Upscaling the Surveillance of Tick-Borne Pathogens in the French Caribbean Islands

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
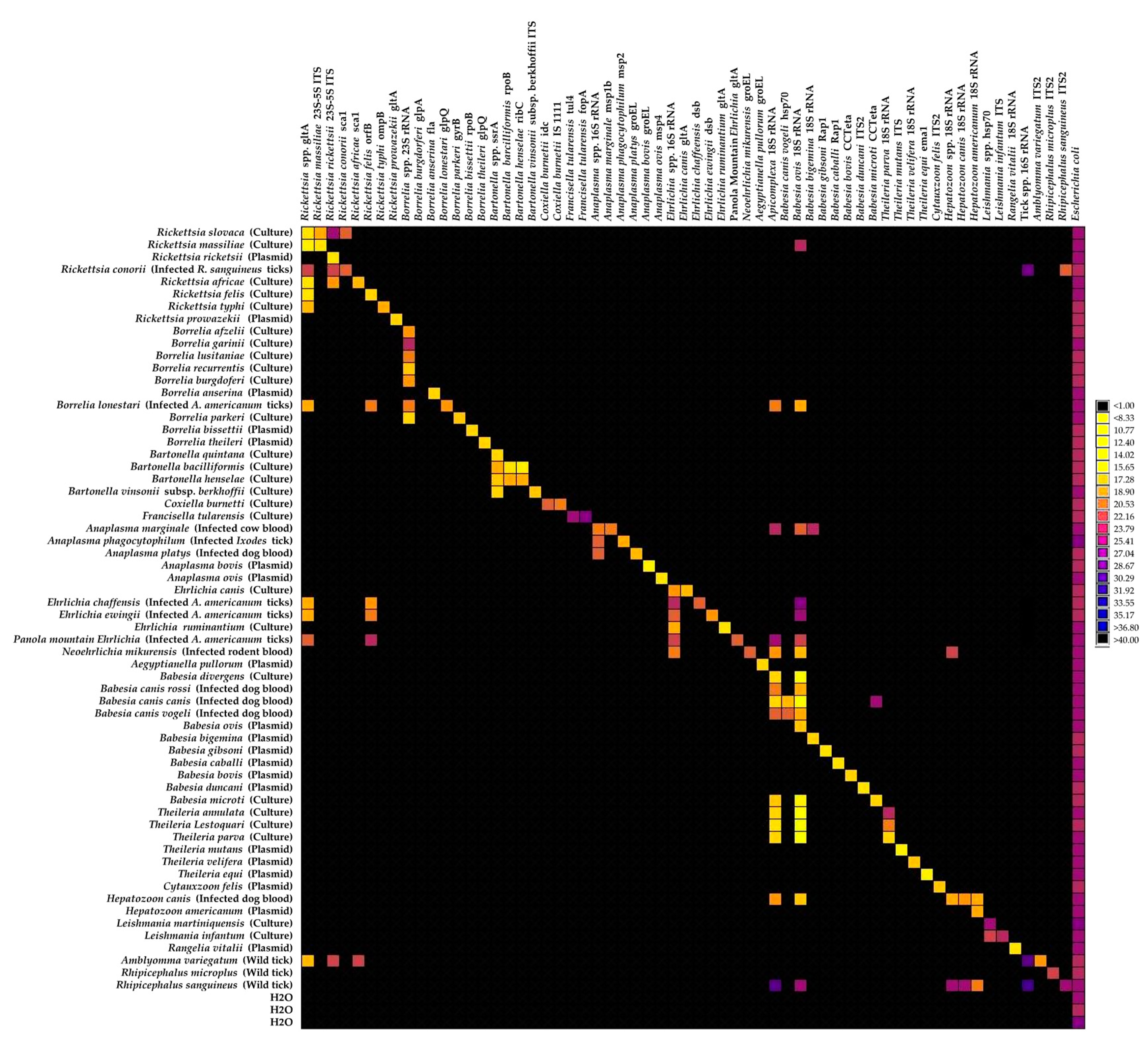
2.1. Implementation of the High-Throughput Microfluidic Real-Time PCR System for Tick-Borne Pathogen Screening
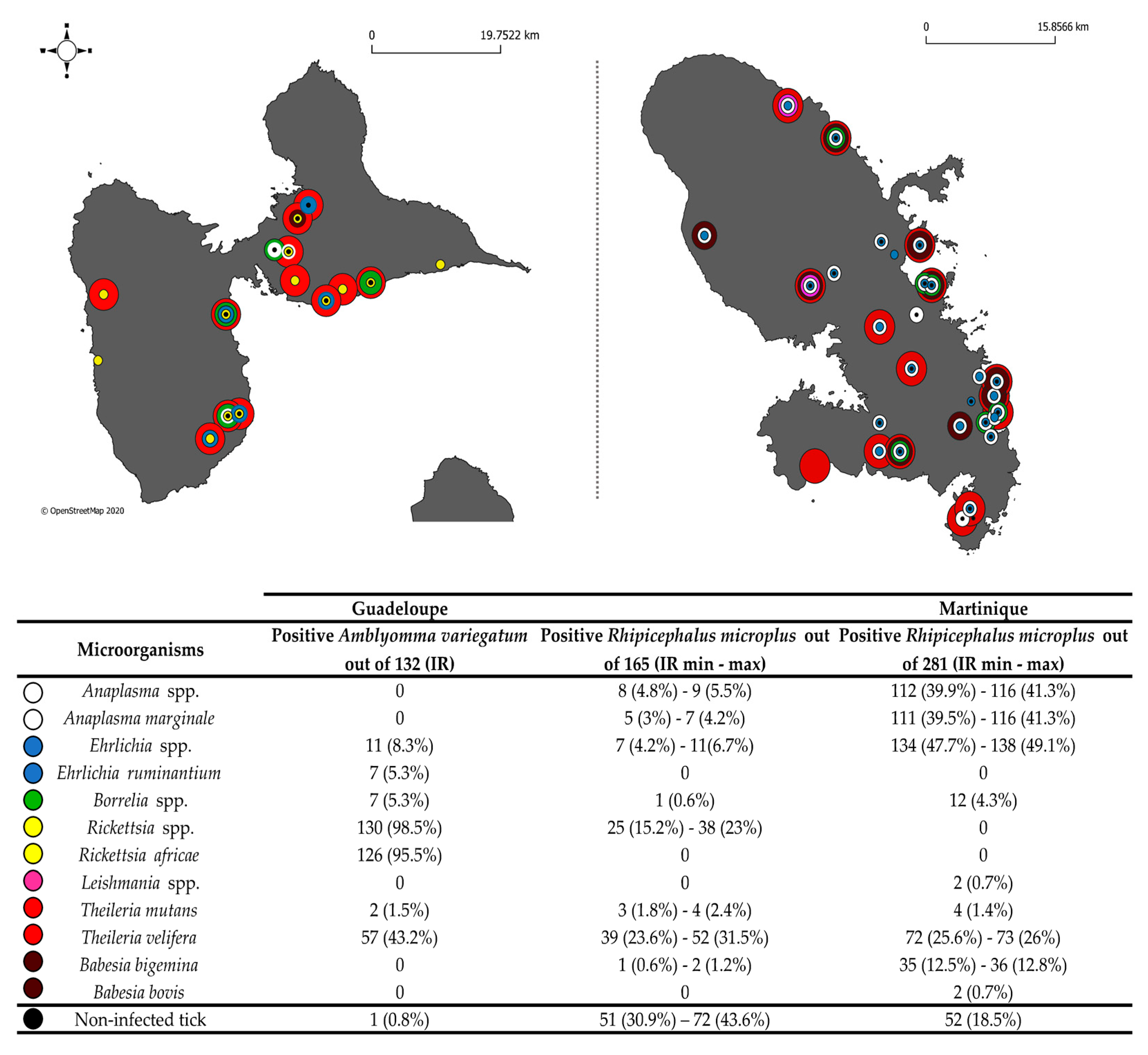
2.2. Large-Scale TBP Detection Survey in Ticks from Guadeloupe and Martinique
2.2.1. Detection of Known TBPs in Caribbean Ticks
2.2.2. Detection of Unexpected Microorganisms in Caribbean Ticks
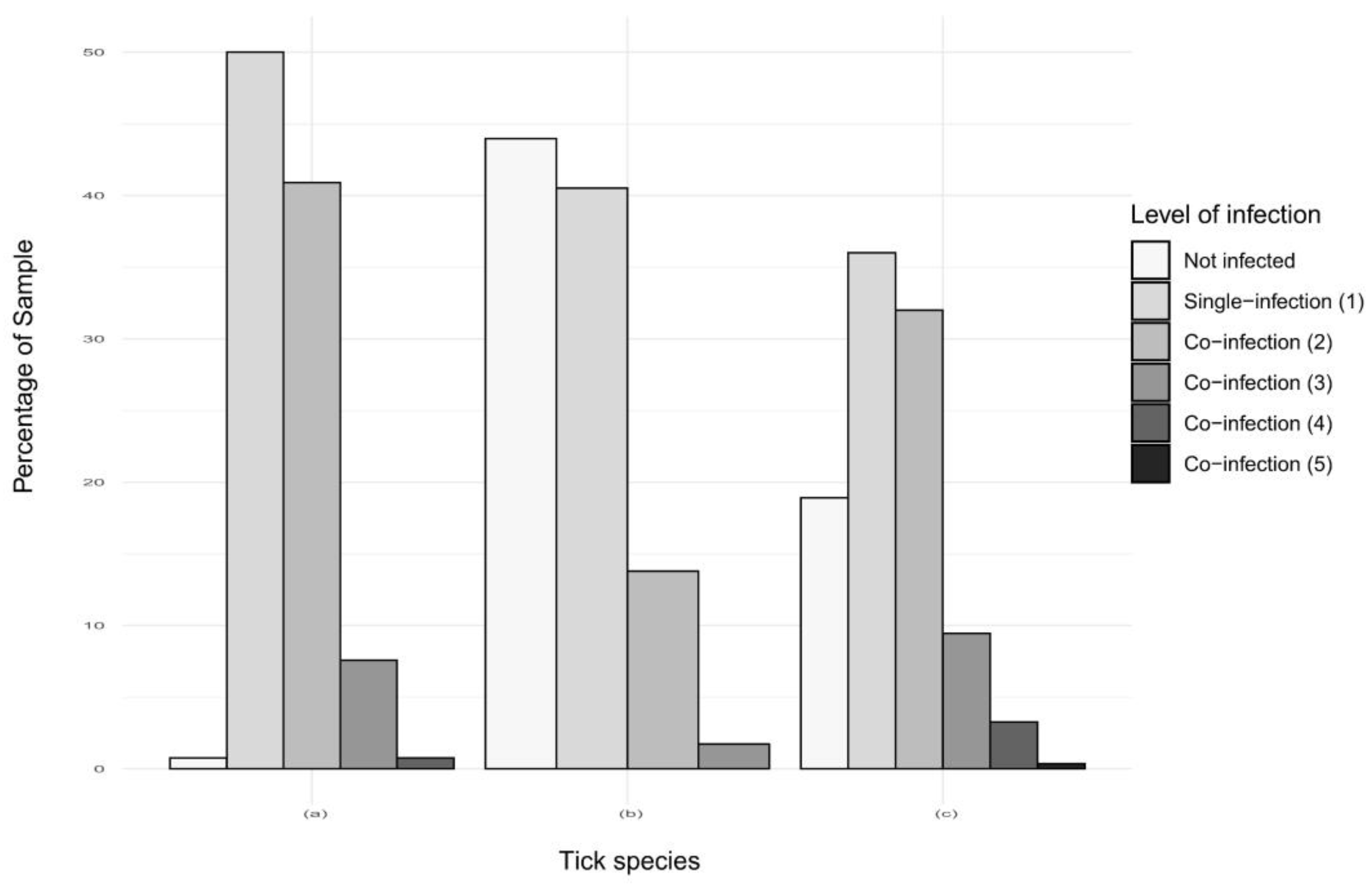
2.2.3. Co-Infections in Ticks in Guadeloupe and Martinique
3. Discussion
4. Materials and Methods
4.1. Ticks Collected in Guadeloupe and Martinique
4.2. DNA Extraction of Ticks Collected in Guadeloupe and Martinique
4.3. Assay Design
4.4. Pre-Amplification of DNA Samples
4.5. High-Throughput Microfluidic Real-Time PCR
4.6. Infection Rates for Ticks from the French West Indies
4.7. PCRs and Sequencing for the Confirmation of Results
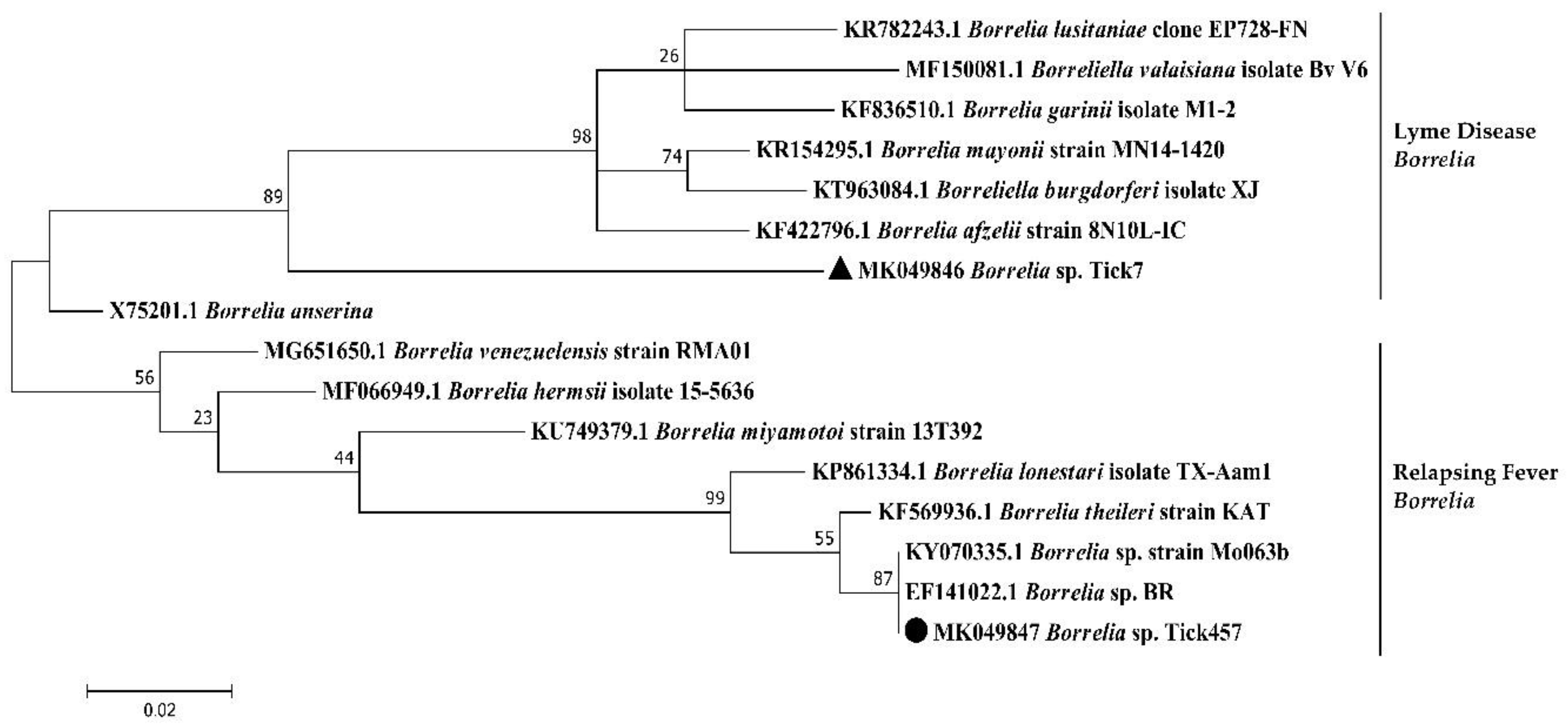
4.8. Phylogenetic Sequence Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Design | Target Detection | Specificity | Outgroup Control Samples |
|---|---|---|---|
| Rickettsia spp. gltA | YES | CI | Borrelia lonestari (Infected A. americanum ticks) |
| YES | CI | Ehrlichia chaffensis (Infected A. americanum ticks) | |
| YES | CI | Ehrlichia ewingii (Infected A. americanum ticks) | |
| YES | CI | Panola Mountain Ehrlichia (Infected A. americanum ticks) | |
| YES | CI | Amblyomma variegatum (Wild tick) | |
| Rickettsia massiliae 23S-5S ITS | YES | CR | Rickettsia slovaca (Culture) |
| Rickettsia rickettsii 23S-5S ITS | YES | CR 1 | Rickettsia slovaca (Culture) |
| YES | Rickettsia conorii (Infected R. sanguineus s.l. ticks) | ||
| YES | Rickettsia africae (Culture) | ||
| YES | Amblyomma variegatum (Wild tick) | ||
| Rickettsia conorii sca1 | YES | CR | Rickettsia slovaca (Culture) |
| Rickettsia africae sca1 | YES | CI | Amblyomma variegatum (Wild tick) |
| Rickettsia felis orfB | YES | CI | Borrelia lonestari (Infected A. americanum ticks) |
| YES | CI | Ehrlichia chaffensis (Infected A. americanum ticks) | |
| YES | CI | Ehrlichia ewingii (Infected A. americanum ticks) | |
| YES | CI | Panola Mountain Ehrlichia (Infected A. americanum ticks) | |
| Rickettsia typhi ompB | YES | YES | |
| Rickettsia prowazekii gltA | YES | YES | |
| Borrelia spp. 23S rRNA | YES | YES | |
| Borrelia burgdorferi sensu stricto glpA | NO 1 | ||
| Borrelia anserina fla | YES | YES | |
| Borrelia lonestari glpQ | YES | YES | |
| Borrelia parkeri gyrB | YES | YES | |
| Borrelia bissettii rpoB | YES | YES | |
| Borrelia theileri glpQ | YES | YES | |
| Bartonella spp. ssrA | YES | YES | |
| Bartonella bacilliformis rpoB | YES | CR | Bartonella henselae (Culture) |
| Bartonella henselae ribC | YES | CR | Bartonella bacilliformis (Culture) |
| Bartonella vinsonii subsp. berkhoffii ITS | YES | YES | |
| Coxiella burnetii icd | YES | YES | |
| Coxiella burnetii IS 1111 | YES | YES | |
| Francisella tularensis tul4 | YES | YES | |
| Francisella tularensis fopA | YES | YES | |
| Anaplasma spp. 16S rRNA | YES | YES | |
| Anaplasma marginale msp1b | YES | YES | |
| Anaplasma phagocytophilum msp2 | YES | YES | |
| Anaplasma platys groEL | YES | YES | |
| Anaplasma bovis groEL | YES | YES | |
| Anaplasma ovis msp4 | YES | YES | |
| Ehrlichia spp. 16S rRNA | YES | YES | |
| Ehrlichia canis gltA | YES | YES | |
| Ehrlichia chaffeensis dsb | YES | YES | |
| Ehrlichia ewingii dsb | YES | YES | |
| Ehrlichia ruminantium gltA | YES | YES | |
| Panola Mountain Ehrlichia gltA | YES | YES | |
| Neoehrlichia mikurensis groEL | YES | YES | |
| Aegyptianella pullorum groEL | YES | YES | |
| Apicomplexa 18S rRNA | YES | CI | Borrelia lonestari (Infected A. americanum ticks) |
| YES | CI | Anaplasma marginale (Infected cow blood) | |
| YES | CI | Panola Mountain Ehrlichia (Infected A. americanum ticks) | |
| YES | CI | Neoehrlichia mikurensis (Infected rodent blood) | |
| YES | CI | Rhipicephalus sanguineus s.l. (Wild tick) | |
| Babesia canis vogeli hsp70 | YES | CR | Babesia canis canis (Infected dog blood) |
| Babesia ovis 18S rRNA | YES | CR 1 | Rickettsia massiliae (Culture) |
| YES | Borrelia lonestari (Infected A. americanum ticks) | ||
| YES | Anaplasma marginale (Infected cow blood) | ||
| YES | Ehrlichia chaffensis (Infected A. americanum ticks) | ||
| YES | Ehrlichia ewingii (Infected A. americanum ticks) | ||
| YES | Panola Mountain Ehrlichia (Infected A. americanum ticks) | ||
| YES | Neoehrlichia mikurensis (Infected rodent blood) | ||
| YES | Babesia divergens (Culture) | ||
| YES | Babesia canis rossi (Infected dog blood) | ||
| YES | Babesia canis canis (Infected dog blood) | ||
| YES | Babesia canis vogeli (Infected dog blood) | ||
| YES | Babesia microti (Culture) | ||
| YES | Theileria annulata (Culture) | ||
| YES | Theileria lestoquardi (Culture) | ||
| YES | Theileria parva (Culture) | ||
| YES | Hepatozoon canis (Infected dog blood) | ||
| YES | Rhipicephalus sanguineus s.l. (Wild tick) | ||
| Babesia bigemina 18S rRNA | YES | CI | Anaplasma marginale (Infected cow blood) |
| Babesia gibsoni Rap1 | YES | YES | |
| Babesia caballi Rap1 | YES | YES | |
| Babesia bovis CCTeta | YES | YES | |
| Babesia duncani ITS2 | YES | YES | |
| Babesia microti CCTeta | YES | CR | Babesia canis canis (Infected dog blood) |
| Theileria parva 18S rRNA | YES | CR | Theileria annulata (Culture) |
| YES | CR | Theileria lestoquardi (Culture) | |
| Theileria mutans ITS | YES | YES | |
| Theileria velifera 18S rRNA | YES | YES | |
| Theileria equi ema1 | YES | YES | |
| Cytauxzoon felis ITS2 | YES | YES | |
| Hepatozoon spp 18S rRNA | YES | CI | Neoehrlichia mikurensis (Infected rodent blood) |
| YES | CI | Rhipicephalus sanguineus s.l. (Wild tick) | |
| Hepatozoon canis 18S rRNA | YES | CI | Rhipicephalus sanguineus s.l. (Wild tick) |
| Hepatozoon americanum 18S rRNA | YES | CR | Hepatozoon canis (Infected dog blood) |
| YES | CI | Rhipicephalus sanguineus s.l. (Wild tick) | |
| Leishmania spp hsp70 | YES | YES | |
| Leishmania infantum ITS | YES | YES | |
| Rangelia vitalii 18S rRNA | YES | YES | |
| Tick spp. 16S rRNA | YES | YES 2 | |
| Amblyomma variegatum ITS2 | YES | YES | |
| Rhipicephalus microplus ITS2 | YES | YES | |
| Rhipicephalus sanguineus s.l. ITS2 | YES | YES |
| Tested Control Sample | Tested for | AN | Length (bp) | Closest Homology | % I | % C | AN |
|---|---|---|---|---|---|---|---|
| Panola Mountain Ehrlichia (Infected A. americanum ticks) | Rickettsia spp. | MK049843 | 382 | Rickettsia spp. | 99 | 100 | MF511253.1 |
| Neoehrlichia mikurensis (Infected rodent blood) | Hepatozoon spp. | MK071735 | 169 | Hepatozoon spp. | 99 | 100 | AB771515.1 |
| Anaplasma marginale | Apicomplexa | MK071737 | 104 | Babesia spp., Theileria spp. (including B. bigemina) | 100 | 99 | MG604302.1 |
| (Infected cow blood) | |||||||
| Panola Mountain Ehrlichia (Infected A. americanum ticks) | Apicomplexa | MK071736 | 102 | Theileria spp. (including T. cervi) | 98 | 100 | MH085203.1 |
| Borrelia lonestari | Apicomplexa | ||||||
| (Infected A. americanum ticks) |
Appendix B
| A. variegatum Sample (Percentage) | R. microplus Sample from Guadeloupe (Percentage) | R. microplus Sample from Martinique (Percentage) | ||
|---|---|---|---|---|
| Total of sample | 132 | 116 | 275 | |
| Total of non-infected sample | 1 (0.8) | 51 (44) | 52 (19) | |
| Total of infected sample | 131 (99.2) | 65 (56) | 223 (81) | |
| Single infections | Total | 69 (52.3) | 47 (40.5) | 99 (36) |
| R. africae 1 | 68 (51.5) | 15 (12.9) | 0 | |
| Anaplasma spp. 2 | 1 (0.9) | 10 (3.6) | ||
| A. marginale | 0 | 2 (1.7) | 22 (8) | |
| Ehrlichia spp. 3 | 0 | 3 (2.6) | 43 (15.6) | |
| Borrelia spp. | 0 | 0 | 3 (1.1) | |
| B. bigemina | 0 | 1 (0.9) | 7 (2.5) | |
| T. velifera | 1 (0.8) | 25 (21.6) | 14 (5.1) | |
| Co-infections (2) | Total | 53 (40.2) | 16 (13.8) | 88 (32) |
| R. africae 1/T. velifera | 47 (35.6) | 6 (5.2) | 0 | |
| R. africae 1/Borrelia spp. | 3 (2.3) | 0 | 0 | |
| R. africae 1/E. ruminantium | 3 (2.3) | 0 | 0 | |
| Anaplasma spp. 2/Borrelia spp. | 0 | 1 (0.9) | 0 | |
| Anaplasma spp. 2/T. velifera | 0 | 2 (1.7) | 3 (1.1) | |
| Anaplasma spp. 2/Ehrlichia spp. 3 | 0 | 0 | 6 (2.2) | |
| A. marginale/T. velifera | 0 | 2 (1.7) | 10 (3.6) | |
| A. marginale/Borrelia spp. | 0 | 0 | 3 (1.1) | |
| A. marginale/Ehrlichia spp. 3 | 0 | 0 | 39 (14.29) | |
| A. marginale/B. bigemina | 0 | 0 | 4 (1.5) | |
| Ehrlichia spp. 3/R. africae 1 | 0 | 2 (1.7) | 0 | |
| Ehrlichia spp. 3/T. velifera | 0 | 1 (0.9) | 8 (2.9) | |
| Ehrlichia spp. 3/Borrelia spp. | 0 | 0 | 3 (1.1) | |
| Ehrlichia spp. 3/Leishmania spp. | 0 | 0 | 1 (0.4) | |
| T. velifera/B. bigemina | 0 | 0 | 10 (3.6) | |
| T. velifera/T. mutans | 0 | 2 (1.7) | 1 (0.4) | |
| Co-infections (3) | Total | 8 (6.1) | 2 (1.7) | 26 (9.5) |
| R.africae 1/E. ruminantium/T. velifera | 3 (2.3) | 0 | 0 | |
| R. africae 1/Borrelia spp./T. velifera | 3 (2.3) | 0 | 0 | |
| R. africae 1/T. velifera/T. mutans | 2 (1.5) | 1 (0.6) | 0 | |
| R. africae 1/A. marginale/Ehrlichia spp. 3 | 0 | 1 (0.6) | 0 | |
| A. marginale/Ehrlichia spp. 3/B. bigemina | 0 | 0 | 8 (2.9) | |
| A. marginale/Ehrlichia spp. 3/B. bovis | 0 | 0 | 1 (0.4) | |
| A. marginale/Ehrlichia spp. 3/T. velifera | 0 | 0 | 15 (5.5) | |
| Anaplasma spp. 2/Ehrlichia spp. 3/B. bigemina | 0 | 0 | 1 (0.4) | |
| Anaplasma spp. 2/Ehrlichia spp. 3/T. velifera | 0 | 0 | 1 (0.4) | |
| Co-infections (4) | Total | 1 (0.8) | 0 | 9 (3.3) |
| R. africae 1/T. velifera/Borrelia spp./ E. ruminantium | 1 (0.8) | 0 | 0 | |
| A. marginale/Borrelia spp./T. velifera/ T. mutans | 0 | 0 | 1 (0.4) | |
| A. marginale/Ehrlichia spp. 3/T. velifera/ B. bigemina | 0 | 0 | 5 (1.8) | |
| A. marginale/Ehrlichia spp. 3 /T. velifera/B. bovis | 0 | 0 | 1 (0.4) | |
| A. marginale/Leishmania spp./T. velifera/ T. mutans | 0 | 0 | 1 (0.4) | |
| Anaplasma spp. 2/Ehrlichia spp. 3/Borrelia spp./ T. velifera | 0 | 0 | 1 (0.4) | |
| Co-infections (5) | Total | 0 | 0 | 1 (0.4) |
| A. marginale/Ehrlichia spp. 3/Borrelia spp./ T. velifera/T. mutans | 0 | 0 | 1 (0.4) | |
| E. ruminantium | Borrelia spp. | R. africae | T. mutans | T. velifera | |
|---|---|---|---|---|---|
| E. ruminantium | 7 | 1 | 7 | 0 | 4 |
| Borrelia spp. | 7 | 7 | 0 | 4 | |
| R. africae | 130 | 2 | 56 | ||
| T. mutans | 2 | 2 | |||
| T. velifera | 57 |
| A. marginale | Borrelia spp. | R. africae | T. mutans | T. velifera | B. bigemina | Anaplasma spp.1 | Ehrlichia spp.2 | |
|---|---|---|---|---|---|---|---|---|
| A. marginale | 5 | 0 | 1 | 0 | 2 | 0 | 0 | 1 |
| Borrelia spp. | 1 | 0 | 0 | 0 | 0 | 1* | 0 | |
| R. africae | 25 | 1 | 7 | 0 | 0 | 3 | ||
| T. mutans | 3 | 3* | 0 | 0 | 0 | |||
| T. velifera | 39 | 0 | 2 | 1 | ||||
| B. bigemina | 1 | 0 | 0 | |||||
| Anaplasma spp.1 | 4 | 0 | ||||||
| Ehrlichia spp.2 | 7 |
| A. marginale | Borrelia spp. | Leishmania spp. | T. mutans | T. velifera | B. bigemina | B. bovis | Anaplasma spp.1 | Ehrlichia spp.2 | |
|---|---|---|---|---|---|---|---|---|---|
| A. marginale | 111 | 5 | 1 | 3 | 34 | 17 | 2 | 0 | 70* |
| Borrelia spp. | 12 | 0 | 2* | 3 | 0 | 0 | 1 | 5 | |
| Leishmania spp. | 2 | 1* | 1 | 0 | 0 | 0 | 1 | ||
| T. mutans | 4 | 4* | 0 | 0 | 0 | 1 | |||
| T. velifera | 72 | 15* | 1 | 5 | 32 | ||||
| B. bigemina | 35 | 0 | 1 | 14 | |||||
| B. bovis | 2 | 0 | 2 | ||||||
| Anaplasma spp.1 | 22 | 9 | |||||||
| Ehrlichia spp.2 | 134 |
References
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- George, J.E.; Davey, R.B.; Pound, J.M. Introduced ticks and tick-borne diseases: The threat and approaches to eradication. Vet. Clin. N. Am. Food Anim. Pract. 2002, 18, 401–416. [Google Scholar] [CrossRef]
- Gondard, M.; Cabezas-Cruz, A.; Charles, R.A.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Ticks and Tick-Borne Pathogens of the Caribbean: Current Understanding and Future Directions for More Comprehensive Surveillance. Front. Cell Infect. Microbiol. 2017, 7, 490. [Google Scholar] [CrossRef] [PubMed]
- Barré, N.; Uilenberg, G. Spread of parasites transported with their hosts: Case study of two species of cattle tick. Rev. Off. Int. Epizoot. 2010, 29, 149–160, 135–147. [Google Scholar] [PubMed]
- Morel, P.-C. Etudes sur les tiques du bétail en Guadeloupe et Martinique. II. Agents pathogènes transmis par les tiques. Revue d’élevage et de médecine vétérinaire des pays tropicaux 1967, 20, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Uilenberg, G.; Camus, E.; Barré, N. [Existence of Theileria mutans and Theileria velifera (Sporozoa, Theileriidae) in Guadeloupe (French West Indies)]. Rev. Elev. Med. Vet. Pays Trop. 1983, 36, 261–264. [Google Scholar]
- Camus, E.; Barre, N. Vector situation of tick-borne diseases in the Caribbean islands. Vet. Parasitol. 1995, 57, 167–176. [Google Scholar] [CrossRef]
- Alonso, M.; Arellano-Sota, C.; Cereser, V.H.; Cordoves, C.O.; Guglielmone, A.A.; Kessler, R.; Mangold, A.J.; Nari, A.; Patarroyo, J.H.; Solari, M.A. Epidemiology of bovine anaplasmosis and babesiosis in Latin America and the Caribbean. Rev. Sci. Tech. 1992, 11, 713–733. [Google Scholar] [CrossRef] [Green Version]
- Parola, P.; Vestris, G.; Martinez, D.; Brochier, B.; Roux, V.; Raoult, D. Tick-borne rickettiosis in Guadeloupe, the French West Indies: Isolation of Rickettsia africae from Amblyomma variegatum ticks and serosurvey in humans, cattle, and goats. Am. J. Trop. Med. Hyg. 1999, 60, 888–893. [Google Scholar] [CrossRef]
- Camus, E.; Montenegro-James, S. Bovine anaplasmosis and babesiosis in the Lesser Antilles: Risk assessment of an unstable epidemiologic situation. Vet. Res. 1994, 25, 313–317. [Google Scholar]
- Barre, N.; Garris, G.I.; Borel, G.; Camus, E. Hosts and Population Dynamics of Amblyomma variegatum (Acari: Ixodidae) on Guadeloupe, French West Indies. J. Med. Entomol. 1988, 25, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Vachiéry, N.; Jeffery, H.; Pegram, R.; Aprelon, R.; Pinarello, V.; Kandassamy, R.L.Y.; Raliniaina, M.; Molia, S.; Savage, H.; Alexander, R.; et al. Amblyomma variegatum ticks and heartwater on three Caribbean Islands. Ann. N. Y. Acad. Sci. 2008, 1149, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Attali, J.; Raoult, D. First detection of Rickettsia africae on Martinique, in the French West Indies. Ann. Trop. Med. Parasitol. 2003, 97, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.; Lucas, H.; Beati, L.; Yowell, C.; Mahan, S.; Dame, J. Rickettsia africae in Amblyomma variegatum and domestic ruminants on eight Caribbean islands. J. Parasitol. 2010, 96, 1086–1088. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Fournier, P.E.; Fenollar, F.; Jensenius, M.; Prioe, T.; de Pina, J.J.; Caruso, G.; Jones, N.; Laferl, H.; Rosenblatt, J.E.; et al. Rickettsia africae, a Tick-Borne Pathogen in Travelers to Sub-Saharan Africa. N. Engl. J. Med. 2001, 344, 1504–1510. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Gray, J.S.; Kahl, O.; Lane, R.S.; Nijhof, A.M. Research on the ecology of ticks and tick-borne pathogens—methodological principles and caveats. Front. Cell. Infect. Microbiol. 2013, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Cabezas-Cruz, A.; Vayssier-Taussat, M.; Greub, G. Tick-borne pathogen detection: What’s new? Microbes Infect. 2018, 20, 441–444. [Google Scholar] [CrossRef] [Green Version]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef]
- Dupraz, M.; Toty, C.; Devillers, E.; Blanchon, T.; Elguero, E.; Vittecoq, M.; Moutailler, S.; McCoy, K.D. Population structure of the soft tick Ornithodoros maritimus and its associated infectious agents within a colony of its seabird host Larus michahellis. Int. J. Parasitol. Parasites Wildl. 2017, 6, 122–130. [Google Scholar] [CrossRef]
- Gioia, G.V.; Vinueza, R.L.; Marsot, M.; Devillers, E.; Cruz, M.; Petit, E.; Boulouis, H.J.; Moutailler, S.; Monroy, F.; Coello, M.A.; et al. Bovine anaplasmosis and tick-borne pathogens in cattle of the Galapagos Islands. Transbound. Emerg. Dis. 2018, 65, 1262–1271. [Google Scholar] [CrossRef]
- Gondard, M.; Michelet, L.; Nisavanh, A.; Devillers, E.; Delannoy, S.; Fach, P.; Aspan, A.; Ullman, K.; Chirico, J.; Hoffmann, B.; et al. Prevalence of tick-borne viruses in Ixodes ricinus assessed by high-throughput real-time PCR. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmsten, J.; Dalin, A.-M.; Moutailler, S.; Devillers, E.; Gondard, M.; Felton, A. Vector-Borne Zoonotic Pathogens in Eurasian Moose (Alces alces alces). Vector Borne Zoonotic Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ahantarig, A.; Trinachartvanit, W.; Baimai, V.; Grubhoffer, L. Hard ticks and their bacterial endosymbionts (or would be pathogens). Folia Microbiol. (Praha) 2013, 58, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Binetruy, F.; Noël, V.; Cremaschi, J.; McCoy, K.D.; Arnathau, C.; Plantard, O.; Goolsby, J.; Pérez de León, A.A.; Heylen, D.J.A.; et al. Evolutionary changes in symbiont community structure in ticks. Mol. Ecol. 2017, 26, 2905–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, S.; Jouglin, M.; Malandrin, L.; Becker, C.; Agoulon, A.; L’hostis, M.; Chauvin, A. Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology 2007, 134, 197–207. [Google Scholar] [CrossRef]
- Yssouf, A.; Almeras, L.; Raoult, D.; Parola, P. Emerging tools for identification of arthropod vectors. Future Microbiol. 2016, 11, 549–566. [Google Scholar] [CrossRef]
- Diarra, A.Z.; Almeras, L.; Laroche, M.; Berenger, J.-M.; Koné, A.K.; Bocoum, Z.; Dabo, A.; Doumbo, O.; Raoult, D.; Parola, P. Molecular and MALDI-TOF identification of ticks and tick-associated bacteria in Mali. PLoS Negl. Trop. Dis. 2017, 11, e0005762. [Google Scholar] [CrossRef]
- Howell, J.M.; Ueti, M.W.; Palmer, G.H.; Scoles, G.A.; Knowles, D.P. Transovarial Transmission Efficiency of Babesia bovis Tick Stages Acquired by Rhipicephalus (Boophilus) microplus during Acute Infection. J. Clin. Microbiol. 2007, 45, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Chauvin, A.; Moreau, E.; Bonnet, S.; Plantard, O.; Malandrin, L. Babesia and its hosts: Adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet. Res. 2009, 40, 37. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.B.; Eremeeva, M.E.; Olson, P.E.; Thornton, S.A.; Medina, M.J.; Sumner, J.W.; Daschi, G.A. New approaches to detection and identification of Rickettsia africae and Ehrlichia ruminantium in Amblyomma variegatum (Acari: Ixodidae) ticks from the Caribbean. J. Med. Entomol. 2009, 46, 942–951. [Google Scholar] [CrossRef] [Green Version]
- Socolovschi, C.; Huynh, T.P.; Davoust, B.; Gomez, J.; Raoult, D.; Parola, P. Transovarial and trans-stadial transmission of Rickettsiae africae in Amblyomma variegatum ticks. Clin. Microbiol. Infect. 2009, 15 (Suppl. 2), 317–318. [Google Scholar] [CrossRef] [Green Version]
- Maina, A.N.; Jiang, J.; Omulo, S.A.; Cutler, S.J.; Ade, F.; Ogola, E.; Feikin, D.R.; Njenga, M.K.; Cleaveland, S.; Mpoke, S.; et al. High Prevalence of Rickettsia africae Variants in Amblyomma variegatum Ticks from Domestic Mammals in Rural Western Kenya: Implications for Human Health. Vector Borne Zoonotic Dis. 2014, 14, 693–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yssouf, A.; Socolovschi, C.; Kernif, T.; Temmam, S.; Lagadec, E.; Tortosa, P.; Parola, P. First molecular detection of Rickettsia africae in ticks from the Union of the Comoros. Parasites Vectors 2014, 7, 444. [Google Scholar] [CrossRef] [Green Version]
- Matsimbe, A.M.; Magaia, V.; Sanches, G.S.; Neves, L.; Noormahomed, E.; Antunes, S.; Domingos, A. Molecular detection of pathogens in ticks infesting cattle in Nampula province, Mozambique. Exp. Appl. Acarol. 2017, 73, 91–102. [Google Scholar] [CrossRef]
- Rodriguez, O.N.; Espaine, L.; Rivas, A.; Rodriguez, P. Epidemiology of cattle diseases caused by haemoparasites in Cuba. Rev. Cuba. Cienc. Vet. 1989, 20, 37–56. [Google Scholar]
- Woolhouse, M.E.J.; Thumbi, S.M.; Jennings, A.; Chase-Topping, M.; Callaby, R.; Kiara, H.; Oosthuizen, M.C.; Mbole-Kariuki, M.N.; Conradie, I.; Handel, I.G.; et al. Co-infections determine patterns of mortality in a population exposed to parasite infection. Sci. Adv. 2015, 1, e1400026. [Google Scholar] [CrossRef] [Green Version]
- Parola, P.; Inokuma, H.; Camicas, J.L.; Brouqui, P.; Raoult, D. Detection and identification of spotted fever group Rickettsiae and Ehrlichiae in African ticks. Emerg. Infect. Dis. 2001, 7, 1014–1017. [Google Scholar] [CrossRef] [Green Version]
- Inokuma, H.; Beppu, T.; Okuda, M.; Shimada, Y.; Sakata, Y. Detection of ehrlichial DNA in Haemaphysalis ticks recovered from dogs in Japan that is closely related to a novel Ehrlichia sp. found in cattle ticks from Tibet, Thailand, and Africa. J. Clin. Microbiol. 2004, 42, 1353–1355. [Google Scholar] [CrossRef] [Green Version]
- Loftis, A.D.; Reeves, W.K.; Szumlas, D.E.; Abbassy, M.M.; Helmy, I.M.; Moriarity, J.R.; Dasch, G.A. Rickettsial agents in Egyptian ticks collected from domestic animals. Exp. Appl. Acarol. 2006, 40, 67–81. [Google Scholar] [CrossRef]
- Kang, Y.-J.; Diao, X.-N.; Zhao, G.-Y.; Chen, M.-H.; Xiong, Y.; Shi, M.; Fu, W.-M.; Guo, Y.-J.; Pan, B.; Chen, X.-P.; et al. Extensive diversity of Rickettsiales bacteria in two species of ticks from China and the evolution of the Rickettsiales. BMC Evol. Biol. 2014, 14, 167. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Tian, J.-H.; Yu, B.; Guo, W.-P.; Holmes, E.C.; Zhang, Y.-Z. Extensive diversity of rickettsiales bacteria in ticks from Wuhan, China. Ticks Tick Borne Dis. 2017, 8, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.-P.; Tian, J.-H.; Lin, X.-D.; Ni, X.-B.; Chen, X.-P.; Liao, Y.; Yang, S.-Y.; Dumler, J.S.; Holmes, E.C.; Zhang, Y.-Z. Extensive genetic diversity of Rickettsiales bacteria in multiple mosquito species. Sci. Rep. 2016, 6, 38770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yparraguirre, L.A.; Machado-Ferreira, E.; Ullmann, A.J.; Piesman, J.; Zeidner, N.S.; Soares, C.A.G. A hard tick relapsing fever group spirochete in a Brazilian Rhipicephalus (Boophilus) microplus. Vector Borne Zoonotic Dis. 2007, 7, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Hagen, R.M.; Frickmann, H.; Ehlers, J.; Krüger, A.; Margos, G.; Hizo-Teufel, C.; Fingerle, V.; Rakotozandrindrainy, R.; von Kalckreuth, V.; Im, J.; et al. Presence of Borrelia spp. DNA in ticks, but absence of Borrelia spp. and of Leptospira spp. DNA in blood of fever patients in Madagascar. Acta Trop. 2018, 177, 127–134. [Google Scholar] [CrossRef]
- Kumsa, B.; Socolovschi, C.; Raoult, D.; Parola, P. New Borrelia species detected in ixodid ticks in Oromia, Ethiopia. Ticks Tick Borne Dis. 2015, 6, 401–407. [Google Scholar] [CrossRef]
- Ehounoud, C.B.; Yao, K.P.; Dahmani, M.; Achi, Y.L.; Amanzougaghene, N.; Kacou N’Douba, A.; N’Guessan, J.D.; Raoult, D.; Fenollar, F.; Mediannikov, O. Multiple Pathogens Including Potential New Species in Tick Vectors in Côte d’Ivoire. PLoS Negl. Trop. Dis. 2016, 10, e0004367. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A.; Álvarez-Jarreta, J.; Cabezas-Cruz, A. Reservoir and vector evolutionary pressures shaped the adaptation of Borrelia. Infect. Genet. Evol. 2018, 66, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, I.; Fernández, C.; Cinco, M.; Pedroso, R.; Fuentes, O. Do Antiborrelial Antibodies Suggest Lyme Disease in Cuba? Emerg. Infect. Dis. 2004, 10, 1698–1700. [Google Scholar] [CrossRef]
- Rodríguez, I.; Fernández, C.; Sánchez, L.; Martínez, B.; Siegrist, H.H.; Lienhard, R. Prevalence of antibodies to Borrelia burgdorferi sensu stricto in humans from a Cuban village. Braz. J. Infect. Dis. 2012, 16, 82–85. [Google Scholar] [CrossRef] [Green Version]
- Dessau, R.B. Infection due to Borrelia burgdorferi most likely does not occur in Cuba. Braz. J. Infect. Dis. 2012, 16, 404, author reply 405–406. [Google Scholar] [CrossRef] [Green Version]
- Flanigan, T.P.; Schwan, T.G.; Armstrong, C.; Van Voris, L.P.; Salata, R.A. Relapsing Fever in the US Virgin Islands: A Previously Unrecognized Focus of Infection. J. Infect. Dis. 1991, 163, 1391–1392. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Jaimungal, S.; Basdeo-Maharaj, K.; Rao, A.V.C.; Teelucksingh, S. Erythema Migrans–like Illness among Caribbean Islanders. Emerg. Infect. Dis. 2010, 16, 1615–1617. [Google Scholar] [CrossRef] [PubMed]
- Pothirat, T.; Tantiworawit, A.; Chaiwarith, R.; Jariyapan, N.; Wannasan, A.; Siriyasatien, P.; Supparatpinyo, K.; Bates, M.D.; Kwakye-Nuako, G.; Bates, P.A. First Isolation of Leishmania from Northern Thailand: Case Report, Identification as Leishmania martiniquensis and Phylogenetic Position within the Leishmania enriettii Complex. PLoS Negl. Trop. Dis. 2014, 8, e3339. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, O.A.; Serrano, M.G.; Camargo, E.P.; Teixeira, M.M.G.; Shaw, J.J. An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania and Endotrypanum. Parasitology 2018, 145, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Leelayoova, S.; Siripattanapipong, S.; Manomat, J.; Piyaraj, P.; Tan-ariya, P.; Bualert, L.; Mungthin, M. Leishmaniasis in Thailand: A Review of Causative Agents and Situations. Am. J. Trop. Med. Hyg. 2017, 96, 534–542. [Google Scholar] [CrossRef] [Green Version]
- Steverding, D. The history of leishmaniasis. Parasit Vectors 2017, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Desbois, N.; Pratlong, F.; Quist, D.; Dedet, J.-P. Leishmania (Leishmania) martiniquensis n. sp. (Kinetoplastida: Trypanosomatidae), description of the parasite responsible for cutaneous leishmaniasis in Martinique Island (French West Indies). Parasite 2014, 21, 12. [Google Scholar] [CrossRef] [Green Version]
- Liautaud, B.; Vignier, N.; Miossec, C.; Plumelle, Y.; Kone, M.; Delta, D.; Ravel, C.; Cabié, A.; Desbois, N. First case of visceral leishmaniasis caused by Leishmania martiniquensis. Am. J. Trop. Med. Hyg. 2015, 92, 317–319. [Google Scholar] [CrossRef] [Green Version]
- Dantas-Torres, F. Ticks as vectors of Leishmania parasites. Trends Parasitol. 2011, 27, 155–159. [Google Scholar] [CrossRef]
- Rakhshanpour, A.; Malmasi, A.; Mohebali, M.; Nabian, S.; Mirhendi, H.; Zarei, Z.; Dalimi, A.; Mohammadiha, A.; Akhoundi, B.; Azarm, A. Transmission of Leishmania infantum by Rhipicephalus sanguineus (Acari: Ixodidae) in Dogs. Iran. J. Parasitol. 2017, 12, 482–489. [Google Scholar] [PubMed]
- Rojas-Jaimes, J.E.; Correa-Nuñez, G.H.; Rojas, N.; Cáceres-Rey, O. Detection of Leishmania (V) guyanensis in Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) collected from Pecari tajacu. Biomedica 2017, 37, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.Z.; Rahman, M.M.; Akter, S.; Talukder, M.H.; Dey, A.R. An investigation about the possible role of cattle and goats as reservoir hosts for Leishmania donovani in Bangladesh. J. Vector Borne Dis. 2018, 55, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Paixão-Marques, M.D.S.; Alves-Martin, M.F.; Guiraldi, L.M.; Dos Santos, W.J.; de Lemos, F.A.; Sánchez, G.P.; Richini-Pereira, V.B.; Lucheis, S.B. First isolation of Leishmania infantum by blood culture in bovines from endemic area for canine visceral leishmaniasis. Parasitology 2019, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lamattina, D.; Nava, S. Ticks infesting humans in Northern Misiones, Argentina. Medicina (B Aires) 2016, 76, 89–92. [Google Scholar] [PubMed]
- Walker, A.R.; Bouattour, A.; Camicas, J.-L.; Estrada-Pena, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; illustrée.; Bioscience Reports: Wisconsin University- Madison; 2003; ISBN 0-9545173-0-X. Available online: https://pdfs.semanticscholar.org/cdda/35b475e0902e3db7559d5d155fa2b8afd42b.pdf?_ga=2.54794787.921540442.1582955237-1934342711.1568961768 (accessed on 12 February 2020).
- Nielsen, E.M.; Andersen, M.T. Detection and Characterization of Verocytotoxin-Producing Escherichia coli by Automated 5′ Nuclease PCR Assay. J. Clin. Microbiol. 2003, 41, 2884–2893. [Google Scholar] [CrossRef] [Green Version]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-J.; Lee, S.-H.; Park, K.-H.; Koh, Y.-S.; Lee, K.-H.; Baik, H.-S.; Choi, M.-S.; Kim, I.-S.; Jang, W.-J. Evaluation of PCR-Based Assay for Diagnosis of Spotted Fever Group Rickettsiosis in Human Serum Samples. Clin. Diagn. Lab. Immunol. 2005, 12, 759–763. [Google Scholar] [CrossRef] [Green Version]
- Hornok, S.; Földvári, G.; Elek, V.; Naranjo, V.; Farkas, R.; de la Fuente, J. Molecular identification of Anaplasma marginale and rickettsial endosymbionts in blood-sucking flies (Diptera: Tabanidae, Muscidae) and hard ticks (Acari: Ixodidae). Vet. Parasitol. 2008, 154, 354–359. [Google Scholar] [CrossRef]
- Loh, S.-M.; Gofton, A.W.; Lo, N.; Gillett, A.; Ryan, U.M.; Irwin, P.J.; Oskam, C.L. Novel Borrelia species detected in echidna ticks, Bothriocroton concolor, in Australia. Parasit Vectors 2016, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, A.; Campino, L.; Cardoso, L.; Cortes, S. Evaluation of four molecular methods to detect Leishmania infection in dogs. Parasit Vectors 2017, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mixson, T.R.; Campbell, S.R.; Gill, J.S.; Ginsberg, H.S.; Reichard, M.V.; Schulze, T.L.; Dasch, G.A. Prevalence of Ehrlichia, Borrelia, and Rickettsial Agents in Amblyomma americanum (Acari: Ixodidae) Collected from Nine States. J. Med. Entomol. 2006, 43, 1261–1268. [Google Scholar] [CrossRef]
- Trout Fryxell, R.T.; DeBruyn, J.M. The Microbiome of Ehrlichia-Infected and Uninfected Lone Star Ticks (Amblyomma americanum). PLoS ONE 2016, 11, e0146651. [Google Scholar] [CrossRef] [Green Version]
- Aktas, M.; Özübek, S. Transstadial Transmission of Hepatozoon canis by Rhipicephalus sanguineus (Acari: Ixodidae) in Field Conditions. J. Med. Entomol. 2017, 54, 1044–1048. [Google Scholar] [CrossRef]
- Hamšíková, Z.; Silaghi, C.; Rudolf, I.; Venclíková, K.; Mahríková, L.; Slovák, M.; Mendel, J.; Blažejová, H.; Berthová, L.; Kocianová, E.; et al. Molecular detection and phylogenetic analysis of Hepatozoon spp. in questing Ixodes ricinus ticks and rodents from Slovakia and Czech Republic. Parasitol. Res. 2016, 115, 3897–3904. [Google Scholar] [CrossRef]
- Hailemariam, Z.; Krücken, J.; Baumann, M.; Ahmed, J.S.; Clausen, P.-H.; Nijhof, A.M. Molecular detection of tick-borne pathogens in cattle from Southwestern Ethiopia. PLoS ONE 2017, 12, e0188248. [Google Scholar] [CrossRef] [Green Version]
- Laird, J.S.; Kocan, A.A.; Kocan, K.M.; Presley, S.M.; Hair, J.A. Susceptibility of Amblyomma americanum to natural and experimental infections with Theileria cervi. J. Wildl. Dis. 1988, 24, 679–683. [Google Scholar] [CrossRef]
- Mans, B.J.; Pienaar, R.; Latif, A.A. A review of Theileria diagnostics and epidemiology. Int. J. Parasitol. Parasites Wildl. 2015, 4, 104–118. [Google Scholar] [CrossRef] [Green Version]
- Njiiri, N.E.; Bronsvoort, B.M.d.; Collins, N.E.; Steyn, H.C.; Troskie, M.; Vorster, I.; Thumbi, S.M.; Sibeko, K.P.; Jennings, A.; van Wyk, I.C.; et al. The epidemiology of tick-borne haemoparasites as determined by the reverse line blot hybridization assay in an intensively studied cohort of calves in western Kenya. Vet. Parasitol. 2015, 210, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oura, C.a.L.; Bishop, R.P.; Wampande, E.M.; Lubega, G.W.; Tait, A. Application of a reverse line blot assay to the study of haemoparasites in cattle in Uganda. Int. J. Parasitol. 2004, 34, 603–613. [Google Scholar] [CrossRef] [PubMed]






| Microorganisms | Target | Design Name | Sequence (5′ à 3′) | Length (bp) | Controls |
|---|---|---|---|---|---|
| Rickettsia spp. | gltA | Rick_spp_gltA_F | GTCGCAAATGTTCACGGTACTT | 78 | **, Culture of R. slovaca |
| Rick_spp_gltA_R | TCTTCGTGCATTTCTTTCCATTG | ||||
| Rick_spp_gltA_P | TGCAATAGCAAGAACCGTAGGCTGGATG | ||||
| Rickettsia massiliae * | ITS | Ri_ma_ITS_F | GTTATTGCATCACTAATGTTATACTG | 128 | Culture |
| Ri_ma_ITS_R | GTTAATGTTGTTGCACGACTCAA | ||||
| Ri_ma_ITS_P | TAGCCCCGCCACGATATCTAGCAAAAA | ||||
| Rickettsia rickettsii * | ITS | Ri_ri_ITS_F | TCTACTCACAAAGTTATCAGGTTAA | 124 | Plasmid |
| Ri_ri_ITS_R | CCTACGATACTCAGCAAAATAATTT | ||||
| Ri_ri_ITS_P | TCGCTGGATATCGTTGCAGGACTACAG | ||||
| Rickettsia conorii | sca1 | Ri_co_sca1_F | GTAGATGCTTCATAGAATACTGC | 88 | Infected Rhipicephalus sanguineus s.l. |
| Ri_co_sca1_R | CCAAATTTAGTCTACCTTGTGATC | ||||
| Ri_co_sca1_P | TCCTCCTGACGTATTAAAAGAAGCTGAAGCT | ||||
| Rickettsia africae | sca1 | Ri_af_sca1_F | GATACGACAAGTACCTCGCAG | 122 | Culture |
| Ri_af_sca1_R | GGATTATATACTTTAGGTTCGTTAG | ||||
| Ri_af_sca1_P | CAGATAGGAACAGTAATTGTAACGGAACCAG | ||||
| Rickettsia felis | orfB | Ri_fel_orfB_F | ACCCTTTTCGTAACGCTTTGC | 163 | Culture |
| Ri_fel_orfB_R | TATACTTAATGCTGGGCTAAACC | ||||
| Ri_fel_orfB_P | AGGGAAACCTGGACTCCATATTCAAAAGAG | ||||
| Rickettsia typhi | ompB | Ri_typ_ompB_F | CAGGTCATGGTATTACTGCTCA | 133 | Culture |
| Ri_typ_ompB_R | GCAGCAGTAAAGTCTATTGATCC | ||||
| Ri_typ_ompB_P | ACAAGCTGCTACTACAAAAAGTGCTCAAAATG | ||||
| Rickettsia prowazekii | gltA | Ri_pro_gltA_F | CAAGTATCGGTAAAGATGTAATCG | 151 | Plasmid |
| Ri_pro_gltA_R | TATCCTCGATACCATAATATGCC | ||||
| Ri_pro_gltA_P | ATATAAGTAGGGTATCTGCGGAAGCCGAT | ||||
| Borrelia spp. * | 23S rRNA | Bo_bu_sl_23S_F | GAGTCTTAAAAGGGCGATTTAGT | 73 | **, Culture of B. afzelii, B. garinii, B. valaisiana, B. lusitaniae, B. recurrentis |
| Bo_bu_sl-23S_R | CTTCAGCCTGGCCATAAATAG | ||||
| Bo_bu_sl_23S_P | TAGATGTGGTAGACCCGAAGCCGAGT | ||||
| Borrelia burgdorferi sensu stricto | glpA | Bo_bu_glpA_F | GCAATTACAAGGGGGTATAAAGC | 206 | Culture |
| Bo_bu_glpA_R | GGCGTGATAAGTGCACATTCG | ||||
| Bo_bu_glpA_P | TTAATTAAACGGGGTGCATTCTTCTCAAGAATG | ||||
| Borrelia anserina | fla | Bor_ans_fla_F | GGAGCACAACAAGAGGGAG | 76 | Plasmid |
| Bor_ans_fla_R | TTGGAGAATTAACCCCACCTG | ||||
| Bor_ans_fla_P | TGCAAGCAACTCCAGCTCCAGTAGCT | ||||
| Borrelia lonestari | glpQ | Bor_lon_glpQ_F | GATCCAGAACTTGATACAACCAC | 99 | Infected Amblyomma americanum |
| Bor_lon_glpQ_R | TTCATCTAGTGAGAAGTCAGTAG | ||||
| Bor_lon_glpQ_P | AGTAATATCGTCCGTCTTCCCTAGCTCG | ||||
| Borrelia parkeri | gyrB | Bor_par_gyrB_F | GCAAAACGATTCAAAGTGAGTCC | 184 | Culture |
| Bor_par_gyrB_R | CTCATTGCCTTTAAGAAACCACTT | ||||
| Bor_par_gyrB_P | TTAAAACCAGCAACATGAGTTCCTCCTTCTC | ||||
| Borrelia bissettii * | rpoB | Bo_bi_rpoB_F | GCAACCAGTCAGCTTTCACAG | 118 | Plasmid |
| Bo_bi_rpoB_R | CAAATCCTGCCCTATCCCTTG | ||||
| Bo_bi_rpoB_P | AAAGTCCTCCCGGCCCAAGAGCATTAA | ||||
| Borrelia theileri | glpQ | Bo_th_glpQ_F | GTGCTAACAAAGGACAATATTCC | 213 | Plasmid |
| Bo_th_glpQ_R | GGTTAGTGGAAAACGGTTAGGAT | ||||
| Bo_th_glpQ_P | TATTATAATTCACGAGCCAGAGCTTGACAC | ||||
| Bartonella spp. | ssrA | Bart_spp_ssrA_F | CGTTATCGGGCTAAATGAGTAG | 118 | **, Culture of B. quintana |
| Bart_spp_ssrA_R | ACCCCGCTTAAACCTGCGA | ||||
| Bart_spp_ssrA_P | TTGCAAATGACAACTATGCGGAAGCACGTC | ||||
| Bartonella barcilliformis * | rpoB | Ba_ba_rpoB_F | GAAGAGTTTGTAGTTTGTCGTCA | 105 | Culture |
| Ba_ba_rpoB_R | AGCAGCTACAGAAACCAACTG | ||||
| Ba_ba_rpoB_P | TGCAGGTGAAGTTTTGATGGTGCCACG | ||||
| Bartonella henselae | ribC | Bar_he_ribC_F | GGGATGCGATTTAATAGTTCTAC | 116 | Culture |
| Bar_he_ribC_R | CGCTTGTTGTTTTGATCCTCG | ||||
| Bar_he_ribC_P | ACGTTATAGTAGCGAAAACTTAGAAATTGGTGC | ||||
| Bartonella vinsonii subsp. berkhoffii | ITS | Bar_vin_ITS_F | GGAATTGCTTAACCCACTGTTG | 141 | Culture |
| Bar_vin_ITS_R | CCTTATTGATTTAGATCTGATGGG | ||||
| Bar_vin_ITS_P2 | AGAAACTCCCGCCTTTATGAGAGAAATCTCT | ||||
| Coxiella burnetii and Coxiella-like * | Icd | Co_bu_icd_F | AGGCCCGTCCGTTATTTTACG | 74 | Culture |
| Co_bu_icd_R | CGGAAAATCACCATATTCACCTT | ||||
| Co_bu_icd_P | TTCAGGCGTTTTGACCGGGCTTGGC | ||||
| IS1111 | Co_bu_IS111_F | TGGAGGAGCGAACCATTGGT | 86 | Culture | |
| Co_bu_IS111_R | CATACGGTTTGACGTGCTGC | ||||
| Co_bu_IS111_P | ATCGGACGTTTATGGGGATGGGTATCC | ||||
| Francisella tularensis and Francisella-like endosymbionts * | tul4 | Fr_tu_tul4_F | ACCCACAAGGAAGTGTAAGATTA | 76 | Culture |
| Fr_tu_tul4_R | GTAATTGGGAAGCTTGTATCATG | ||||
| Fr_tu_tul4_P | AATGGCAGGCTCCAGAAGGTTCTAAGT | ||||
| fopA | Fr_tu_fopA_F | GGCAAATCTAGCAGGTCAAGC | 91 | Culture | |
| Fr_tu_fopA_R | CAACACTTGCTTGAACATTTCTAG | ||||
| Fr_tu_fopA_P | AACAGGTGCTTGGGATGTGGGTGGTG | ||||
| Anaplasma spp. | 16S rRNA | Ana_spp_16S_F | CTTAGGGTTGTAAAACTCTTTCAG | 160 | ** |
| Ana_spp_16S_R | CTTTAACTTACCAAACCGCCTAC | ||||
| Ana_spp_16S_P | ATGCCCTTTACGCCCAATAATTCCGAACA | ||||
| Anaplasma marginale * | msp1b | An_ma_msp1_F | CAGGCTTCAAGCGTACAGTG | 85 | Experimentally infected bovine blood sample |
| An_ma_msp1_R | GATATCTGTGCCTGGCCTTC | ||||
| An_ma_msp1_P | ATGAAAGCCTGGAGATGTTAGACCGAG | ||||
| Anaplasma phagocytophilum * | msp2 | An_ph_msp2_F | GCTATGGAAGGCAGTGTTGG | 77 | Infected Ixodes spp. tick |
| An_ph_msp2_R | GTCTTGAAGCGCTCGTAACC | ||||
| An_ph_msp2_P | AATCTCAAGCTCAACCCTGGCACCAC | ||||
| Anaplasma platys * | groEL | An_pla_groEL_F | TTCTGCCGATCCTTGAAAACG | 75 | Infected canine blood sample |
| An_pla_groEL_R | CTTCTCCTTCTACATCCTCAG | ||||
| An_pla_groEL_P | TTGCTAGATCCGGCAGGCCTCTGC | ||||
| Anaplasma bovis * | groEL | An_bo_groEL_F | GGGAGATAGTACACATCCTTG | 73 | Plasmid |
| An_bo_groEL_R | CTGATAGCTACAGTTAAGCCC | ||||
| An_bo_groEL_P | AGGTGCTGTTGGATGTACTGCTGGACC | ||||
| Anaplasma ovis * | msp4 | An_ov_msp4_F | TCATTCGACATGCGTGAGTCA | 92 | Plasmid |
| An_ov_msp4_r | TTTGCTGGCGCACTCACATC | ||||
| An_ov_msp4_P | AGCAGAGAGACCTCGTATGTTAGAGGC | ||||
| Ehrlichia spp. * | 16S rRNA | Neo_mik_16S_F | GCAACGCGAAAAACCTTACCA | 98 | ** |
| Neo_mik_16S_R | AGCCATGCAGCACCTGTGT | ||||
| Neo_mik_16S_P | AAGGTCCAGCCAAACTGACTCTTCCG | ||||
| Ehrlichia canis | gltA | Eh_ca_gltA_F | GACCAAGCAGTTGATAAAGATGG | 136 | Culture |
| Eh_ca_gltA_R | CACTATAAGACAATCCATGATTAGG | ||||
| Eh_ca_gltA_P | ATTAAAACATCCTAAGATAGCAGTGGCTAAGG | ||||
| Ehrlichia chaffeensis * | dsb | Eh_ch_dsb_F | TATTGCTAATTACCCTCAAAAAGTC | 117 | Infected Amblyomma americanum |
| Eh_ch_dsb_R | GAGCTATCCTCAAGTTCAGATTT | ||||
| Eh_ch_dsb_P | ATTGACCTCCTAACTAGAGGGCAAGCA | ||||
| Ehrlichia ewingii * | dsb | Eh_ew_dsb_F | CAATACTTGGAGAAGCATCATTG | 111 | Infected Amblyomma americanum |
| Eh_ew_dsb_R | TTGCTTATGGCTTAATGCTGCAT | ||||
| Eh_ew_dsb_P | AAAGCAGTACGTGCAGCATTGGCTGTA | ||||
| Ehrlichia ruminantium | gltA | Eh_ru_gltA_F | CCAGAAAACTGATGGTGAGTTAG | 116 | Culture |
| Eh_ru_gltA_R | AGCCTACATCAGCTTGAATGAAG | ||||
| Eh_ru_gltA_P | AGTGTAAACTTGCTGTTGCTAAGGTAGCATG | ||||
| Panola Mountain Ehrlichia | gltA | Eh_PME_gltA_F | GCTAGTTATGAGTTAGAATGTAAAC | 121 | Infected Amblyomma americanum |
| Eh_PME_gltA_R | TACTATAGGATAATCTTGAATCAGC | ||||
| Eh_PME_gltA_P | TTGCTATCGCTAAAATTCCAAGTATGATTGCG | ||||
| Neoehrlichia mikurensis * | groEL | Neo_mik_groEL_F | AGAGACATCATTCGCATTTTGGA | 96 | Infected rodent blood sample |
| Neo_mik_groEL_R | TTCCGGTGTACCATAAGGCTT | ||||
| Neo_mik_groEL_P | AGATGCTGTTGGATGTACTGCTGGACC | ||||
| Aegyptianella pullorum | groEL | Ae_pul_groEL_F | AGCCAGTATTATCGCTCAAGG | 168 | Plasmid |
| Ae_pul_groEL_R | GCCTCACGTGCCTTCATAAC | ||||
| Ae_pul_groEL_P | TGCTTCTCAGTGTAACGACAGGGTTGG | ||||
| Apicomplexa | 18S rRNA | Apic_18S_F | TGAACGAGGAATGCCTAGTATG | 104 | **, Infected canine blood sample, with B. canis rossi, B. canis canis; Culture of B. divergens, T. lestoquari, T. annulata |
| Apic_18S_R | CACCGGATCACTCGATCGG | ||||
| Apic_18S_S | TAGGAGCGACGGGCGGTGTGTAC | ||||
| Babesia canis vogeli * | hsp70 | Ba_vo_hsp70_F | TCACTGTGCCTGCGTACTTC | 87 | Infected canine blood sample |
| Ba_vo_hsp70_R | TGATACGCATGACGTTGAGAC | ||||
| Ba_vo_hsp70_P | AACGACTCCCAGCGCCAGGCCAC | ||||
| Babesia ovis * | 18S rRNA | Ba_ov_18S_F | TCTGTGATGCCCTTAGATGTC | 92 | Plasmid |
| Ba_ov_18S_R | GCTGGTTACCCGCGCCTT | ||||
| Ba_ov_18S_P | TCGGAGCGGGGTCAACTCGATGCAT | ||||
| Babesia bigemina * | 18S rRNA | Ba_big_RNA18S_F | ATTCCGTTAACGAACGAGACC | 99 | Plasmid |
| Ba_big_RNA18S_R | TTCCCCCACGCTTGAAGCA | ||||
| Ba_big_RNA18S_P | CAGGAGTCCCTCTAAGAAGCAAACGAG | ||||
| Babesia gibsoni | Rap1 | Ba_gib_rap1_F | CTCTTGCTCATCATCTTTTCGG | 130 | Plasmid |
| Ba_gib_rap1_R | TCAGCGTATCCATCCATTATATG | ||||
| Ba_gib_rap1_S | TTTAATGCGTGCTACGTTGTACTTCCCAAAG | ||||
| Babesia caballi * | Rap1 | Ba_cab_rap1_F | GTTGTTCGGCTGGGGCATC | 94 | Plasmid |
| Ba_cab_rap1_R | CAGGCGACTGACGCTGTGT | ||||
| Ba_cab_rap1_P | TCTGTCCCGATGTCAAGGGGCAGGT | ||||
| Babesia bovis * | CCTeta | Ba_bo_CCTeta_F | GCCAAGTAGTGGTAGACTGTA | 100 | Plasmid |
| Ba_bo_CCTeta_R | GCTCCGTCATTGGTTATGGTA | ||||
| Ba_bo_CCTeta_P | TAAAGACAACACTGGGTCCGCGTGG | ||||
| Babesia duncani * | ITS2 | Ba_du_ITS_F | ATTTCCGTTTGCGAGAGTTGC | 87 | Plasmid |
| Ba_du_ITS_R | AGGAAGCATCAAGTCATAACAAC | ||||
| Ba_du_ITS_P | AACAAGAGGCCCCGAGATCAAGGCAA | ||||
| Babesia microti * | CCTeta | Bab_mi_CCTeta_F | ACAATGGATTTTCCCCAGCAAAA | 145 | Culture |
| Bab_mi_CCTeta_R | GCGACATTTCGGCAACTTATATA | ||||
| Bab_mi_CCTeta_P | TACTCTGGTGCAATGAGCGTATGGGTA | ||||
| Theileria parva * | 18S rRNA | Th_pa_18S_F | GAGTATCAATTGGAGGGCAAG | 173 | Culture |
| Th_pa_18S_R | CAGACAAAGCGAACTCCGTC | ||||
| Th_pa_18S_P | AAATAAGCCACATGCAGAGACCCCGAA | ||||
| Theileria mutans | ITS | The_mu_ITS_F | CCTTATTAGGGGCTACCGTG | 119 | Plasmid |
| The_mu_ITS_R | GTTTCAAATTTGAAGTAACCAAGTG | ||||
| The_mu_ITS_P | ATCCGTGAAAAACGTGCCAAACTGGTTAC | ||||
| Theileria velifera | 18S rRNA | The_ve_18S_F | TGTGGCTTATCTGGGTTCGC | 151 | Plasmid |
| The_ve_18S_R | CCATTACTTTGGTACCTAAAACC | ||||
| The_ve_18S_P | TTGCGTTCCCGGTGTTTTACTTTGAGAAAG | ||||
| Theileria equi | ema1 | Th_eq_ema1_F4 | CGGCAAGAAGCACACCTTC | 167 | Plasmid |
| Th_eq_ema1_R4 | TGCCATCGCCCTTGTAGAG | ||||
| Th_eq_ema1_P4 | AAGGCTCCAGGCAAGCGCGTCCT | ||||
| Cytauxzoon felis | ITS2 | Cy_fel_ITS2_F | AAGATCCGAACGGAGTGAGG | 119 | Plasmid |
| Cy_fel_ITS2_R | GTAGTCTCACCCAATTTCAGG | ||||
| Cy_fel_ITS2_S | AAGTGTGGGATGTACCGACGTGTGAG | ||||
| Hepatozoon spp. | 18S rRNA | Hepa_spp_18S_F | ATTGGCTTACCGTGGCAGTG | 175 | ** |
| Hepa_spp_18S_R | AAAGCATTTTAACTGCCTTGTATTG | ||||
| Hepa_spp_18S_S | ACGGTTAACGGGGGATTAGGGTTCGAT | ||||
| Hepatozoon canis | 18S rRNA | He_can_18S_F | TTCTAACAGTTTGAGAGAGGTAG | 221 | Infected canine blood sample |
| He_can_18S_R | AGCAGACCGGTTACTTTTAGC | ||||
| He_can_18S_S | AGAACTTCAACTACGAGCTTTTTAACTGCAAC | ||||
| Hepatozoon americanum | 18S rRNA | He_ame_18S_F2 | GGTATCATTTTGGTGTGTTTTTAAC | 159 | Plasmid |
| He_ame_18S_R2 | CTTATTATTCCATGCTCCAGTATTC | ||||
| He_ame_18S_P2 | AAAAGCGTAAAAGCCTGCTAAAAACACTCTAC | ||||
| Leishmania spp. | hsp70 | Leish_spp_hsp70_F | CGACCTGTTCCGCAGCAC | 78 | ** and culture of L. martiniquensis |
| Leish_spp_hsp70_R | TCGTGCACGGAGCGCTTG | ||||
| Leish_spp_hsp70_S | TCCATCTTCGCGTCCTGCAGCACG | ||||
| Leishmania infantum | ITS | Le_inf_ITS_F | CGCACCGCCTATACAAAAGC | 103 | Culture |
| Le_inf_ITS_R | GTTATGTGAGCCGTTATCCAC | ||||
| Le_inf_ITS_S | ACACGCACCCACCCCGCCAAAAAC | ||||
| Rangelia vitalii | 18S rRNA | Ra_vit_18S_F | TAACCGTGCTAATTGTAGGGC | 92 | Plasmid |
| Ra_vit_18S_R | GAATCACCAAACCAAATGGAGG | ||||
| Ra_vit_18S_S | TAATACACGTTCGAGGGCGCGTTTTGC | ||||
| Tick spp. | 16S rRNA | Tick_spp_16S_F | AAATACTCTAGGGATAACAGCGT | 99 | ** |
| Tick_spp_16S_R | TCTTCATCAAACAAGTATCCTAATC | ||||
| Tick_spp_16S_P | CAACATCGAGGTCGCAAACCATTTTGTCTA | ||||
| Amblyomma variegatum | ITS2 | Amb_var_ITS2_F | GCCAGCCTCTGAAGTGACG | 117 | Tick extract (Guadeloupe) |
| Amb_var_ITS2_R | TTCTGCGGTTTAAGCGACGC | ||||
| Amb_var_ITS2_P | TCTTGCCACTCGACCCGTGCCTC | ||||
| Rhipicephalus microplus | ITS2 | Rhi_mic_ITS2_F | GCTTAAGGCGTTCTCGTCG | 144 | Tick extract (Galapagos Islands) |
| Rhi_mic_ITS2_R | CAAGGGCAGCCACGCAG | ||||
| Rhi_mic_ITS2_P | TAGTCCGCCGTCGGTCTAAGTGCTTC | ||||
| Rhipicephalus sanguineus sensu lato | ITS2 | Rhi_san_ITS2_F | TTGAACGCTACGGCAAAGCG | 110 | Tick extract (France) |
| Rhi_san_ITS2_R | CCATCACCTCGGTGCAGTC | ||||
| Rhi_san_ITS2_P | ACAAGGGCCGCTCGAAAGGCGAGA |
| Biomark Id | Sequence Name | T | S | AN | L | Closest Homology | Id% | Reference |
|---|---|---|---|---|---|---|---|---|
| Rickettsia spp. | Rickettsia africae Tick208 | 30 | 14 | MK049851 | 248 | Rickettsia africae | 100 | AF123706.1 |
| Leishmania spp. | Leishmania martiniquensis Tick389 | 2 | 1 | MK049850 | 272 | Leishmania martiniquensis | 100 | AF303938.1 |
| Leishmania siamensis | 100 | GQ226033.1 | ||||||
| Borrelia spp. | Borrelia sp. Tick7 | 30 | 1 | MK049846 | 245 | Borrelia anserina | 90 | X75201.1 |
| Borrelia sp. Tick457 | 4 | MK049847 | 327 | Borrelia sp. BR | 100 | EF141022.1 | ||
| Borrelia sp. strain Mo063b-flaB | 100 | KY070335.1 | ||||||
| Borrelia theileri | 99 | KF569936.1 | ||||||
| Anaplasma spp. | Anaplasma sp. Tick314 | 2 | 2 | MK049845 | 245 | Candidatus Anaplasma boleense | 100 | KX987335.1 |
| Anaplasma marginale | Anaplasma sp. Tick283 | 2 | 2 | MK049844 | 244 | Anaplasma marginale | 100 | MH155593.1 |
| Anaplasma centrale | 100 | MF289482.1 | ||||||
| Anaplasma ovis | 100 | MG770440.1 | ||||||
| Anaplasma capra | 100 | MF000917.1 | ||||||
| Anaplasma phagocytophilum | 100 | DQ648489.1 | ||||||
| Ehrlichia spp. | Ehrlichia sp. Tick428 | 2 | 2 | MK049849 | 246 | Ehrlichia spp. | 100 | KY594915.1 |
| Ehrlichia canis | 99 | KY594915.1 | ||||||
| Ehrlichia ewingii | 99 | U96436.1 | ||||||
| Ehrlichia chaffeensis | 99 | NR_074500.2 | ||||||
| Ehrlichia muris | 99 | KU535865.1 | ||||||
| Ehrlichia minasensis | 99 | NR_148800.1 | ||||||
| Ehrlichia ruminantium | Ehrlichia ruminantium Tick116 | 1 | 1 | MK049848 | 207 | Ehrlichia ruminantium | 100 | NR_074155.1 |
| Babesia bigemina | Babesia bigemina Tick222 | 2 | 1 | MK071738 | 99 | Babesia bigemina | 100 | KP710227.1 |
| Babesia bovis | Babesia bovis Tick497 | 2 | 2 | MK071739 | 100 | Babesia bovis | 99 | AB367921.1 |
| Pathogen | Targeted Gene | Primer Name | Sequence (5′ → 3′) | Length (bp) | References |
|---|---|---|---|---|---|
| Rickettsia spp. | gltA | Rsfg877 | GGGGGCCTGCTCACGGCGG | 381 | [67] |
| Rsfg1258 | ATTGCAAAAAGTACAGTGAACA | ||||
| ompB | Rc.rompB.4362p | GTCAGCGTTACTTCTTCGATGC | 475 | [68] | |
| Rc.rompB.4836n | CCGTACTCCATCTTAGCATCAG | ||||
| Rc.rompB.4496p | CCAATGGCAGGACTTAGCTACT | 267 | |||
| Rc.rompB.4762n | AGGCTGGCTGATACACGGAGTAA | ||||
| Anaplasma/Ehrlichia spp. | 16S rRNA | EHR16SD | GGTACCYACAGAAGAAGTCC | 345 | [69] |
| EHR16SR | TAGCACTCATCGTTTACAGC | ||||
| Borrelia spp. | flaB | FlaB280F | GCAGTTCARTCAGGTAACGG | 645 | [70] |
| FlaRL | GCAATCATAGCCATTGCAGATTGT | ||||
| FlaB737F | GCATCAACTGTRGTTGTAACATTAACAGG | 407 | |||
| FlaLL | ACATATTCAGATGCAGACAGAGGT | ||||
| Leishmania spp. | SSU rRNA | R221 | GGTTCCTTTCCTGATTTACG | 603 | [71] |
| R332 | GGCCGGTAAAGGCCGAATAG | ||||
| R223 | TCCATCGCAACCTCGGTT | 358 | |||
| R333 | AAAGCGGGCGCGGTGCTG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gondard, M.; Delannoy, S.; Pinarello, V.; Aprelon, R.; Devillers, E.; Galon, C.; Pradel, J.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Upscaling the Surveillance of Tick-Borne Pathogens in the French Caribbean Islands. Pathogens 2020, 9, 176. https://doi.org/10.3390/pathogens9030176
Gondard M, Delannoy S, Pinarello V, Aprelon R, Devillers E, Galon C, Pradel J, Vayssier-Taussat M, Albina E, Moutailler S. Upscaling the Surveillance of Tick-Borne Pathogens in the French Caribbean Islands. Pathogens. 2020; 9(3):176. https://doi.org/10.3390/pathogens9030176
Chicago/Turabian StyleGondard, Mathilde, Sabine Delannoy, Valérie Pinarello, Rosalie Aprelon, Elodie Devillers, Clémence Galon, Jennifer Pradel, Muriel Vayssier-Taussat, Emmanuel Albina, and Sara Moutailler. 2020. "Upscaling the Surveillance of Tick-Borne Pathogens in the French Caribbean Islands" Pathogens 9, no. 3: 176. https://doi.org/10.3390/pathogens9030176