Renibacterium salmoninarum—The Causative Agent of Bacterial Kidney Disease in Salmonid Fish


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Search Strategy
3. Historical Background and Classification
4. Distribution of the Disease
4.1. Geographical and Temporal Distribution
4.2. Susceptible Species
5. Routes of Infection and Risk Factors of R. salmoninarum
6. Clinical Signs and Diagnosis
6.1. Clinical Signs
6.2. Histopathology
6.3. Cultivation and Biochemical Profile
6.4. Immunodiagnostic Methods
6.5. Molecular Diagnostic
7. Virulence Mechanisms
8. Treatment, Control and Disinfection Equipment of R. salmoninarum
8.1. Selection
8.2. Vaccination
8.3. Antibiotherapy
8.4. Nutritional Supplements
8.5. Culling and Segregation Infected Brood Stock
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
List of Abbreviations
References
- Bayliss, S.C.; Verner-Jeffreys, D.W.; Ryder, D.; Suarez, R.; Ramirez, R.; Romero, J.; Pascoe, B.; Sheppard, S.K.; Godoy, M.; Feil, E.J. Genomic epidemiology of the commercially important pathogen Renibacterium salmoninarum within the Chilean salmon industry. Microb. Genom. 2018, 4, 1–12. [Google Scholar] [CrossRef]
- Murray, A.G.; Munro, L.A.; Wallace, I.S.; Allan, C.E.T.; Peeler, E.J.; Thrush, M.A. Epidemiology of Renibacterium salmoninarum in Scotland and the potential for compartmentalised management of salmon and trout farming areas. Aquaculture 2012. [Google Scholar] [CrossRef]
- Balfry, S.K.; Albright, L.J.; Evelyn, T.P.T. Horizontal transfer of Renibacterium salmoninarum among farmed salmonids via the fecal-oral route. Dis. Aquat. Organ. 1996, 25, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Boerlage, A.S.; Elghafghuf, A.; Stryhn, H.; Sanchez, J.; Hammell, K.L. Risk factors associated with time to first clinical case of Bacterial Kidney Disease (BKD) in farmed Atlantic Salmon (Salmo salar L.) in New Brunswick, Canada. Prev. Vet. Med. 2018, 149, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Pascho, R.J.; Elliott, D.G.; Streufert, J.M. Brood stock segregation of spring chinook salmon Oncorhynchus tshawytscha by use of the enzyme-linked immunosorbent assay (ELISA) and the fluorescent antibody technique (FAT) affects the prevalence and levels of Renibacterium salmoninarum. Dis. Aquat. Organ. 1991, 12, 25–40. [Google Scholar] [CrossRef]
- Bruno, D.W. Prevalence and diagnosis of bacterial kidney disease (BKD) in Scotland between 1990 and 2002. Dis. Aquat. Organ. 2004, 59, 125–130. [Google Scholar] [CrossRef]
- Johansen, L.; Jensen, I.; Mikkelsen, H.; Bjørn, P.; Jansen, P.A.; Bergh, Ø. Disease interaction and pathogens exchange between wild and farmed fish populations with special reference to Norway. Aquaculture 2011, 315, 167–186. [Google Scholar] [CrossRef] [Green Version]
- Wiens, G.D. Bacterial Kidney Disease (Renibacterium salmoninarum), Fish Diseases and Disorders: Viral, Bacterial and Fungal Infections, 2nd ed.; Woo, P.T.K., Bruno, D.W., Eds.; CAB International: Wallingford, UK, 2011; pp. 338–374. [Google Scholar]
- Evenden, A.J.; Grayson, T.H.; Gilpin, M.L.; Munn, C.B. Renibacterium salmoninarum and bacterial kidney disease—The unfinished jigsaw. Annu. Rev. Fish Dis. 1993, 3, 87–104. [Google Scholar] [CrossRef]
- Boerlage, A.S.; Stryhn, H.; Sanchez, J.; Hammell, K.L. Case definition for clinical and subclinical bacterial kidney disease (BKD) in Atlantic Salmon (Salmo salar L.) in New Brunswick, Canada. J. Fish Dis. 2017, 40, 395–409. [Google Scholar] [CrossRef]
- Bruno, D.W. Scottish experience with bacterial kidney disease in farmed salmonids between 1976 and 1985. Aquac. Res. 1986, 17, 185–190. [Google Scholar] [CrossRef]
- Evelyn, T.P. Bacterial Kidney Disease-BKD, Bacterial Disease of Fish; Inglis, V., Roberts, R.J., Bromage, N.R., Eds.; Blackwell: Oxford, UK, 1993; pp. 177–195. [Google Scholar]
- Jónsdóttir, H.; Malmquist, H.J.; Snorrason, S.S.; Gudbergsson, G.; Gudmundsdottir, S. Epidemiology of Renibacterium salmoninarum in wild Arctic charr and brown trout in Iceland. J. Fish Biol. 1998, 53, 322–339. [Google Scholar] [CrossRef]
- Bruneau, N.; Thorburn, M. Occurrence of Aeromonas salmonicida, Renibacterium salmoninarum, and infectious pancreatic necrosis virus in Ontario salmonid populations. J. Aquat. Anim. Health. 1999, 11, 350–357. [Google Scholar] [CrossRef]
- Ahne, W.; Winton, J.R.; Kimura, T. Prevention of infectious diseases in aquaculture. J. Vet. Med. 1989, 36, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Misaka, N.; Mizuno, S.; Sasaki, Y. Subclinical infection of Renibacterium salmoninarum in fry and juveniles Chum salmon Oncorhynchus keta in Hokkaido, Japan. Fish Pathol. 2017, 52, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Belding, D.L.; Merrill, B. A preliminary report upon a hatchery disease of the Salmonidae. Trans. Am. Fish. Soc. 1935, 65, 76–84. [Google Scholar] [CrossRef]
- Fryer, J.L.; Sanders, J.E. Bacterial kidney disease of salmonid fish. Annu. Rev. Microbiol. 1981, 35, 273–298. [Google Scholar] [CrossRef]
- Matejusova, I.; Bain, N.; Colquhoun, D.J.; Feil, E.J.; Mccarthy, U.; Mclennan, D.; Snow, M.; Verner-jeffreys, D.; Wallace, I.S.; Weir, S.J.; et al. Multilocus variable-number tandem-repeat genotyping of Renibacterium salmoninarum, a bacterium causing bacterial kidney disease in salmonid fish. BMC Mic. 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Eissa, A.; Elsayed, E. An overview on bacterial kidney disease. Life Sci. J. 2006, 3, 58–76. [Google Scholar]
- Sanders, E.; Fryer, J.L. Causative agent of bacterial kidney disease in salmonid fishes. Int. J. Syst. Bacteriol. 1980, 30, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish, 6th ed.; Springer: Cham, Switzerland, 2016; pp. 84–106. [Google Scholar]
- Sanders, J.E.; Fryer, J.L. Corynebacterium salmoninus sp. nov. the causative agent of bacterial kidney disease; selected biochemical properties and pathogenesis in salmonid fishes. In Proceedings of the Joint 3rd Biennial Fish Health Section/American Fisheries Society and 9th Annual Midwest Fish Disease Workshops, Kansas City, MO, USA, 1978; pp. 28–33. [Google Scholar]
- Plumb, J.A. Health Maintenance of Cultured Fishes: Principal Microbial Diseases; Wiley-Blackwell: Hoboken, NJ, USA, 2018. [Google Scholar]
- Wiens, G.D.; Rockey, D.D.; Wu, Z.; Chang, J.; Levy, R.; Crane, S.; Chen, D.S.; Capri, G.R.; Burnett, J.R.; Sudheesh, P.S.; et al. Genome sequence of the fish pathogen Renibacterium salmoninarum suggests reductive evolution away from an environmental arthrobacter ancestor. J. Bacteriol. 2008, 190, 6970–6982. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.O. Molecular diagnosis of fish and shellfish diseases: Present status and potential use in disease control. Aquaculture 2002, 206, 19–55. [Google Scholar] [CrossRef]
- Brynildsrud, O.; Feil, E.J.; Bohlin, J.; Castillo-Ramirez, S.; Colquhoun, D.; McCarthy, U.; Matejusova, I.M.; Rhodes, L.D.; Wiens, G.D.; Verner-Jeffreys, D.W. Microevolution of Renibacterium salmoninarum: Evidence for intercontinental dissemination associated with fish movements. ISME J. 2014, 8, 746–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brynildsrud, O.; Gulla, S.; Feil, E.J.; Nørstebø, S.F.; Rhodes, L.D. Identifying copy number variation of the dominant virulence factors msa and p22 within genomes of the fish pathogen Renibacterium salmoninarum. Microb. Genom. 2016, 2, e000055. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.S.; Southgate, P.C.; Tucker, C.S. Aquaculture: Farming Aquatic Animals and Plants, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; p. 664. [Google Scholar]
- Purcell, M.K.; McKibben, C.L.; Pearman-Gillman, S.; Elliott, D.G.; Winton, J.R. Effects of temperature on Renibacterium salmoninarum infection and transmission potential in Chinook salmon, Oncorhynchus tshawytscha (Walbaum). J. Fish Dis. 2016, 39, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Pfeil-Putzien, C.; Hoffmann, R.; Popp, W. Preliminary report on the occurrence of bacterial kidney disease in Germany. Bull. Eur. Assoc. Fish Pathol. 1985, 5, 30–31. [Google Scholar]
- Rimaila-Pärnänen, E. First case of bacterial kidney disease (BKD) in whitefish (Coregonus lavaretus) in Finland. Bull. Eur. Assoc. Fish Pathol. 2002, 22, 403–404. [Google Scholar]
- Bell, G.R.; Hoffmann, R.W.; Brown, L.L. Pathology of experimental infections of the sablefish, Anoplopoma fimbria (Pallas), with Renibacterium salmoninarum, the agent of bacterial kidney disease in salmonids. J. Fish Dis. 1990, 13, 355–367. [Google Scholar] [CrossRef]
- Elliott, D.G. Renibacterium salmoninarum, Fish Viruses and Bacteria: Pathobiology and Protection; Woo, P.T.K., Cipriano, R.C., Eds.; CABI Publishing: Wallingford, UK, 2017; pp. 286–297. [Google Scholar]
- Nagai, T.; Iida, Y. Occurrence of bacterial kidney disease in cultured ayu Gyobyo Kenkyu. Fish Pathol. 2002, 37, 77–81. [Google Scholar] [CrossRef]
- Buller, N.B. Bacteria and Fungi from Fish and Other Aquatic Animals: A Practical Identification Manual, 2nd ed.; CABI Publishing: Wallingford, UK, 2014; p. 904. [Google Scholar]
- Paclibare, J.O.; Evelyn, T.P.T.; Albright, L.J.; Prosperi-Porta, L. Clearing of the kidney disease bacterium Renibacterium salmoninarum from seawater by the blue mussel Mytilus edulis, and the status of the mussel as a reservoir of the bacterium. Dis. Aquat. Organ. 1994, 18, 129–133. [Google Scholar] [CrossRef]
- Sakai, M.; Kobayashi, M. Detection of Renibacterium salmoninarum, the causative agent of bacterial kidney disease in salmonid fish, from pen-cultured coho salmon. Appl. Environ. Microbiol. 1992, 58, 1061–1063. [Google Scholar] [CrossRef] [Green Version]
- Chambers, E.; Gardiner, R.; Peeler, E.J. An investigation into the prevalence of Renibacterium salmoninarum in farmed rainbow trout, Oncorhynchus mykiss (Walbaum), and wild fish populations in selected river catchments in England and Wales between 1998 and 2000. J. Fish Dis. 2008, 31, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Wallace, I.S.; McKay, P.; Murray, A.G. A historical review of the key bacterial and viral pathogens of Scottish wild fish. J. Fish Dis. 2017, 40, 1741–1756. [Google Scholar] [CrossRef]
- Wallace, I.S.; Munro, L.A.; Kilburn, R.; Hall, M.; Black, J.; Raynard, R.S.; Murray, A.G. A Report on the Effectiveness of Cage and Farm-Level Fallowing for the Control of Bacterial Kidney Disease and Sleeping Disease on Large Cage-Based Trout Farms in Scotland. Available online: https://www2.gov.scot/Publications/2011/08/25130431/0 (accessed on 15 October 2020).
- Eissa, A.E.; Elsayed, E.E.; McDonald, R.; Faisal, M. First record of Renibacterium salmoninarum in the sea Lamprey (Petromyzon Marinus). J. Wildl. Dis. 2006, 42, 556–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisheries and Oceans Canada (DFO). Evaluation of Bacterial Kidney Disease (BKD) Impacts on the Canadian Salmon Aquaculture Industry. 2010. Available online: https://www.cahs-bc.ca/wp-content/uploads/2019/03/BKDWhitePaper_FinalApr2010_WithAppendices.pdf (accessed on 15 October 2020).
- Iwama, G.; Nakanishi, T. (Eds.) The Fish Immune System: Organism, Pathogen, and Environment; Academic Press: Cambridge, MA, USA, 1997; p. 312. [Google Scholar]
- Inglis, V.; Roberts, J.R.; Bromage, N.R. (Eds.) Bacterial Diseases of Fish; Blackwell Scientific Publication: Oxford, UK, 1993; p. 312. [Google Scholar]
- Mitchum, D.L.; Sherman, L.E. Transmission of bacterial kidney disease from wild to stocked hatchery trout. Can. J. Fish. Aquat. Sci. 1981, 38, 547–551. [Google Scholar] [CrossRef]
- Delghandi, M.R.; Menanteau-Ledouble, S.; Waldner, K.; El-Matbouli, M. Renibacterium salmoninarum and Mycobacterium spp.: Two bacterial pathogens present at low levels in wild brown trout (Salmo trutta fario) populations in Austrian rivers. BMC Vet. Res. 2020, 16, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Misaka, N.; Mizuno, S.; Sasaki, Y. PCR-based detection and quantification of Renibacterium salmoninarum in ovarian fluid of returning Chum salmon Oncorhynchus keta and Masu salmon O. masou in Hokkaido, Japan. Fish Pathol. 2017, 52, 100–103. [Google Scholar] [CrossRef] [Green Version]
- Evelyn, T.P.T.; Prosperi-Porta, L.; Ketcheson, J.E. Experimental intra-ovum infection of salmonid eggs with Renibacterium salmoninarum and vertical transmission of the pathogen with such eggs despite their treatment with erythromycin. Dis. Aquat. Organ. 1986, 1, 197–202. [Google Scholar] [CrossRef]
- Larson, D.L.; Faisal, M.; Tempelman, R.J.; Yu, H.; Scribner, K.T. Effects of hatchery rearing density, handling, and nutrition on Renibacterium salmoninarum infection prevalence in juvenile Chinook salmon (Oncorhynchus tshawytscha). J. Aquat. Anim. Health. 2020. [CrossRef]
- Yildiz, H.Y.; Seçer, F.S. Stress and Fish Health: Towards an Understanding of Allostatic Load. In Trends in Fisheries and Aquatic Animal Health; Berillis, P., Ed.; Bentham Science Publishers: Sharjah, UAE, 2017; pp. 133–154. [Google Scholar]
- Richards, C.A.; Murphy, C.A.; Brenden, T.O.; Loch, T.P.; Faisal, M. Detection accuracy of Renibacterium salmoninarum in Chinook salmon, Oncorhynchus tshawytscha (Walbaum) from non-lethally collected samples: Effects of exposure route and disease severity. Prev. Vet. Med. 2017, 145, 110–120. [Google Scholar] [CrossRef]
- Snieszko, S.F.; Griffin, P.J. Kidney disease in brook trout and its treatment. Progress. Fish-Cult. 1955, 17, 3–13. [Google Scholar] [CrossRef]
- Bell, G.R. Two epidemics of apparent kidney disease in cultured pink salmon (Oncorhynchus gorbuscha). J. Fish. Board Can. 1961, 18, 559–562. [Google Scholar] [CrossRef]
- Flaño, E.; López-Fierro, P.; Razquin, B.; Kaattari, S.L.; Villena, A. Histopathology of the renal and splenic haemopoietic tissues of coho salmon Oncorhynchus kisutch experimentally infected with Renibacterium salmoninarum. Dis. Aquat. Organ. 1996, 24, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Kent, M.L. Diseases of Seawater Netpen-Reared Salmonid Fishes in the Pacific Northwest; Canadian Special Publication of Fisheries and Aquatic Sciences; Department of Fisheries and Oceans: Nanaimo, BC, Canada, 1992; Volume 116, p. 76.
- Kent, M.L.; Benda, S.; St-hilaire, S.; Schreck, C.B. Sensitivity and specificity of histology for diagnoses of four common pathogens and detection of nontarget pathogens in adult Chinook salmon (Oncorhynchus tshawytscha) in fresh water. J. Vet. Diagnostic Investig. 2013, 25, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Speare, D.J. Differences in patterns of meningoencephalitis due to bacterial kidney disease in farmed Atlantic and Chinook salmon. Res. Vet. Sci. 1997, 62, 79–80. [Google Scholar] [CrossRef]
- Hirvelä-Koski, V.; Pohjanvirta, T.; Koski, P.; Sukura, A. Atypical growth of Renibacterium salmoninarum in subclinical infections. J. Fish Dis. 2006, 29, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Teska, J.D. In vitro growth of the bacterial kidney disease organism Renibacterium salmoninarum on a nonserum, noncharcoal-based “Homospecies-Metabolite” medium. J. Wildl. Dis. 1994, 30, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Chambers, E.; Barker, G. Comparison of culture media for the isolation of Renibacterium salmoninarum from naturally infected rainbow trout (Oncorhynchus mykiss). Bull. Assoc. FISH Pathol. 2006, 26, 137–142. [Google Scholar]
- Matsui, T.; Nishizawa, T.; Yoshimizu, M. Modification of KDM-2 with culture-spent medium for isolation of Renibacterium salmoninarum. Fish Pathol. 2009, 44, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Hirvelä-Koski, V. The fish pathogen Renibacterium salmoninarum: Growth in a microaerophilic atmosphere. Vet. Microbiol. 2008, 127, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Noga, E.J. Fish Disease: Diagnosis and Treatment, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; p. 519. [Google Scholar]
- World Organisation for Animal Health (OIE). Manual of Diagnostic Tests for Aquatic Animals, 4th ed.; The World Organisation of Animal Health (Office International des Epizooties): Paris, France, 2003; Volume 33, p. 355. [Google Scholar]
- Action, I.; Health, A.A.; Zrnčić, S.; Radosavljević, V. West Balkans Regional Aquatic Animal Disease Diagnostic Manual; Food and Agriculture Organization (FAO): Rome, Italy, 2017.
- Bethke, J.; Quezada, J.; Irgang, R.; Yáñez, A.; Oliver, C. Biochemical, serological, and genetic characterisation of Renibacterium salmoninarum isolates recovered from salmonids in Chile. Bull. Eur. Assoc. Fish Pathol. 2017, 37, 169–180. [Google Scholar]
- Brattgjerd, S.; Evensen, O.; Lauve, A. Effect of injected yeast glucan on the activity of macrophages in Atlantic salmon, Salmo salar L., as evaluated by in vitro hydrogen peroxide production and phagocytic capacity. Immunology 1994, 83, 288–294. [Google Scholar] [PubMed]
- Bullock, G.L.; Stuckey, H.M. Fluorescent antibody identification and detection of the Corynebacterium causing kidney disease of salmonids. J. Fish. Res. Board Can. 1975, 32, 2224–2227. [Google Scholar] [CrossRef]
- Elliott, D.G.; McKibben, C.L. Comparison of two fluorescent antibody techniques (FATS) for detection and quantification of Renibacterium salmoninarum in coelomic fluid of spawning chinook salmon Oncorhynchus tshawytscha. Dis. Aquat. Org. 1997, 30, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Sakai, M.; Amaaki, N.; Atsuta, S.; Kobayashi, M. Comparative sensitivities of several dot blot methods used to detect bacterial kidney diseases. J. Fish Dis. 1987, 10, 229–231. [Google Scholar] [CrossRef]
- Jansson, E.; Hongslo, T.; Höglund, J.; Ljungberg, O. Comparative evaluation of bacterial culture and two ELISA techniques for the detection of Renibacterium salmoninarum antigens in salmonid kidney tissues. Dis. Aquat. Org. 1996, 27, 197–206. [Google Scholar] [CrossRef]
- Meyers, T.R.; Short, S.; Farrington, C.; Lipson, K.; Geiger, H.J.; Gates, R. Comparison of the enzyme-linked immunosorbent assay (ELISA) and the fluorescent antibody test (FAT) for measuring the prevalences and levels of Renibacterium salmoninarum in wild and hatchery stocks of salmonid fishes in Alaska, USA. Dis. Aquat. Organ. 1993, 16, 181–189. [Google Scholar] [CrossRef]
- Arnason, I.Ö.; Sigurdardottir, S.; Kristmundsson, A.; Svansson, V.; Gudmundsdottir, S. Evaluation of a semi-nested PCR for detection of Renibacterium salmoninarum in samples from kidney, gill and ovarian fluid of Atlantic salmon broodfish. Icelandic Agric. Sci. 2013, 26, 49–57. [Google Scholar]
- Pascho, R.; Elliott, D.G.; Chase, D.M. Comparison of Traditional and Molecular Methods for Detection of Renibacterium salmoninarum.” Molecular Diagnosis of Salmonid Diseases; Cunningham, C.O., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 157–209. [Google Scholar]
- Jaramillo, D.; Gardner, I.A.; Stryhn, H.; Burnley, H.; Hammell, K.L. Bayesian latent class analysis of diagnostic sensitivity and specificity of tests for surveillance for bacterial kidney disease in Atlantic salmon Salmo salar. Aquaculture 2017, 476, 86–93. [Google Scholar] [CrossRef]
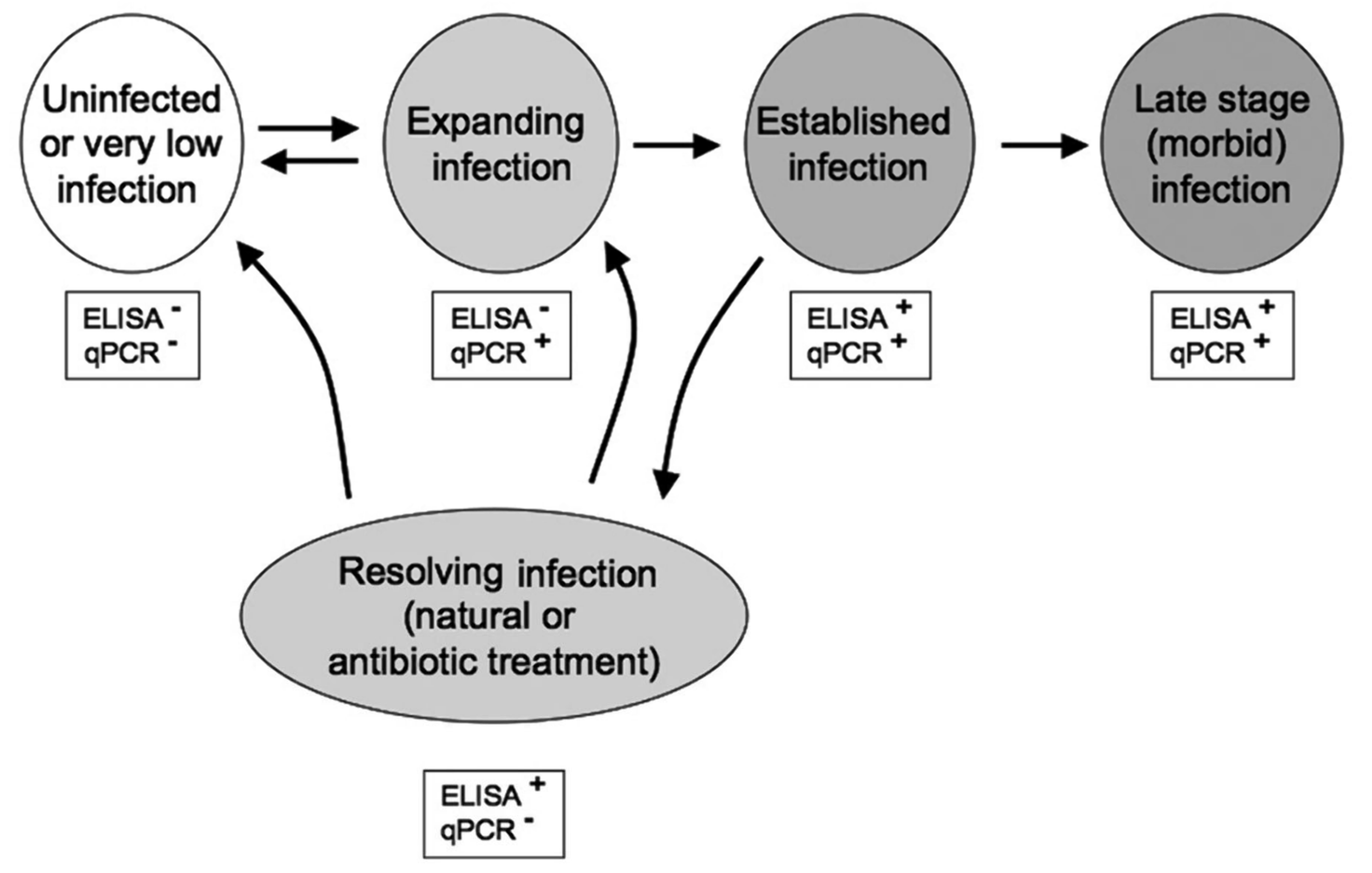
- Nance, S.L.; Riederer, M.; Zubkowski, T.; Trudel, M.; Rhodes, L.D. Interpreting dual ELISA and qPCR data for bacterial kidney disease of salmonids. Dis. Aquat. Organ. 2010, 91, 113–119. [Google Scholar] [CrossRef]
- Bruno, D.W.; Collet, B.; Turnbull, A.; Kilburn, R.; Walker, A.; Pendrey, D.; McIntosh, A.; Urquhart, K.; Taylor, G. Evaluation and development of diagnostic methods for Renibacterium salmoninarum causing bacterial kidney disease (BKD) in the UK. Aquaculture 2007, 269, 114–122. [Google Scholar] [CrossRef]
- Clarridge, J.E. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coady, A.M.; Murray, A.L.; Elliott, D.G.; Rhodes, L.D. Both msa genes in Renibacterium salmoninarum are needed for full virulence in bacterial kidney disease. Appl. Environ. Microbiol. 2006, 72, 2672–2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, D.G.; Applegate, L.J.; Murray, A.L.; Purcell, M.K.; Mckibben, C.L. Bench-top validation testing of selected immunological and molecular Renibacterium salmoninarum diagnostic assays by comparison with quantitative bacteriological culture. J. Fish Dis. 2013, 36, 779–809. [Google Scholar] [CrossRef]
- Cook, M.; Lynch, W.H. A sensitive nested reverse transcriptase PCR assay to detect viable cells of the fish pathogen Renibacterium salmoninarum in Atlantic salmon (Salmo salar L.). Appl. Environ. Microbiol. 1999, 65, 3042–3047. [Google Scholar] [CrossRef] [Green Version]
- Chase, D.M.; Elliott, D.G.; Pascho, R.J. Detection and quantification of Renibacterium salmoninarum DNA in salmonid tissues by real-time quantitative polymerase chain reaction analysis. J. Vet. Diagnostic Investig. 2006, 18, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Halaihel, N.; Vendrell, D.; Ruiz-Zarzuela, I.; de Blas, I.; Alonso, J.L.; Gironés, O.; Muzquiz, J.L. A new real time PCR-based assay for diagnosing Renibacterium salmoninarum in rainbow trout (Oncorhynchus mykiss) and comparison with other techniques. J. Microbiol. Methods. 2009, 76, 75–80. [Google Scholar] [CrossRef]
- Brown, L.L.; Iwama, G.K.; Evelyn, T.T.; Nelson, W.S.; Levine, R.P. Use of the polymerase chain reaction (PCR) to detect DNA from Renibacterium salmoninarum within individual salmonid eggs. Dis. Aquat. Organ. 1994, 18, 165–171. [Google Scholar] [CrossRef]
- McIntosh, D.; Meaden, P.G.; Austin, B. A simplified PCR-based method for the detection of Renibacterium salmoninarum utilizing preparations of rainbow trout (Oncorhynchus mykiss, Walbaum) lymphocytes. Appl. Environ. Microbiol. 1996, 62, 3929–3932. [Google Scholar] [CrossRef] [Green Version]
- Purcell, M.K.; Getchell, R.G.; McClure, C.A.; Garver, K.A. Quantitative polymerase chain reaction (PCR) for detection of aquatic animal pathogens in a diagnostic laboratory setting. J. Aquat. Anim. Health. 2011, 23, 148–161. [Google Scholar] [CrossRef]
- Sandell, T.A.; Jacobson, K.C. Comparison and evaluation of Renibacterium salmoninarum quantitative PCR diagnostic assays using field samples of Chinook and Coho salmon. Dis. Aquat. Organ. 2011, 93, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-rentería, M.; Rodríguez-gutiérrez, M.; Monroy-dosta, M.C.; Bustamante-, J.D. PCR optimization and evaluation for molecular detection of Renibacterium salmoninarum in rainbow trout (Oncorhynchus mykiss). Int. J. Aquat. Sci. 2019, 10, 3–10. [Google Scholar]
- Saleh, M.; Soliman, H.; El-Matbouli, M. Loop-mediated isothermal amplification (LAMP) for rapid detection of Renibacterium salmoninarum, the causative agent of bacterial kidney disease. Dis. Aquat. Organ. 2008, 81, 143–151. [Google Scholar] [CrossRef]
- Sudheesh, P.S.; Crane, S.; Cain, K.D.; Strom, M.S. Sortase inhibitor phenyl vinyl sulfone inhibits Renibacterium salmoninarum adherence and invasion of host cells. Dis. Aquat. Org. 2007, 78, 115–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grayson, T.H.; Cooper, L.F.; Wrathmell, A.B.; Roper, J.; Evenden, A.J.; Gilpin, M.L. Host responses to Renibacterium salmoninarum and specific components of the pathogen reveal the mechanisms of immune suppression and activation. Immunology 2002, 106, 273–283. [Google Scholar] [CrossRef]
- Piganelli, J.D.; Wiens, G.D.; Zhang, J.A.; Christensen, J.M.; Kaattari, S.L. Evaluation of a whole cell, p57- vaccine against Renibacterium salmoninarum. Dis. Aquat. Org. 1999, 36, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H. Intracellular bacterial infections: A challenge for developing cellular mediated immunity vaccines for farmed fish. Microorganisms 2018, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Wiens, G.D.; Pascho, R.; Winton, J.R. A single Ala139-to-Glu substitution in the Renibacterium salmoninarum virulence-associated protein p57 results in antigenic variation and is associated with enhanced p57 binding to Chinook salmon leukocytes. Appl. Environ. Microbiol. 2002, 68, 3969–3977. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.D.; Coady, A.M.; Deinhard, R.K. Identification of a third msa gene in Renibacterium salmoninarum and the associated virulence phenotype. Appl. Environ. Microbiol. 2004, 70, 6488–6494. [Google Scholar] [CrossRef] [Green Version]
- Wiens, G.D.; Dale, O.B. Renibacterium salmoninarum p57 antigenic variation is restricted in geographic distribution and correlated with genomic markers. Dis. Aquat. Organ. 2009, 83, 123–131. [Google Scholar] [CrossRef]
- Fredriksen, Å.; Endresen, C.; Wergeland, H.I. Immunosuppressive effect of a low molecular weight surface protein from Renibacterium salmoninarum on lymphocytes from Atlantic salmon (Salmo salar L.). Fish Shellfish Immunol. 1997, 7, 273–282. [Google Scholar] [CrossRef]
- Bethke, J.; Poblete-Morales, M.; Irgang, R.; Yáñez, A.; Avendaño-Herrera, R. Iron acquisition and siderophore production in the fish pathogen Renibacterium salmoninarum. J. Fish Dis. 2016, 39, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Grayson, T.; Bruno, D.W.; Evenden, A.; Gilpin, M.; Munn, C. Iron acquisition by Renibacterium salmoninarum: Contribution of iron reductase. Dis. Aquat. Organ. 1995, 22, 157–162. [Google Scholar] [CrossRef]
- Bethke, J.; Arias-Muñoz, E.; Yáñez, A.; Avendaño-Herrera, R. Renibacterium salmoninarum iron-acquisition mechanisms and ASK cell line infection: Virulence and immune response. J. Fish Dis. 2019, 1283–1291. [Google Scholar] [CrossRef]
- Bandin, I.; Santos, Y.; Bruno, D.W.; Raynard, R.S.; Toranzo, A.E.; Barja, J.L. Lack of biological activities in the extracellular products of Renibacterium salmoninarum. Can. J. Fish. Aquat. Sci. 1991, 48, 421–425. [Google Scholar] [CrossRef]
- Grayson, T.H.; Gilpin, M.L.; Evenden, A.J.; Munn, C.B. Evidence for the immune recognition of two haemolysins of Renibacterium salmoninarum by fish displaying clinical symptoms of bacterial kidney disease (BKD). Fish Shellfish Immunol. 2001, 11, 367–370. [Google Scholar] [CrossRef]
- Concha, M.I.; León, G. Cloning, functional expression and partial characterization of the glucose kinase from Renibacterium salmoninarum. FEMS Microbiol. Lett. 2000, 186, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Bethke, J.; Yáñez, A.J.; Avendaño-Herrera, R. Comparative genomic analysis of two Chilean Renibacterium salmoninarum isolates and the type strain ATCC 33209T. Genome Biol. Evol. 2018, 10, 1816–1822. [Google Scholar] [CrossRef] [Green Version]
- Kotob, M.H.; Menanteau-Ledouble, S.; Kumar, G.; Abdelzaher, M.; El-Matbouli, M. The impact of co-infections on fish: A review. Vet. Res. 2016, 47, 98. [Google Scholar] [CrossRef] [Green Version]
- Holborn, M.K.; Ang, K.P.; Elliott, J.A.K.; Powell, F.; Boulding, E.G. Genome wide association analysis for bacterial kidney disease resistance in a commercial North American Atlantic salmon (Salmo salar) population using a 50 K SNP panel. Aquaculture 2018, 495, 465–471. [Google Scholar] [CrossRef]
- Gjedrem, T.; Gjøen, H.M. Genetic variation in susceptibility of Atlantic salmon, Salmo salar L., to furunculosis, BKD and cold water vibriosis. Aquac. Res. 1995, 26, 129–134. [Google Scholar] [CrossRef]
- Eslamloo, K.; Kumar, S.; Caballero-Solares, A.; Gnanagobal, H.; Santander, J.; Rise, M.L. Profiling the transcriptome response of Atlantic salmon head kidney to formalin-killed Renibacterium salmoninarum. Fish Shellfish Immunol. 2020, 98, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.G.; Griffiths, S.G.; Kew, A.K.; Moore, A.R.; Olivier, G. Characterization of attenuated Renibacterium salmoninarum strains and their use as live vaccines. Dis. Aquat. Organ. 2001, 44, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salonius, K.; Siderakis, C.; MacKinnon, A.M.; Griffiths, S.G. Use of Arthrobacter davidanieli as a live vaccine against Renibacterium salmoninarum and Piscirickettsia salmonis in salmonids. Dev. Biol. 2005, 121, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Burnley, T.A.; Stryhn, H.; Burnley, H.J.; Hammell, K.L. Randomized clinical field trial of a bacterial kidney disease vaccine in Atlantic salmon, Salmo salar L. J. Fish Dis. 2010, 33, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Elliott, D.G.; Wiens, G.D.; Hammell, K.L.; Rhodes, L.D. Fish Vaccination “Vaccination against Bacterial Kidney Disease”; Gudding, R., Lillehaug, A., Evensen, O., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 255–272. [Google Scholar]
- Toranzo, A.E.; Romalde, J.L.; Magariños, B.; Barja, J.L. Present and future of aquaculture vaccines against fish bacterial diseases. In The Use of Veterinary Drugs and Vaccines in Mediterranean Aquaculture; Rogers, C., Basurco, B., Eds.; International Center for Advanced Mediterranean Agronomic Studies (CIHEAM): Zaragoza, Spain, 2009; pp. 155–176. [Google Scholar] [CrossRef]
- Griffiths, S.G.; Melville, K.J.; Salonius, K. Reduction of Renibacterium salmoninarum culture activity in Atlantic salmon following vaccination with avirulent strains. Fish Shellfish Immunol. 1998, 8, 607–619. [Google Scholar] [CrossRef]
- Alcorn, S.; Murray, A.L.; Pascho, R.J.; Varney, J. A cohabitation challenge to compare the efficacies of vaccines for bacterial kidney disease (BKD) in Chinook salmon Oncorhynchus tshawytscha. Dis. Aquat. Organ. 2005, 63, 151–160. [Google Scholar] [CrossRef]
- Rhodes, L.D.; Rathbone, C.K.; Corbett, S.C.; Harrell, L.W.; Strom, M.S. Efficacy of cellular vaccines and genetic adjuvants against bacterial kidney disease in Chinook salmon (Oncorhynchus tshawytscha). Fish Shellfish Immunol. 2004, 16, 461–474. [Google Scholar] [CrossRef]
- Rucker, R.R.; Bernier, A.F.; Whipple, W.J.; Burrows, R.E. Sulfadiazine for kidney disease. Progress. Fish Culturist. 1951, 13, 135–137. [Google Scholar] [CrossRef]
- Moffitt, C.M.; Bjornn, T.C. Protection of Chinook salmon smolts with oral doses of erythromycin against acute challenges of Renibacterium salmoninarum. J. Aquat. Anim. Health. 1989, 1, 227–232. [Google Scholar] [CrossRef]
- Fairgrieve, W.T.; Masada, C.L.; McAuley, W.C.; Peterson, M.E.; Myers, M.S.; Strom, M.S. Accumulation and clearance of orally administered erythromycin and its derivative, azithromycin, in juvenile fall Chinook salmon Oncorhynchus tshawytscha. Dis. Aquat. Organ. 2005, 64, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.M.; Wooster, G.A.; Bowser, P.R. Efficacy of enrofloxacin for the treatment of salmonids with bacterial kidney disease, caused by Renibacterium salmoninarum. J. Aquat. Anim. Health. 1994, 6, 220–223. [Google Scholar] [CrossRef]
- Moffitt, R. Incentive effects of the U.S. welfare system: A review. J. Econ. Lit. 1992, 30, 1–61. [Google Scholar]
- Rhodes, L.D.; Nguyen, O.T.; Deinhard, R.K.; White, T.M.; Harrell, L.W.; Roberts, M.C. Characterization of Renibacterium salmoninarum with reduced susceptibility to macrolide antibiotics by a standardized antibiotic susceptibility test. Dis. Aquat. Organ. 2008, 80, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, D.G.; Pascho, R.J.; Bullock, G.L. Developments in the control of bacterial kidney disease of salmonid fishes. Dis. Aquat. Org. 1989, 6, 201–215. [Google Scholar] [CrossRef]
- Keen, P.L.; Montforts, M.H.M.M. Antimicrobial Resistance in the Environment; Wiley-Blackwell: Hoboken, NJ, USA, 2012; p. 602. [Google Scholar]
- Woodall, A.N.; LaRoche, G. Nutrition of salmonid fishes: XI. iodide requirements of Chinook salmon. J. Nutr. 1964, 82, 475–482. [Google Scholar] [CrossRef]
- Bell, G.R.; Higgs, D.A.; Traxler, G.S. The effect of dietary ascorbate, zinc, and manganese on the development of experimentally induced bacterial kidney disease in sockeye salmon (Oncorhynchus nerka). Aquaculture 1984, 36, 293–311. [Google Scholar] [CrossRef]
- Thorarinsson, R.; Landolt, M.L.; Elliott, D.G.; Pascho, R.J.; Hardy, R.W. Effect of dietary vitamin E and selenium on growth, survival and the prevalence of Renibacterium salmoninarum infection in Chinook salmon (Oncorhynchus tshawytscha). Aquaculture 1994, 121, 343–358. [Google Scholar] [CrossRef]
- Sakai, D.K.; Nagat, M.; Iwami, T.; Koide, N.; Tamiya, Y.; Ito, Y.; Atoda, M. Attempt to control BKD by dietary modification and erythromycin chemotherapy in hatchery-reared masu salmon Oncorhynchus masou Brevoort. Bull. Jpn. Soc. Sci. Fish. 1986, 52, 1141–1147. [Google Scholar] [CrossRef]
- Armstrong, R.D.; Martin, S.W.; Evelyn, T.P.T.; Hicks, B.; Dorward, W.J.; Ferguson, H.W. A field evaluation of an indirect fluorescent antibody-based brood stock screening test used to control the vertical transmission of Renibacterium salmoninarum in Chinook salmon (Oncorhynchus tshawytscha). Can. J. Vet. Res. 1989, 53, 385–389. [Google Scholar]
- Elliott, D.G.; Mckibben, C.L.; Conway, C.M.; Purcell, M.K.; Chase, D.M.; Applegate, L.J. Testing of candidate non-lethal sampling methods for detection of Renibacterium salmoninarum in juvenile Chinook salmon Oncorhynchus tshawytscha. Dis. Aquat. Organ. 2015, 114, 21–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu∂mundsdóttir, S.; Helgason, S.; Sigurjónsdóttir, H.; Matthıasdóttir, S.; Jónsdóttir, H.; Laxdal, B.; Benediktsdóttir, E. Measures applied to control Renibacterium salmoninarum infection in Atlantic salmon: A retrospective study of two sea ranches in Iceland. Aquaculture 2000, 186, 193–203. [Google Scholar] [CrossRef]
- Laurin, E.; Morrison, D.; Gardner, I.A.; Siah, A.; Powell, J.F.F.; Kamaitis, M. Bayesian latent class analysis of ELISA and RT-rPCR diagnostic accuracy for subclinical Renibacterium salmoninarum infection in Atlantic salmon (Salmo salar) broodstock. J. Fish Dis. 2019, 42. [Google Scholar] [CrossRef]
- Murray, A.G.; Hall, M.; Munro, L.A.; Wallace, I.S. Modelling management strategies for a disease including undetected sub-clinical infection: Bacterial kidney disease in Scottish salmon and trout farms. Epidemic 2011, 3, 171–182. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delghandi, M.R.; El-Matbouli, M.; Menanteau-Ledouble, S. Renibacterium salmoninarum—The Causative Agent of Bacterial Kidney Disease in Salmonid Fish. Pathogens 2020, 9, 845. https://doi.org/10.3390/pathogens9100845
Delghandi MR, El-Matbouli M, Menanteau-Ledouble S. Renibacterium salmoninarum—The Causative Agent of Bacterial Kidney Disease in Salmonid Fish. Pathogens. 2020; 9(10):845. https://doi.org/10.3390/pathogens9100845
Chicago/Turabian StyleDelghandi, Mohammad Reza, Mansour El-Matbouli, and Simon Menanteau-Ledouble. 2020. "Renibacterium salmoninarum—The Causative Agent of Bacterial Kidney Disease in Salmonid Fish" Pathogens 9, no. 10: 845. https://doi.org/10.3390/pathogens9100845