The Surface Protein Fructose-1, 6 Bisphosphate Aldolase of Klebsiella pneumoniae Serotype K1: Role of Interaction with Neutrophils



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains
2.3. Isolation of Neutrophils
2.4. Opsonization and Infection of Neutrophils
2.5. Protein Extraction
2.6. Two-Dimensional Gel Electrophoresis and Image Analysis
2.7. Preparation of Recombinant Fructose-Bisphosphate Aldolase (FBA) Protein Andproduction of a Rabbit Antiserum against Purified Recombinant FBA
2.8. Sub-Cellular Localization of FBA
2.9. SDS-PAGE and Immunoblotting
2.10. Quantitative Reverse-Transcription Polymerase Chain-Reaction (qRT-PCR) Assay
2.11. Competition Inhibition Assays
2.12. Human Leukocyte Bactericidal Activity Assay after Block FBA of KP-M1 with Anti-FBA Antibody
2.13. Statistical Analyses
3. Result
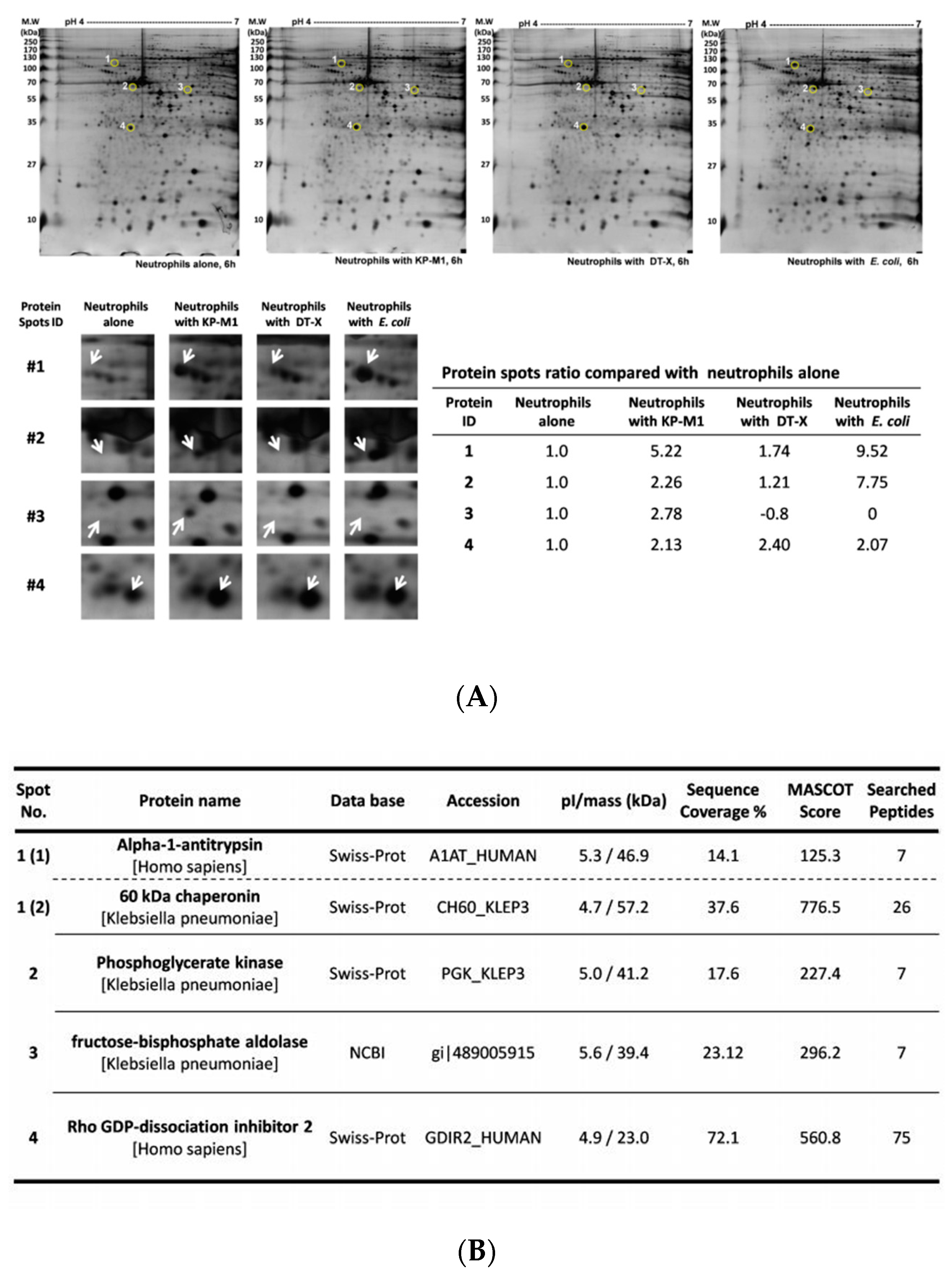
3.1. Characterization of Protein Profiles of Neutrophils Infected by Bacteria
3.2. KP-M1 FBA Is Localized to Cytoplasm and Outer Membrane
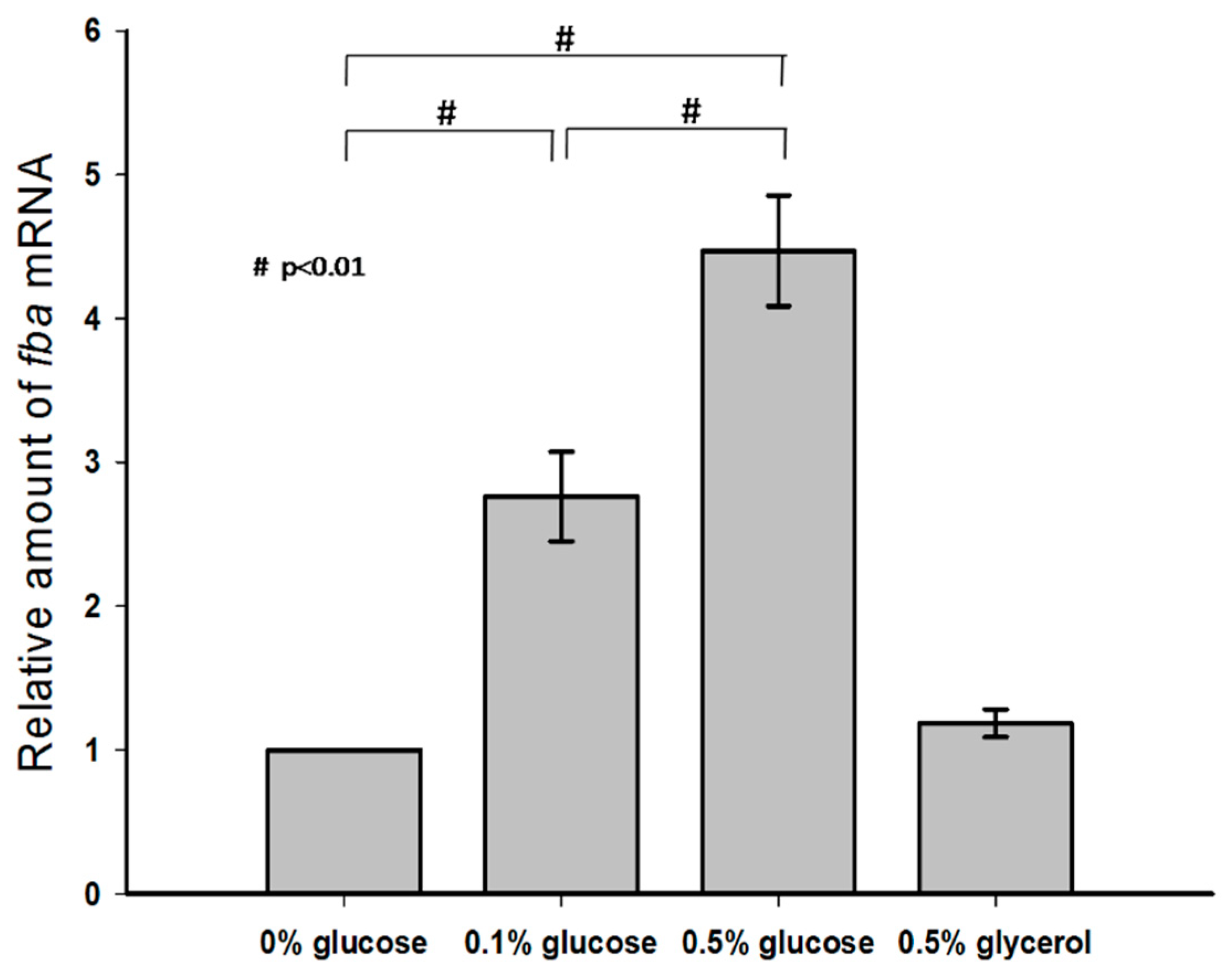
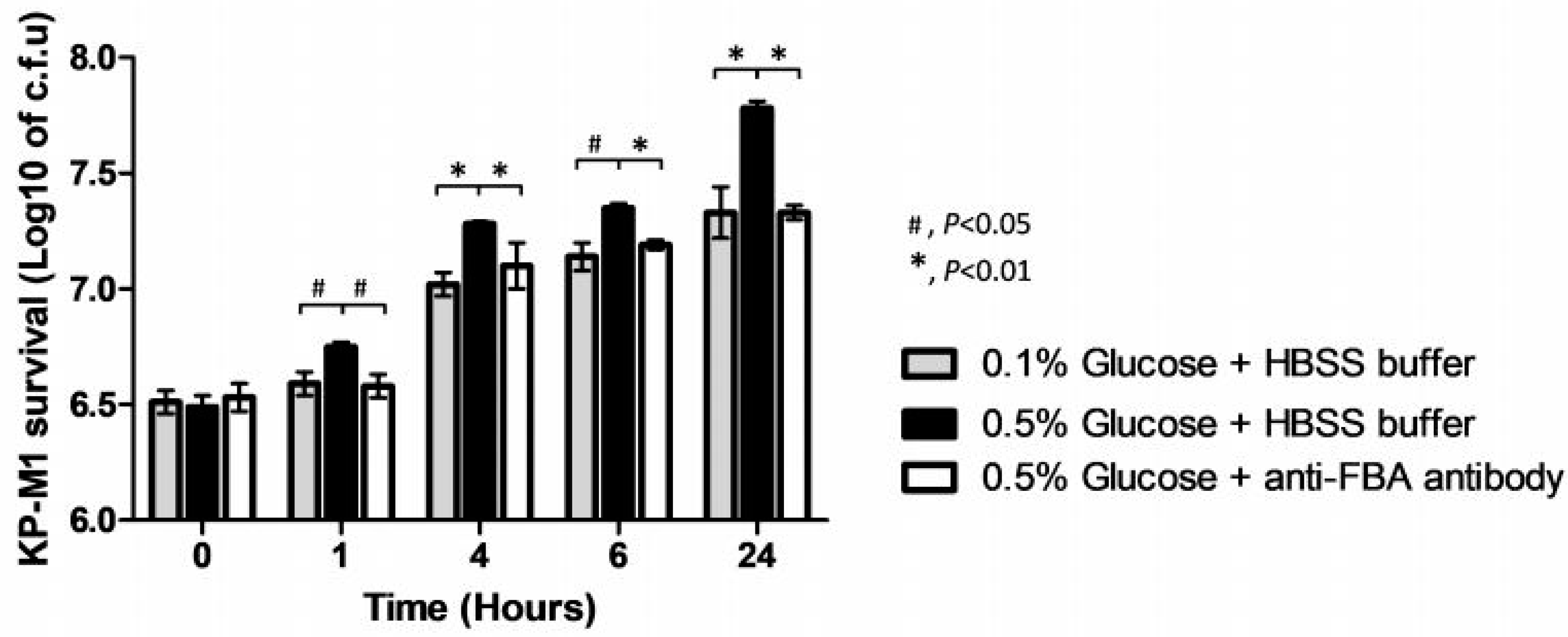
3.3. Effect of Exogenous Glucose on KP-M1 fba Transcription
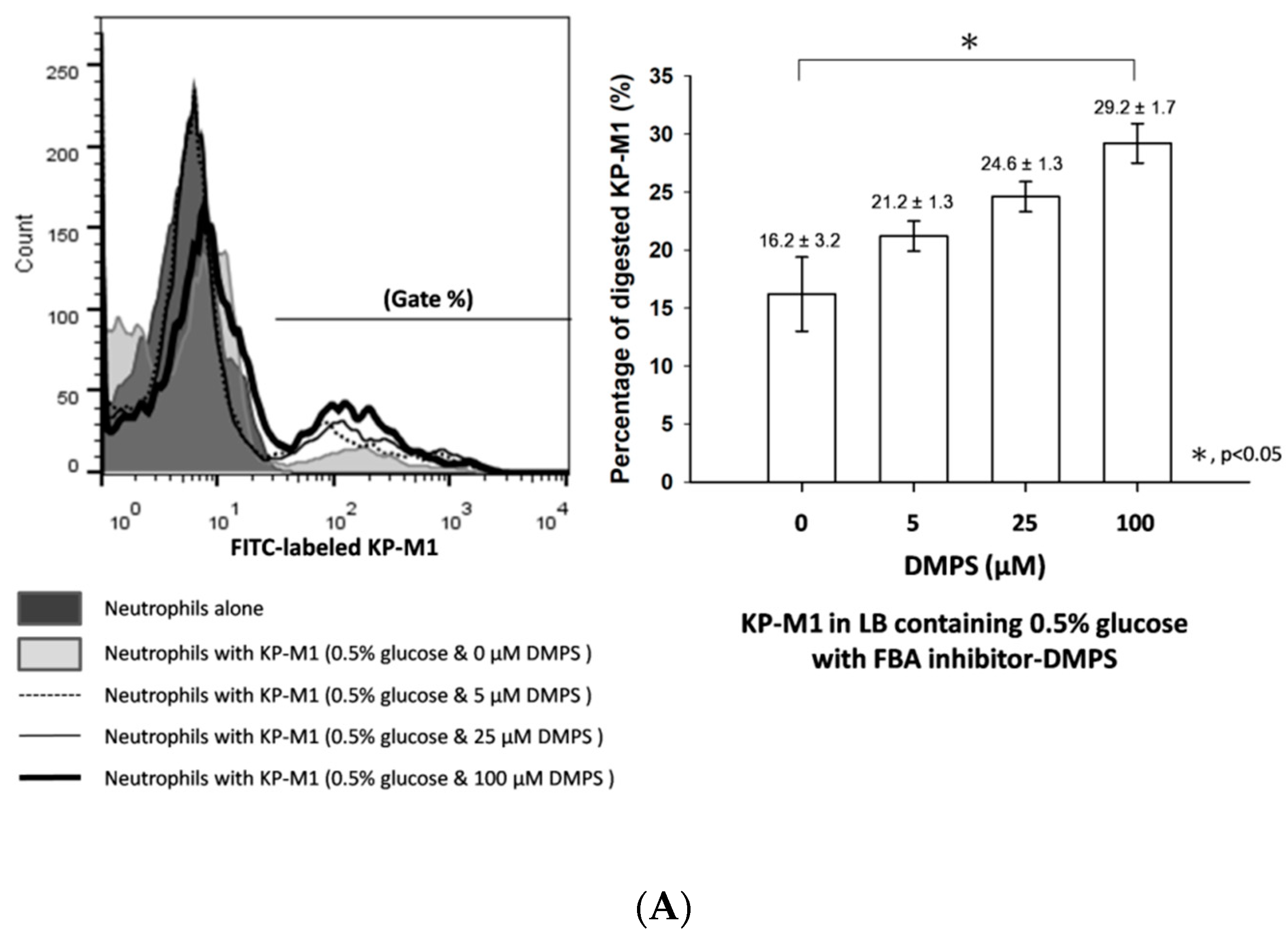
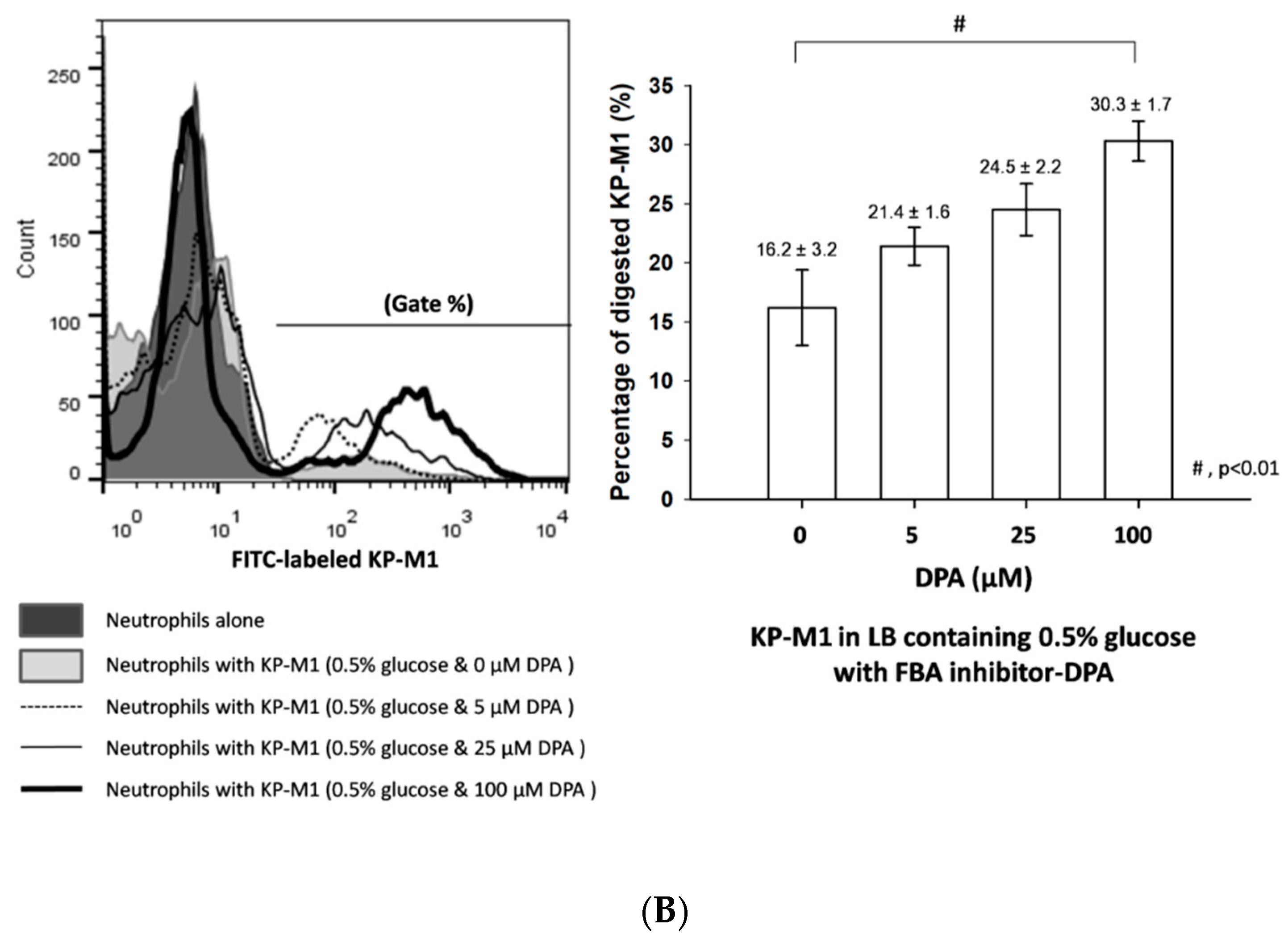
3.4. Phagocytosis of KP-M1 after CompetitionInhibition Assays
3.5. Anti-FBA Antibody Enhances Human Neutrophils Bactericidal Activity against KP-M1
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siu, L.K.; Yeh, K.M.; Lin, J.C.; Fung, C.P.; Chang, F.Y. Klebsiella pneumoniae liver abscess: A new invasive syndrome. Lancet Infect. Dis. 2012, 12, 881–887. [Google Scholar] [CrossRef]
- Chen, Y.C.; Lin, C.H.; Chang, S.N.; Shi, Z.Y. Epidemiology and clinical outcome of pyogenic liver abscess: An analysis from the National Health Insurance Research Database of Taiwan, 2000–2011. J. Microbiol. Immunol. Infect. 2016, 49, 646–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jepsen, P.; Vilstrup, H.; Schonheyder, H.C.; Sorensen, H.T. A nationwide study of the incidence and 30-day mortality rate of pyogenic liver abscess in Denmark, 1977–2002. Aliment. Pharmacol. Ther. 2005, 21, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- Meddings, L.; Myers, R.P.; Hubbard, J.; Shaheen, A.A.; Laupland, K.B.; Dixon, E.; Coffin, C.; Kaplan, G.G. A population-based study of pyogenic liver abscesses in the United States: Incidence, mortality, and temporal trends. Am. J. Gastroenterol. 2010, 105, 117–124. [Google Scholar] [CrossRef]
- Lin, J.C.; Siu, L.K.; Fung, C.P.; Tsou, H.H.; Wang, J.J.; Chen, C.T. Impaired phagocytosis of capsular serotype K1 or K2 Klebsiella pneumoniae in type 2 diabetic mellitus patients with poor glycemic control. J. Clin. Endocrinol. Metab. 2006, 91, 3084–3087. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Chen, I.L.; Chuah, S.K.; Tai, W.C.; Chang, C.C.; Chen, F.J.; Chen, J.F. Impact of glycemic control on capsular polysaccharide biosynthesis and opsonophagocytosis of Klebsiella pneumoniae: Implications for invasive syndrome in patients with diabetes mellitus. Virulence 2016, 7, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.L.; Yang, F.L.; Yang, A.S.; Peng, H.P.; Li, T.L.; Tsai, M.D.; Wu, S.H.; Wang, J.T. Amino acid substitutions of MagA in Klebsiella pneumoniae affect the biosynthesis of the capsular polysaccharide. PLoS ONE 2012, 7, e46783. [Google Scholar] [CrossRef] [Green Version]
- Fung, C.P.; Chang, F.Y.; Lin, J.C.; Ho, D.M.; Chen, C.T.; Chen, J.H.; Yeh, K.M.; Chen, T.L.; Lin, Y.T.; Siu, L.K. Immune response and pathophysiological features of Klebsiella pneumoniae liver abscesses in an animal model. Lab. Investig. 2011, 91, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Cortés, G.; Borrell, N.; de Astorza, B.; Gómez, C.; Sauleda, J.; Albertí, S. Molecular analysis of the contribution of the capsular polysaccharide and the lipopolysaccharide O side chain to the virulence of Klebsiella pneumoniae in a murine model of pneumonia. Infect. Immun. 2002, 70, 2583–2590. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Chang, C.C.; Liu, J.W.; Chen, R.F.; Yang, K.D. Sialic acid involved in hypermucoviscosity phenotype of Klebsiella pneumoniae and associated with resistance to neutrophil phagocytosis. Virulence 2014, 5, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Chuah, S.K.; Tai, W.C.; Chang, C.C.; Chen, F.J. Delay in human neutrophil constitutive apoptosis after infection with Klebsiella pneumoniae serotype K1. Front. Cell Infect. Microbiol. 2017, 7, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, J.J.; Kuo, H.C.; Lee, K.F.; Tsai, T.H. Proteomic analysis of plasma from rats following total parenteral nutrition-induced liver injury. Proteomics 2015, 15, 3865–3874. [Google Scholar] [CrossRef] [PubMed]
- Tunio, S.A.; Oldfield, N.J.; Berry, A.; Ala’Aldeen, D.A.; Wooldridge, K.G.; Turner, D.P. The moonlighting protein fructose-1, 6-bisphosphate aldolase of Neisseria meningitidis: Surface localization and role in host cell adhesion. Mol. Microbiol. 2010, 76, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziveri, J.; Tros, F.; Guerrera, I.C.; Chhuon, C.; Audry, M.; Dupuis, M.; Barel, M.; Korniotis, S.; Fillatreau, S.; Gales, L.; et al. The metabolic enzyme fructose-1,6-bisphosphate aldolase acts as a transcriptional regulator in pathogenic Francisella. Nat. Commun. 2017, 8, 853. [Google Scholar] [CrossRef] [PubMed]
- Labbé, G.; Krismanich, A.P.; de Groot, S.; Rasmusson, T.; Shang, M.; Brown, M.D.; Dmitrienko, G.I.; Guillemette, J.G. Development of metal-chelating inhibitors for the Class II fructose 1,6-bisphosphate (FBP) aldolase. J. Inorg. Biochem. 2012, 112, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Heinzelmann, M.; Gardner, S.A.; Mercer-Jones, M.; Roll, A.J.; Polk, H.C., Jr. Quantification of phagocytosis in human neutrophils by flow cytometry. Microbiol. Immunol. 1999, 43, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Hampton, M.B.; Vissers, M.C.; Winterbourn, C.C. A single assay for measuring the rates of phagocytosis and bacterial killing by neutrophils. J. Leukoc. Biol. 1994, 55, 147–152. [Google Scholar] [CrossRef]
- Green, J.N.; Winterbourn, C.C.; Hampton, M.B. Analysis of neutrophil bactericidal activity. Methods Mol. Biol. 2007, 412, 319–332. [Google Scholar]
- McDaniel, L.S.; Sheffield, J.S.; Delucchi, P.; Briles, D.E. PspA, a surface protein of Streptococcus pneumoniae, is capable of eliciting protection against pneumococci of more than one capsular type. Infect. Immun. 1991, 59, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.H.; Hsu, T.L.; Lin, S.Y.; Pan, Y.J.; Jan, J.T.; Wang, J.T.; Khoo, K.H.; Wu, S.H. Phosphoproteomics of Klebsiella pneumoniae NTUH-K2044 reveals a tight link between tyrosine phosphorylation and virulence. Mol. Cell Proteom. 2009, 8, 2613–2623. [Google Scholar] [CrossRef] [Green Version]
- Katebi, A.R.; Jernigan, R.L. Aldolases utilize different oligomeric states to preserve their functional dynamics. Biochemistry 2015, 54, 3543–3554. [Google Scholar] [CrossRef] [PubMed]
- Chhatwal, G.S. Anchorless adhesins and invasins of Gram-positive bacteria: A new class of virulence factors. Trends Microbiol. 2002, 10, 205–208. [Google Scholar] [CrossRef]
- Henderson, B.; Martin, A. Bacterial virulence in the moonlight: Multitasking bacterial moonlighting proteins are virulence determinants in infectious disease. Infect. Immun. 2011, 79, 3476–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Xia, Y.; Cui, J.; Gu, Z.; Song, Y.; Chen, Y.Q.; Chen, H.; Zhang, H.; Chen, W. The roles of moonlighting proteins in bacteria. Curr. Issues Mol. Biol. 2014, 16, 15–22. [Google Scholar]
- Shams, F.; Oldfield, N.J.; Wooldridge, K.G.; Turner, D.P. Fructose-1,6-bisphosphate aldolase (FBA)-a conserved glycolytic enzyme with virulence functions in bacteria: ‘ill met by moonlight’. Biochem. Soc. Trans. 2014, 42, 1792–1795. [Google Scholar] [CrossRef]
- Pancholi, V.; Chhatwal, G.S. Housekeeping enzymes as virulence factors for pathogens. Int. J. Med. Microbiol. 2003, 293, 391–401. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, W.; Lu, C. Immunoproteomic assay of surface proteins of Streptococcus suis serotype 9. FEMS Immunol. Med. Microbiol. 2008, 53, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Blau, K.; Portnoi, M.; Shagan, M.; Kaganovich, A.; Rom, S.; Kafka, D.; Chalifa Caspi, V.; Porgador, A.; Givon-Lavi, N.; Gershoni, J.M.; et al. Flamingo cadherin: A putative host receptor for Streptococcus pneumoniae. J. Infect. Dis. 2007, 195, 1828–1837. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhao, Y.; Liu, C.; Chen, Z.; Zhou, D. Molecular pathogenesis of Klebsiella pneumoniae. Future Microbiol. 2014, 9, 1071–1081. [Google Scholar] [CrossRef]
- Zhang, X.H.; Tee, L.Y.; Wang, X.G.; Huang, Q.S.; Yang, S.H. Off-target Effects in CRISPR/Cas9-mediated Genome Engineering. Mol. Ther. Nucleic Acids. 2015, 4, e264. [Google Scholar] [CrossRef]
- Mackow, N.A.; Shen, J.; Adnan, M.; Khan, A.S.; Fries, B.C.; Diago-Navarro, E. CRISPR-Cas influences the acquisition of antibiotic resistance in Klebsiella pneumoniae. PLoS ONE 2019, 14, e0225131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.-H.; Chuah, S.-K.; Chang, C.-C.; Chen, F.-J. The Surface Protein Fructose-1, 6 Bisphosphate Aldolase of Klebsiella pneumoniae Serotype K1: Role of Interaction with Neutrophils. Pathogens 2020, 9, 1009. https://doi.org/10.3390/pathogens9121009
Lee C-H, Chuah S-K, Chang C-C, Chen F-J. The Surface Protein Fructose-1, 6 Bisphosphate Aldolase of Klebsiella pneumoniae Serotype K1: Role of Interaction with Neutrophils. Pathogens. 2020; 9(12):1009. https://doi.org/10.3390/pathogens9121009
Chicago/Turabian StyleLee, Chen-Hsiang, Seng-Kee Chuah, Chia-Chi Chang, and Fang-Ju Chen. 2020. "The Surface Protein Fructose-1, 6 Bisphosphate Aldolase of Klebsiella pneumoniae Serotype K1: Role of Interaction with Neutrophils" Pathogens 9, no. 12: 1009. https://doi.org/10.3390/pathogens9121009