020, 127, 154955. [Google Scholar] [CrossRef]
- Brereton, C.F.; Sutton, C.E.; Ross, P.J.; Iwakura, Y.; Pizza, M.; Rappuoli, R.; Lavelle, E.C.; Mills, K.H. Escherichia coli heat-labile enterotoxin promotes protective Th17 responses against infection by driving innate IL-1 and IL-23 production. J. Immunol. 2011, 186, 5896–5906. [Google Scholar] [CrossRef] [Green Version]
- Pang, W.; Wang, H.; Shi, L.; Sun, Y.; Wang, X.; Wang, M.; Li, J.; Wang, H.; Shi, G. Immunomodulatory effects of Escherichia coli ATCC 25922 on allergic airway inflammation in a mouse model. PLoS ONE 2013, 8, e59174. [Google Scholar] [CrossRef] [Green Version]
- Kadafi, K.T.; Wibowo, S. Differences in systemic humoral immune response among Balb/c mice administered with probiotic, LPS Escherichia coli, and probiotic-LPS E. coli. Iran J. Microbiol. 2019, 11, 294–299. [Google Scholar]
- Bruballa, A.C.; Shiromizu, C.M.; Bernal, A.M.; Pineda, G.E.; Sabbione, F.; Trevani, A.S.; Bentancor, L.V.; Ramos, M.V.; Fernández-Brando, R.J.; Muñoz, M.J.; et al. Role of Shiga Toxins in Cytotoxicity and Immunomodulatory Effects of Escherichia coli O157:H7 during Host-Bacterial Interactions in vitro. Toxins 2020, 12, 48. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Tang, J.; Cai, Z.; Zhou, K.; Chang, L.; Bai, Y.; Ma, Y. Prevotella Induces the Production of Th17 Cells in the Colon of Mice. J Immunol. Res. 2020, 2020, 9607328. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chou, H.C.; Chang, L.C.; Fan, W.L.; Dinh, M.C.V.; Kuo, Y.L.; Chung, W.H.; Lai, H.C.; Hsieh, W.P.; Su, S.C. Integration of metagenomics-metabolomics reveals specific signatures and functions of airway microbiota in mite-sensitized childhood asthma. Allergy 2020, 75, 2846–2857. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, Z.; Zeng, M.; Wei, C.; We, H.; Liu, M.; Huang, J.; Zeng, Y.; Sun, X. Oral administration of Clostridium butyricum CGMCC0313-1 reduces ovalbumin-induced allergic airway inflammation in mice. Respirology 2017, 22, 898–904. [Google Scholar] [CrossRef]
- Yu, H.; Chen, K.; Sun, Y.; Carter, M.; Garey, K.W.; Savidge, T.C.; Devaraj, S.; Tessier, M.E.; von Rosenvinge, E.C.; Kelly, C.P.; et al. Cytokines Are Markers of the Clostridium difficile-Induced Inflammatory Response and Predict Disease Severity. Clin. Vaccine Immunol. 2017, 24, e00037-17. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.B.; Jia, L.; Yan, Q.Q.; Deng, Q.; Wei, B. Effect of Clostridium butyricum and Butyrate on Intestinal Barrier Functions: Study of a Rat Model of Severe Acute Pancreatitis With Intra-Abdominal Hypertension. Front. Physiol. 2020, 11, 561061. [Google Scholar] [CrossRef]
- Adlerberth, I.; Strachan, D.P.; Matricardi, P.M.; Ahrné, S.; Orfei, L.; Aberg, N.; Perkin, M.R.; Tripodi, S.; Hesselmar, B.; Saalman, R.; et al. Gut microbiota and development of atopic eczema in 3 European birth cohorts. J. Allergy Clin. Immunol. 2007, 120, 343–350. [Google Scholar] [CrossRef]
- Willers, S.M.; Wijga, A.H.; Brunekreef, B.; Scholtens, S.; Postma, D.S.; Kerkhof, M.; de Jongste, J.C.; Smit, H.A. Childhood diet and asthma and atopy at 8 years of age: The PIAMA birth cohort study. Eur. Respir. J. 2011, 37, 1060–1067. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Shi, L.; Pang, W.; Liu, W.; Li, J.; Wang, H.; Shi, G. Dietary Fiber Intake Regulates Intestinal Microflora and Inhibits Ovalbumin-Induced Allergic Airway Inflammation in a Mouse Model. PLoS ONE 2016, 11, e0147778. [Google Scholar] [CrossRef] [Green Version]
- De Boeck, I.; van den Broek, M.F.L.; Allonsius, C.N.; Spacova, I.; Wittouck, S.; Martens, K.; Wuyts, S.; Cauwenberghs, E.; Jokicevic, K.; Vandenheuvel, D.; et al. Lactobacilli Have a Niche in the Human Nose. Cell Rep. 2020, 31, 107674. [Google Scholar] [CrossRef]
- Wei, X.; Jiang, P.; Liu, J.; Sun, R.; Zhu, L. Association between probiotic supplementation and asthma incidence in infants: A meta-analysis of randomized controlled trials. J. Asthma 2020, 57, 167–178. [Google Scholar] [CrossRef]
- Du, X.; Wang, L.; Wu, S.; Yuan, L.; Tang, S.; Xiang, Y.; Qu, X.; Liu, H.; Qin, X.; Liu, C. Efficacy of probiotic supplementary therapy for asthma, allergic rhinitis, and wheeze: A meta-analysis of randomized controlled trials. Allergy Asthma Proc. 2019, 40, 250–260. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, J.; Kou, F.; Men, X.; Liu, Y.; Tang, L.; Wen, S. The Changes in Bacterial Microbiome Associated with Immune Disorder in Allergic Respiratory Disease. Microorganisms 2022, 10, 2066. https://doi.org/10.3390/microorganisms10102066
Lyu J, Kou F, Men X, Liu Y, Tang L, Wen S. The Changes in Bacterial Microbiome Associated with Immune Disorder in Allergic Respiratory Disease. Microorganisms. 2022; 10(10):2066. https://doi.org/10.3390/microorganisms10102066
Chicago/Turabian StyleLyu, Juanjuan, Fangfang Kou, Xiangyu Men, Yinhui Liu, Li Tang, and Shu Wen. 2022. "The Changes in Bacterial Microbiome Associated with Immune Disorder in Allergic Respiratory Disease" Microorganisms 10, no. 10: 2066. https://doi.org/10.3390/microorganisms10102066