
Deletion of the Viral Thymidine Kinase in a Meq-Deleted Recombinant Marek’s Disease Virus Reduces Lymphoid Atrophy but Is Less Protective

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Animals
2.2. Cells and Viruses
Ablation of the Viral Thymidine Kinase
2.3. Assessment of Recombinant MDVs In Vitro
2.4. Quantification of Viremia Produced by Recombinant MDVs
2.5. Assessment of Lymphoid Atrophy Caused by Recombinant MDVs
2.6. Assessment of Protection Provided by rMDVs
2.7. Significance Level and Statistical Analysis of Data
3. Results
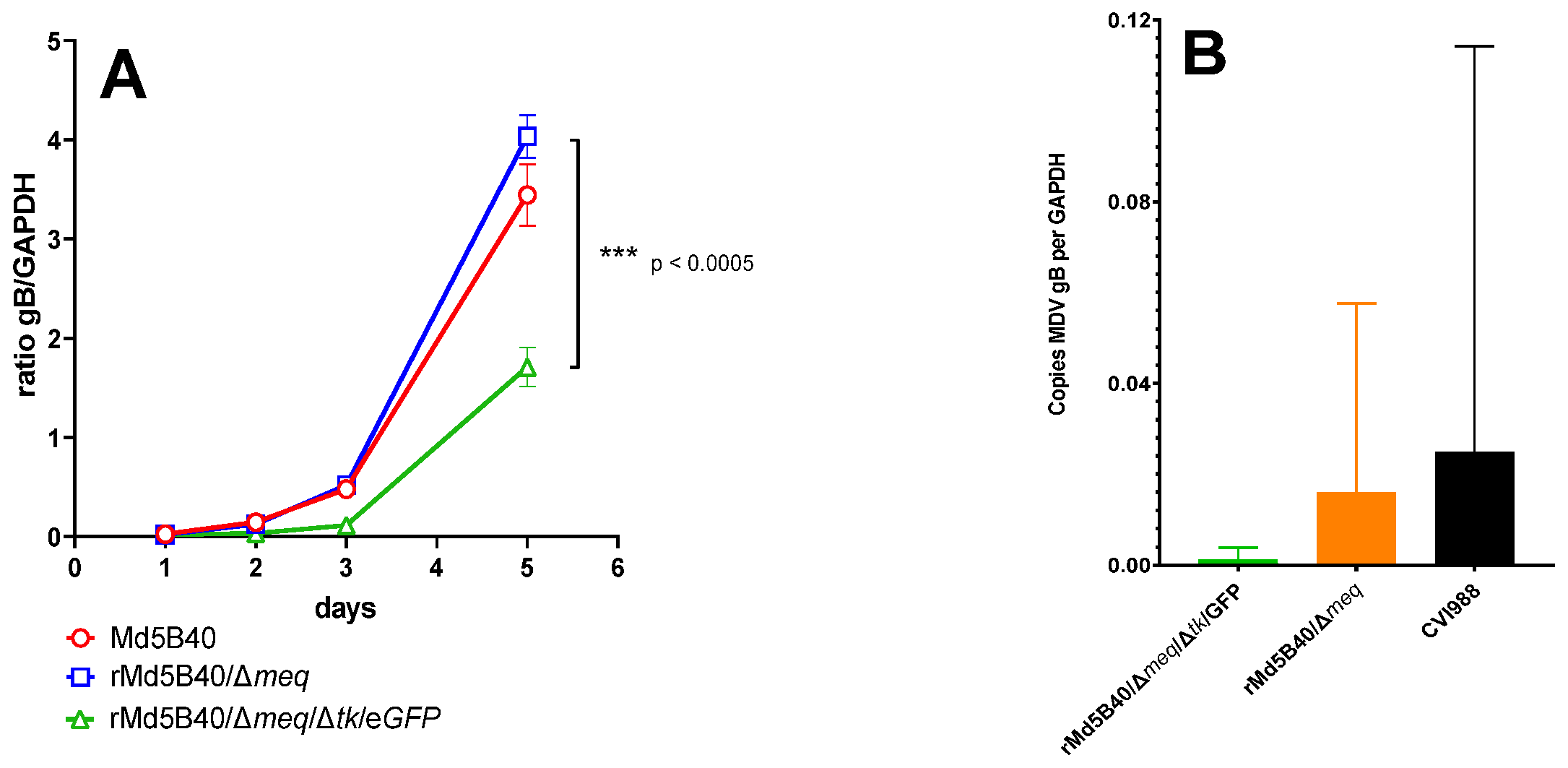
3.1. Ablation of the Viral tk from the rMd5B40/Δmeq Virus Attenuates Replication In Vitro
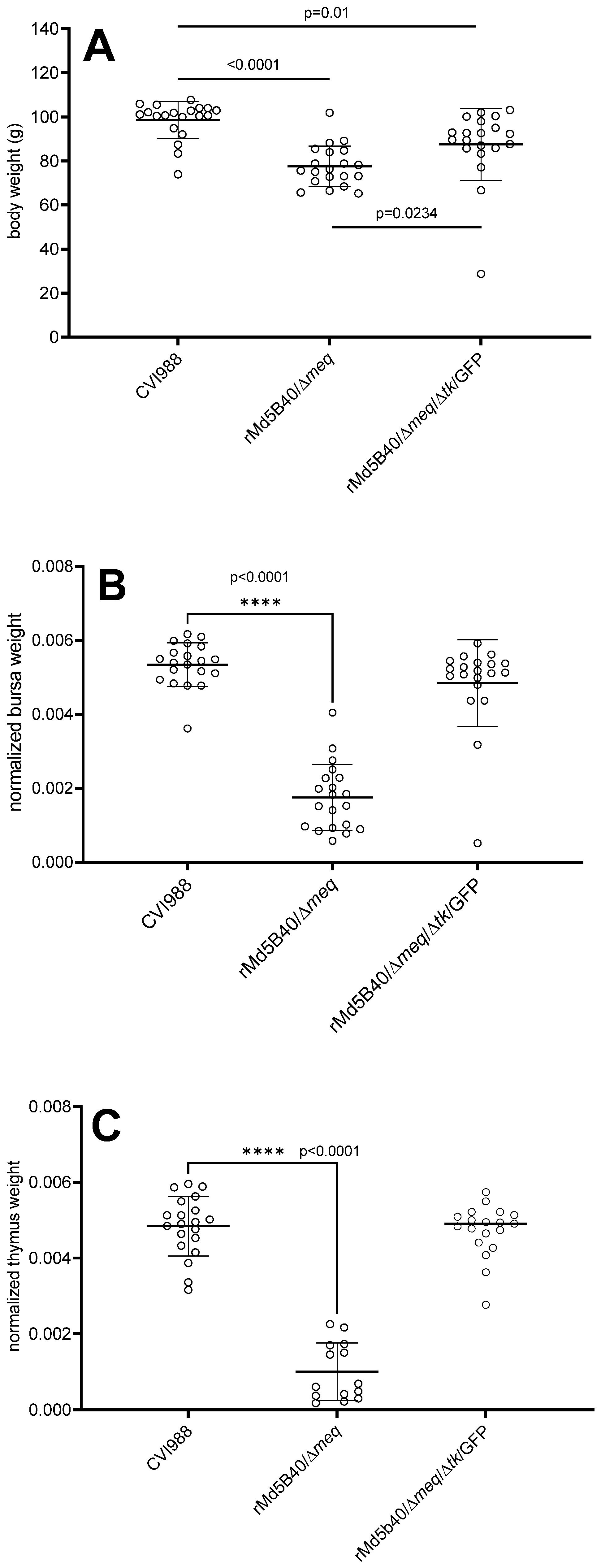
3.2. Attenuation of the rMd5B40/Δmeq Virus by Ablation of the Viral tk Spared Chickens from Lymphoid Atrophy
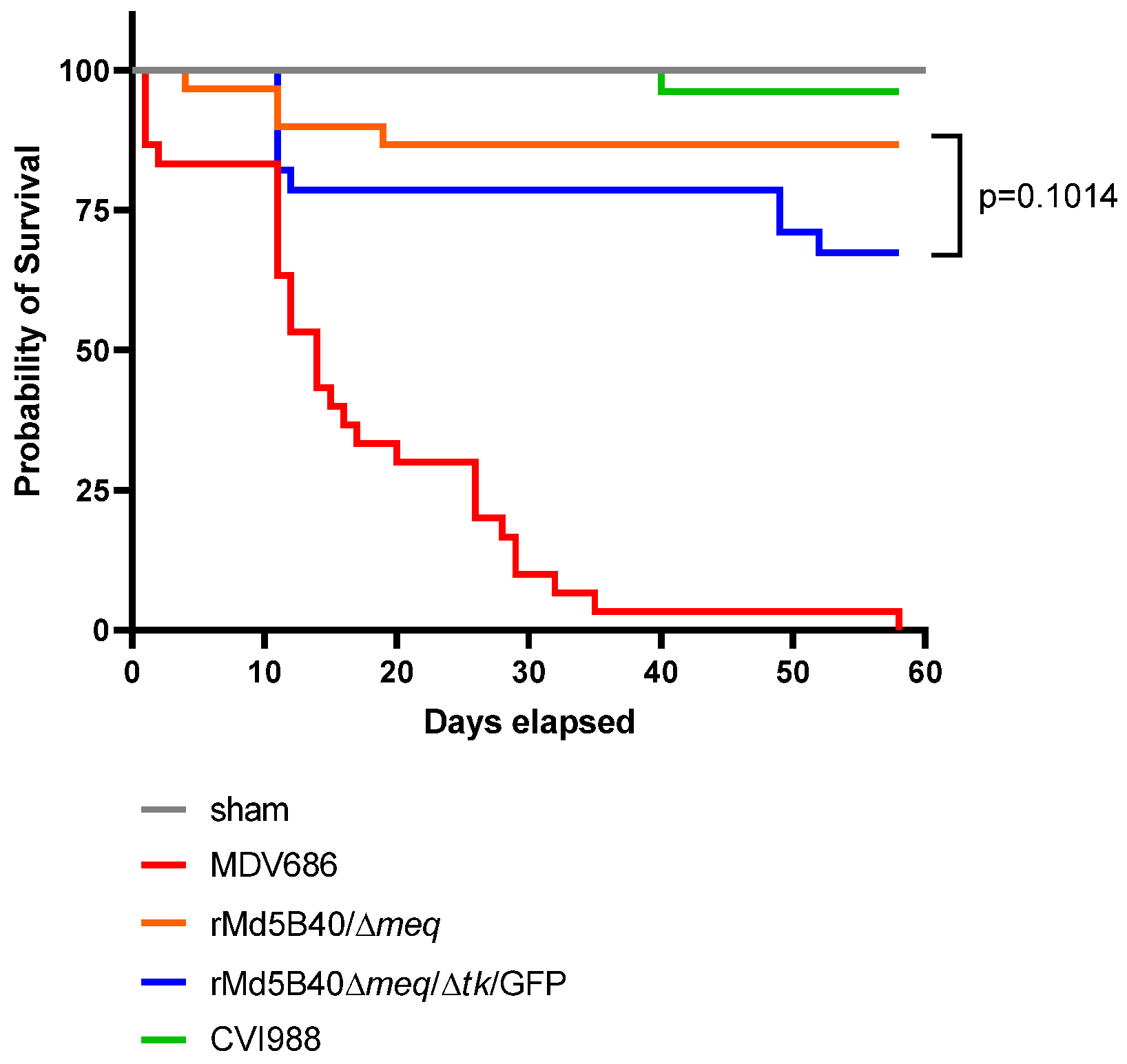
3.3. The rMd5B40/Δmeq/Δtk/GFP Viruse Displayed Reduced Protection against MD, against the Development of Visceral Tumors and against Mortality When Challenged with a vv+ MD
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s Disease Virus: From Miasma to Model. Nat. Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef]
- Bertzbach, L.D.; Conradie, A.M.; You, Y.; Kaufer, B.B. Latest Insights into Marek’s Disease Virus Pathogenesis and Tumorigenesis. Cancers 2020, 12, 647. [Google Scholar] [CrossRef] [Green Version]
- Calnek, B.W. Pathogenesis of Marek’s Disease Virus Infection. In Marek’s Disease; Hirai, K., Ed.; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2001; Volume 255. [Google Scholar] [CrossRef]
- Jarosinski, K.W. Marek’s Disease Virus Late Protein Expression in Feather Follicle Epithelial Cells as Early as 8 Days Postinfection. Avian Dis. 2012, 56, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Bertzbach, L.D.; Laparidou, M.; Härtle, S.; Etches, R.J.; Kaspers, B.; Schusser, B.; Kaufer, B.B. Unraveling the Role of B Cells in the Pathogenesis of an Oncogenic Avian Herpesvirus. Proc. Natl. Acad. Sci. USA 2018, 115, 11603–11607. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.T.; Selvaraj, R.K.; Kamil, J.P.; Osterrieder, N.; Kaufer, B.B. Marek’s Disease Viral Interleukin-8 Promotes Lymphoma Formation through Targeted Recruitment of B Cells and CD4+ CD25+ T Cells. J. Virol. 2012, 86, 8536–8545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boodhoo, N.; Gurung, A.; Sharif, S.; Behboudi, S. Marek’s Disease in Chickens: A Review with Focus on Immunology. Veter. Res. 2016, 47, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, C.M.; Cheng, H.H.; Delany, M.E. Temporal Kinetics of Marek’s Disease Herpesvirus: Integration Occurs Early after Infection in Both B and T Cells. Cytogenet. Genome Res. 2014, 144, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Parcells, M.; Arumugaswami, V.; Prigge, J.; Pandya, K.; Dienglewicz, R. Marek’s Disease Virus Reactivation from Latency: Changes in Gene Expression at the Origin of Replication. Poult. Sci. 2003, 82, 893–898. [Google Scholar] [CrossRef]
- Rispens, B.H.; van Vloten, H.; Mastenbroek, N.; Maas, H.J.; Schat, K.A. Control of Marek’s disease in the Netherlands. I. Isolation of an avirulent Marek’s disease virus (strain CVI 988) and its use in laboratory vaccination trials. Avian Dis. 1972, 16, 108–125. [Google Scholar] [CrossRef]
- Read, A.F.; Baigent, S.J.; Powers, C.; Kgosana, L.B.; Blackwell, L.; Smith, L.P.; Kennedy, D.A.; Walkden-Brown, S.W.; Nair, V.K. Imperfect Vaccination Can Enhance the Transmission of Highly Virulent Pathogens. PLoS Biol. 2015, 13, e1002198. [Google Scholar] [CrossRef]
- Bailey, R.I.; Cheng, H.H.; Chase-Topping, M.; Mays, J.K.; Anacleto, O.; Dunn, J.R.; Doeschl-Wilson, A. Pathogen transmission from vaccinated hosts can cause dose-dependent reduction in virulence. PLoS Biol. 2020, 18, e3000619. [Google Scholar] [CrossRef] [Green Version]
- Witter, R.L. Increased Virulence of Marek’s Disease Virus Field Isolates. Avian Dis. 1997, 41, 149. [Google Scholar] [CrossRef] [PubMed]
- Trimpert, J.; Groenke, N.; Jenckel, M.; He, S.; Kunec, D.; Szpara, M.L.; Spatz, S.J.; Osterrieder, N.; McMahon, D.P. A Phylogenomic Analysis of Marek’s Disease Virus Reveals Independent Paths to Virulence in Eurasia and North America. Evol. Appl. 2017, 10, 1091–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witter, R.L. The Changing Landscape of Marek’s Disease. Avian Pathol. 1998, 27, S46–S53. [Google Scholar] [CrossRef]
- Jones, D.; Lee, L.; Liu, J.L.; Kung, H.J.; Tillotson, J.K. Marek Disease Virus Encodes a Basic-Leucine Zipper Gene Resembling the Fos/Jun Oncogenes That Is Highly Expressed in Lymphoblastoid Tumors. Proc. Natl. Acad. Sci. USA 1992, 89, 4042–4046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Rock, D.L.; Kutish, G.F. The Genome of a Very Virulent Marek’s Disease Virus. J. Virol. 2000, 74, 7980–7988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchodolski, P.F.; Izumiya, Y.; Lupiani, B.; Ajithdoss, D.K.; Gilad, O.; Lee, L.F.; Kung, H.-J.; Reddy, S.M. Homodimerization of Marek’s Disease Virus-Encoded Meq Protein Is Not Sufficient for Transformation of Lymphocytes in Chickens. J. Virol. 2008, 83, 859–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, A.M.; Izumiya, Y.; Brunovskis, P.; Xia, L.; Parcells, M.S.; Reddy, S.M.; Lee, L.; Chen, H.-W.; Kung, H.-J. Characterization of the Chromosomal Binding Sites and Dimerization Partners of the Viral Oncoprotein Meq in Marek’s Disease Virus-Transformed T Cells. J. Virol. 2003, 77, 12841–12851. [Google Scholar] [CrossRef] [Green Version]
- Suchodolski, P.F.; Izumiya, Y.; Lupiani, B.; Ajithdoss, D.K.; Lee, L.F.; Kung, H.-J.; Reddy, S.M. Both Homo and Heterodimers of Marek’s Disease Virus Encoded Meq Protein Contribute to Transformation of Lymphocytes in Chickens. Virology 2010, 399, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Levy, A.M.; Gilad, O.; Xia, L.; Izumiya, Y.; Choi, J.; Tsalenko, A.; Yakhini, Z.; Witter, R.; Lee, L.; Cardona, C.J.; et al. Marek’s Disease Virus Meq Transforms Chicken Cells via the v-Jun Transcriptional Cascade: A Converging Transforming Pathway for Avian Oncoviruses. Proc. Natl. Acad. Sci. USA 2005, 102, 14831–14836. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Li, X.; Shen, Y.; Qiu, Y.; Shi, Z.; Shao, D.; Jin, Y.; Chen, H.; Ding, C.; Li, L.; et al. The Meq Oncoprotein of Marek’s Disease Virus Interacts with P53 and Inhibits Its Transcriptional and Apoptotic Activities. Virol. J. 2010, 7, 348. [Google Scholar] [CrossRef] [Green Version]
- Lupiani, B.; Lee, L.F.; Cui, X.; Gimeno, I.; Anderson, A.; Morgan, R.W.; Silva, R.F.; Witter, R.L.; Kung, H.-J.; Reddy, S.M. Marek’s Disease Virus-Encoded Meq Gene Is Involved in Transformation of Lymphocytes but Is Dispensable for Replication. Proc. Natl. Acad. Sci. USA 2004, 101, 11815–11820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Fu, S.; Deng, M.; Xie, Q.; Xu, H.; Liu, Z.; Hu, C.; Chen, H.; Guo, A. Attenuation of Bovine Herpesvirus Type 1 by Deletion of Its Glycoprotein G and Tk Genes and Protection against Virulent Viral Challenge. Vaccine 2011, 29, 8943–8950. [Google Scholar] [CrossRef]
- Sun, P.; Cui, N.; Liu, L.; Su, S.; Cheng, Z.; Chen, R.; Li, Y.; Cui, Z. Attenuation of a Recombinant Marek’s Disease Virus Lacking the Meq Oncogene and Evaluation on Its Immune Efficacy against Marek’s Disease Virus. Poult. Sci. 2020, 99, 1939–1945. [Google Scholar] [CrossRef]
- Niikura, M.; Kim, T.; Silva, R.F.; Dodgson, J.; Cheng, H.H. Virulent Marek’s Disease Virus Generated from Infectious Bacterial Artificial Chromosome Clones with Complete DNA Sequence and the Implication of Viral Genetic Homogeneity in Pathogenesis. J. Gen. Virol. 2010, 92, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Noll, J.C.G.; Joshi, L.R.; do Nascimento, G.M.; Fernandes, M.H.V.; Sharma, B.; Flores, E.F.; Diel, D.G. Deletion of the Thymidine Kinase Gene Attenuates Caprine Alphaherpesvirus 1 in Goats. Veter. Microbiol. 2019, 237, 108370. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wu, L.; Wang, M.; Cheng, A.; Yang, Q.; Wu, Y.; Jia, R.; Zhu, D.; Zhao, X.; Chen, S.; et al. Alpha-Herpesvirus Thymidine Kinase Genes Mediate Viral Virulence and Are Potential Therapeutic Targets. Front. Microbiol. 2019, 10, 941. [Google Scholar] [CrossRef] [Green Version]
- Kokoris, M.S.; Black, M.E. Characterization of Herpes Simplex Virus Type 1 Thymidine Kinase Mutants Engineered for Improved Ganciclovir or Acyclovir Activity. Protein Sci. 2002, 11, 2267–2272. [Google Scholar] [CrossRef] [Green Version]
- Sanford, B.; Holinka, L.G.; O’Donnell, V.; Krug, P.W.; Carlson, J.; Alfano, M.; Carrillo, C.; Wu, P.; Lowe, A.; Risatti, G.R.; et al. Deletion of the Thymidine Kinase Gene Induces Complete Attenuation of the Georgia Isolate of African Swine Fever Virus. Virus Res. 2016, 213, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Fan, J.; Ding, Y.; Zhang, J.; Zhou, B.; Zhang, Y.; Huang, B. Oncolytic Efficacy of Thymidine Kinase-Deleted Vaccinia Virus Strain Guang9. Oncotarget 2014, 5, 40533–40543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, R.F.; Dunn, J.R.; Cheng, H.H.; Niikura, M. A MEQ-Deleted Marek’s Disease Virus Cloned as a Bacterial Artificial Chromosome Is a Highly Efficacious Vaccine. Avian Dis. 2010, 54, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Karstentischer, B.; von Einem, J.; Kaufer, B.; Osterrieder, N. Two-Step Red-Mediated Recombination for Versatile High-Efficiency Markerless DNA Manipulation in Escherichia coli. Biotechniques 2006, 40, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Cadore, G.C.; Weiss, M.; Anziliero, D.; Brum, M.C.S.; Weiblen, R.; Flores, E.F. A Thymidine Kinase-Deleted Bovine Herpesvirus 5 Establishes Latent Infection but Reactivates Poorly in a Sheep Model. Pesquisa Veterinária Brasileira 2013, 33, 331–338. [Google Scholar] [CrossRef]
- Liao, Y.; Reddy, S.M.; Khan, O.A.; Sun, A.; Lupiani, B. A Novel Effective and Safe Vaccine for Prevention of Marek’s Disease Caused by Infection with a Very Virulent Plus (Vv+) Marek’s Disease Virus. Vaccines 2021, 9, 159. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Vaccine Strain | Challenge Strain | MD+/Total |
|---|---|---|
| none | MDV686 | 25/30 |
| CVI988/Rispens | MDV686 | 01/30 |
| rMd5B40/Δmeq | MDV686 | 03/30 |
| rMd5B40/Δmeq/tk/GFP | MDV686 | 23/32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conrad, S.J.; Oluwayinka, E.B.; Heidari, M.; Mays, J.K.; Dunn, J.R. Deletion of the Viral Thymidine Kinase in a Meq-Deleted Recombinant Marek’s Disease Virus Reduces Lymphoid Atrophy but Is Less Protective. Microorganisms 2022, 10, 7. https://doi.org/10.3390/microorganisms10010007
Conrad SJ, Oluwayinka EB, Heidari M, Mays JK, Dunn JR. Deletion of the Viral Thymidine Kinase in a Meq-Deleted Recombinant Marek’s Disease Virus Reduces Lymphoid Atrophy but Is Less Protective. Microorganisms. 2022; 10(1):7. https://doi.org/10.3390/microorganisms10010007
Chicago/Turabian StyleConrad, Steven J., Eniope B. Oluwayinka, Mohammad Heidari, Jody K. Mays, and John R. Dunn. 2022. "Deletion of the Viral Thymidine Kinase in a Meq-Deleted Recombinant Marek’s Disease Virus Reduces Lymphoid Atrophy but Is Less Protective" Microorganisms 10, no. 1: 7. https://doi.org/10.3390/microorganisms10010007