NcPuf1 Is a Key Virulence Factor in Neospora caninum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell and Parasite Culture
2.2. Construction of Transgenic Parasite Strains
2.3. Western Blotting Assay and Immunofluorescence Assays
2.4. Quantitative Real-Time PCR
2.5. Plaque Assay and Parasite Intracellular Replication Assay
2.6. Bradyzoite Differentiation Assay In Vitro
2.7. Animal Infection Experiments
2.8. Statistical Analysis
3. Results
3.1. NcPuf1 Protein Is Expressed in N. caninum
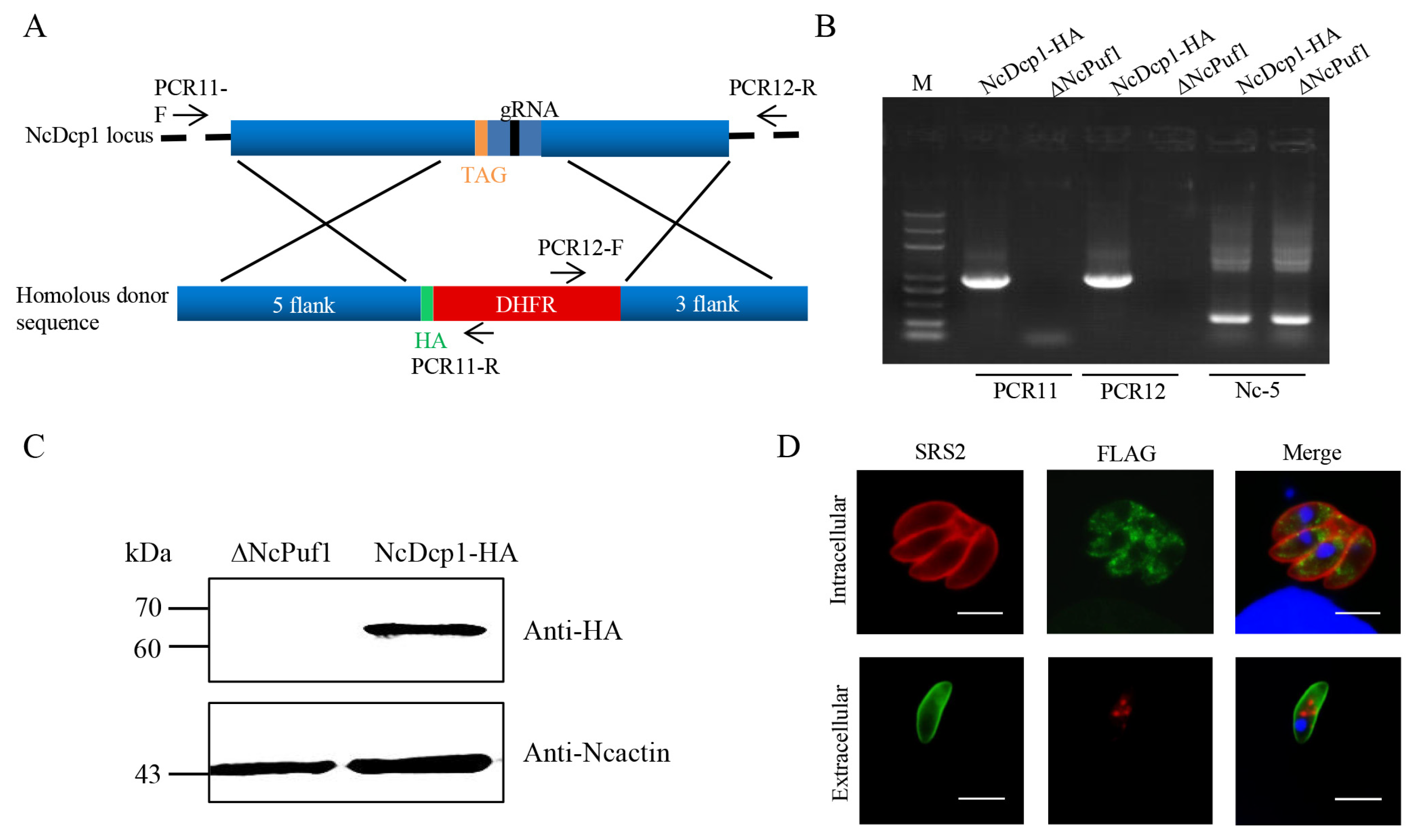
3.2. NcPuf1 Is Recruited into Processing Bodies in Extracellular Parasites
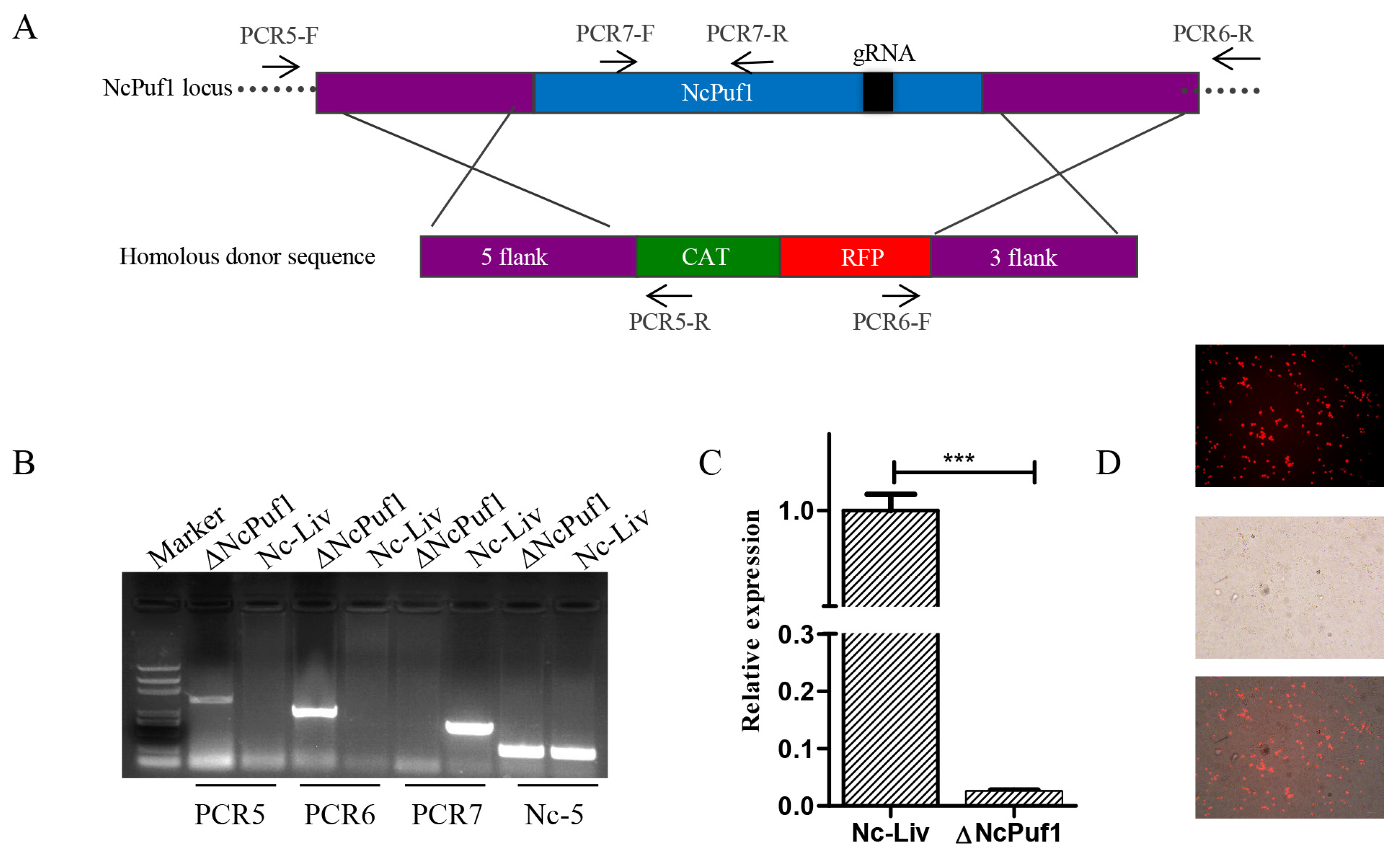
3.3. NcPuf1 Knockout and Complementary Strains
3.4. NcPuf1 Is Vital for Parasite Growth In Vitro and Virulence in Mice
3.5. NcPuf1 Is Not Required for the Formation of P-Bodies in N. caninum
3.6. ΔNcPuf1 does Not Affect the Differentiation from Tachyzoites to Bradyzoites In Vitro and Tissue Cysts Formation In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Schares, G.; Ortega-Mora, L.M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 2007, 20, 323. [Google Scholar] [CrossRef] [Green Version]
- Reichel, M.P.; Ellis, J.T. Neospora caninum--how close are we to development of an efficacious vaccine that prevents abortion in cattle? Int. J. Parasitol. 2009, 39, 1173–1187. [Google Scholar] [CrossRef] [Green Version]
- Dubey, J.P.; Buxton, D.; Wouda, W. Pathogenesis of bovine neosporosis. J. Comp. Pathol. 2006, 134, 267–289. [Google Scholar] [CrossRef]
- Reichel, M.P.; Alejandra Ayanegui-Alcérreca, M.; Gondim, L.F.; Ellis, J.T. What is the global economic impact of Neospora caninum in cattle—The billion dollar question. Int. J. Parasitol. 2013, 43, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Goodswen, S.J.; Kennedy, P.J.; Ellis, J.T. A review of the infection, genetics, and evolution of Neospora caninum: From the past to the present. Infect. Genet. Evol. 2013, 13, 133–150. [Google Scholar] [CrossRef]
- Kang, S.W.; Kweon, C.H.; Lee, E.H.; Choe, S.E.; Jung, S.C.; Dong, V.Q. The differentiation of transcription between tachyzoites and bradyzoites of in vitro cultured Neospora caninum. Parasitol. Res. 2008, 103, 1011–1018. [Google Scholar] [CrossRef]
- Kang, S.W.; Lee, E.H.; Jean, Y.H.; Choe, S.E.; Dong, V.Q.; Lee, M.S. The differential protein expression profiles and immunogenicity of tachyzoites and bradyzoites of in vitro cultured Neospora caninum. Parasitol. Res. 2008, 103, 905–913. [Google Scholar] [CrossRef]
- Glisovic, T.; Bachorik, J.L.; Yong, J.; Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008, 582, 1977–1986. [Google Scholar] [CrossRef] [Green Version]
- Wickens, M.; Bernstein, D.S.; Kimble, J.; Parker, R. A PUF family portrait: 3′UTR regulation as a way of life. Trends Genet. 2002, 18, 150–157. [Google Scholar] [CrossRef]
- Qiu, C.; Mccann, K.L.; Wine, R.N.; Baserga, S.J.; Hall, T.M.T. A divergent pumilio repeat protein family for pre-rRNA processing and mRNA localization. Proc. Natl. Acad. Sci. USA 2014, 111, 18554–18559. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.; Matuschewski, K.; Silvie, O. The puf-Family RNA-Binding protein puf2 controls sporozoite conversion to liver stages in the malaria parasite. PLoS ONE 2011, 6, e19860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Miao, J.; Liu, T.; Sullivan, W.J.; Cui, L.; Chen, X. Characterization of TgPuf1, a member of the Puf family RNA-binding proteins from Toxoplasma gondii. Parasit. Vectors 2014, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomessantos, C.S.S.; Braks, J.A.M.; Prudencio, M.; Carret, C.; Gomes, A.R.; Pain, A.; Feltwell, T.; Khan, S.M.; Waters, A.P.; Janse, C.J. Transition of Plasmodium sporozoites into liver stage-like forms is regulated by the RNA binding protein pumilio. PLoS Pathog. 2011, 7, e1002046. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Liu, J.; Ma, L.; Zhang, X.; Zhang, X.; Zhou, B.; Zhu, X.; Liu, Q. NcGRA17 is an important regulator of parasitophorous vacuole morphology and pathogenicity of Neospora caninum. Vet. Parasitol. 2018, 264, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Brown, K.M.; Lee, T.D.; Sibley, L.D. Efficient gene disruption in diverse strains of Toxoplasma gondii using CRISPR/CAS9. mBio 2014, 5, e01114. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Liu, G.; Liu, J.; Li, M.; Zhang, H.; Tang, D.; Liu, Q. Neospora caninum ROP16 play an important role in the pathogenicity by phosphorylating host cell STAT3. Vet. Parasitol. 2017, 243, 135–147. [Google Scholar] [CrossRef]
- Song, X.; Lin, M.; Li, M.; Yang, X.; Liu, J.; Liu, Q. Toxoplasma gondii metacaspase 2 is an important factor that influences bradyzoite formation in the Pru strain. Parasitol. Res. 2020, 119, 2287–2298. [Google Scholar] [CrossRef]
- Weiss, L.M.; Ma, Y.F.; Halonen, S.K.; Mcallister, M.M.; Zhang, Y.W. The in vitro development of Neospora caninum bradyzoites. Int. J. Parasitol. 1999, 29, 1713–1723. [Google Scholar] [CrossRef] [Green Version]
- Lirussi, D.; Matrajt, M. RNA granules present only in extracellular Toxoplasma Gondii increase parasite viability. Int. J. Biol. Sci. 2011, 7, 960–967. [Google Scholar] [CrossRef] [Green Version]
- Lall, S.; Piano, F.; Davis, R.E. Caenorhabditis elegans decapping proteins: Localization and functional analysis of Dcp1, Dcp2, and DcpS during embryogenesis. Mol. Biol. Cell 2005, 16, 5880–5890. [Google Scholar] [CrossRef] [Green Version]
- Sheth, U.; Parker, R. Decapping and decay of messenger RNA occur in cytoplasmic processing bodies. Science 2003, 300, 805–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Lei, T.; Liu, J.; Li, M.; Nan, H.; Liu, Q. A nuclear factor of high mobility group box protein in Toxoplasma gondii. PLoS ONE 2014, 9, e111993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schorer, M.; Debache, K.; Barna, F.; Monney, T.; Müller, J.; Boykin, D.W.; Stephens, C.E.; Hemphill, A. Di-cationic arylimidamides act against Neospora caninum tachyzoites by interference in membrane structure and nucleolar integrity and are active against challenge infection in mice. Int. J. Parasitol. Drugs Drug Resist. 2012, 2, 109–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Hart, K.J.; Dong, G.; Siddiqui, F.A.; Sebastian, A.; Li, X.; Albert, I.; Miao, J.; Lindner, S.E.; Cui, L. Puf3 participates in ribosomal biogenesis in malaria parasites. J. Cell Sci. 2018, 131, jcs212597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droll, D.; Archer, S.; Fenn, K.; Delhi, P.; Matthews, K.; Clayton, C. The trypanosome pumilio-domain protein PUF7 associates with a nuclear cyclophilin and is involved in ribosomal RNA maturation. FEBS Lett. 2010, 584, 1156–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassola, A.; De Gaudenzi, J.G.; Frasch, A.C. Recruitment of mRNAs to cytoplasmic ribonucleoprotein granules in trypanosomes. Mol. Microbiol. 2007, 65, 655–670. [Google Scholar] [CrossRef]
- Cassola, A. RNA granules living a post-transcriptional life: The trypanosome’s case. Curr. Chem. Biol. 2011, 5. [Google Scholar] [CrossRef]
- Voronina, E.; Paix, A.; Seydoux, G. The P granule component PGL-1 promotes the localization and silencing activity of the PUF protein FBF-2 in germline stem cells. Development 2012, 139, 3732–3740. [Google Scholar] [CrossRef] [Green Version]
- Hubstenberger, A.; Courel, M.; Bénard, M.; Souquere, S.; Ernoult-Lange, M.; Chouaib, R.; Yi, Z.; Morlot, J.-B.; Munier, A.; Fradet, M.; et al. P-Body purification reveals the condensation of repressed mRNA regulons. Mol. Cell. 2017, 68, 144–157. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, W.-Y.; Wang, Y.-T.; Wang, S.-W. Fission yeast Puf2, a Pumilio and FBF family of RNA-binding protein, links stress granules to processing bodies. Mol. Cell Biol. 2020, 40, e00589-19. [Google Scholar] [CrossRef]
- Vessey, J.P.; Vaccani, A.; Xie, Y.; Dahm, R.; Karra, D.; Kiebler, M.A.; Macchi, P. Dendritic localization of the translational repressor pumilio 2 and its contribution to dendritic stress granules. J. Neurosci. 2006, 26, 6496–6508. [Google Scholar] [CrossRef] [PubMed]
- Cassola, A.; Romaniuk, M.A.; Primrose, D.; Cervini, G.; D’Orso, I.; Frasch, A.C. Association of UBP1 to ribonucleoprotein complexes is regulated by interaction with the trypanosome ortholog of the human multifunctional P32 protein. Mol. Microbiol. 2015, 97, 1079–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivas, W.; Parker, R. The Puf3 protein is a transcript-specific regulator of mRNA degradation in yeast. EMBO J. 2000, 19, 6602–6611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, M.W.; Stowell, J.A.; Passmore, L.A. RNA-binding proteins distinguish between similar sequence motifs to promote targeted deadenylation by Ccr4-Not. eLife 2019, 8, e40670. [Google Scholar] [CrossRef] [PubMed]
- Goldstrohm, A.C.; Hook, B.A.; Seay, D.J.; Wickens, M. PUF proteins bind Pop2p to regulate messenger RNAs. Nat. Struct. Mol. Biol. 2006, 13, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Qiang, W.; Zhu, M.; Ding, Y.; Shi, Q.; Chen, X.; Zsiros, E.; Wang, K.; Yang, X.; Kurita, T. Mammalian Pum1 and Pum2 control body size via translational regulation of the cell cycle inhibitor Cdkn1b. Cell Rep. 2019, 26, 2434. [Google Scholar] [CrossRef] [Green Version]
- Ariz, M.; Mainpal, R.; Subramaniam, K. C. elegans RNA-binding proteins PUF-8 and MEX-3 function redundantly to promote germline stem cell mitosis. Dev. Biol. 2009, 326, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Shigunov, P.; Sotelosilveira, J.R.; Kuligovski, C.; De Aguiar, A.M.; Rebelatto, C.L.K.; Moutinho, J.A.; Brofman, P.R.S.; Krieger, M.A.; Goldenberg, S.; Munroe, D.J. PUMILIO-2 is involved in the positive regulation of cellular proliferation in human adipose-derived Stem Cells. Stem Cells Dev. 2012, 21, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Chen, S.; Liu, Y.; Wang, L.L.; Zhao, Y.; Zong, Z.H. PUM1 promotes ovarian cancer proliferation, migration and invasion. Biochem. Biophys. Res. Commun. 2018, 497, 313–318. [Google Scholar] [CrossRef]
- Dai, H.; Shen, K.; Yang, Y.; Shuai, L.; Zheng, P.; Chen, Z.; Bie, P. PUM1 knockdown prevents tumor progression by activating the PERK/eIF2/ATF4 signaling pathway in pancreatic adenocarcinoma cells. Cell Death Dis. 2019, 10, 595. [Google Scholar] [CrossRef] [Green Version]
- Lindner, S.E.; Mikolajczak, S.A.; Vaughan, A.M.; Moon, W.; Joyce, B.R.; Sullivan, W.J., Jr.; Kappe, S.H. Perturbations of plasmodium Puf2 expression and RNA-seq of Puf2-deficient sporozoites reveal a critical role in maintaining RNA homeostasis and parasite transmissibility. Cell Microbiol. 2013, 15, 1266–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, J.; Li, J.; Fan, Q.; Li, X.; Li, X.; Cui, L. The Puf-family RNA-binding protein PfPuf2 regulates sexual development and sex differentiation in the malaria parasite Plasmodium falciparum. J. Cell Sci. 2010, 123, 1039–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ellenbecker, M.; Hickey, B.; Day, N.J.; Osterli, E.; Terzo, M.; Voronina, E. Antagonistic control of Caenorhabditis elegans germline stem cell proliferation and differentiation by PUF proteins FBF-1 and FBF-2. eLife 2020, 9, e52788. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Yang, C.; Liu, J.; Liu, Q. NcPuf1 Is a Key Virulence Factor in Neospora caninum. Pathogens 2020, 9, 1019. https://doi.org/10.3390/pathogens9121019
Wang C, Yang C, Liu J, Liu Q. NcPuf1 Is a Key Virulence Factor in Neospora caninum. Pathogens. 2020; 9(12):1019. https://doi.org/10.3390/pathogens9121019
Chicago/Turabian StyleWang, Chenrong, Congshan Yang, Jing Liu, and Qun Liu. 2020. "NcPuf1 Is a Key Virulence Factor in Neospora caninum" Pathogens 9, no. 12: 1019. https://doi.org/10.3390/pathogens9121019