Immune Modulation and the Development of Fowl Typhoid: A Model of Human Disease?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. S. enterica Strains
2.2. In Vitro Survival of S. enterica in Isolated chMDMs
2.3. Isolation of Chicken CD4+ T Cells and Avian Macrophage/CD4+ T Cell Model In Vitro
2.4. Phenotypic Analysis of Infected chMDMs and CD4+T Cells from Co-Culture with chMDMs
2.5. Salmonella Infection In Vivo
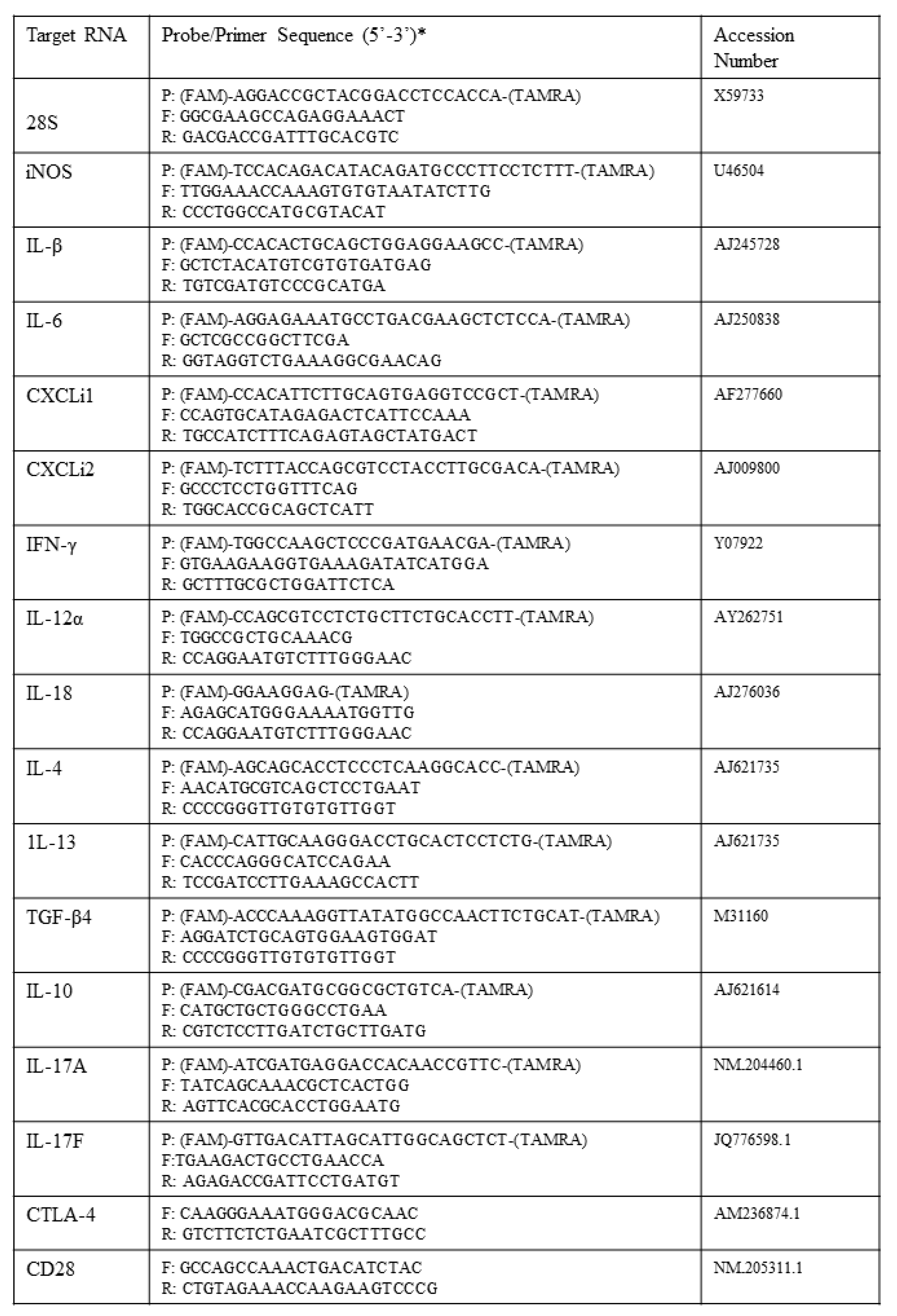
2.6. Gene Expression Analysis by qRT-PCR
2.7. Statistical Analysis
3. Results
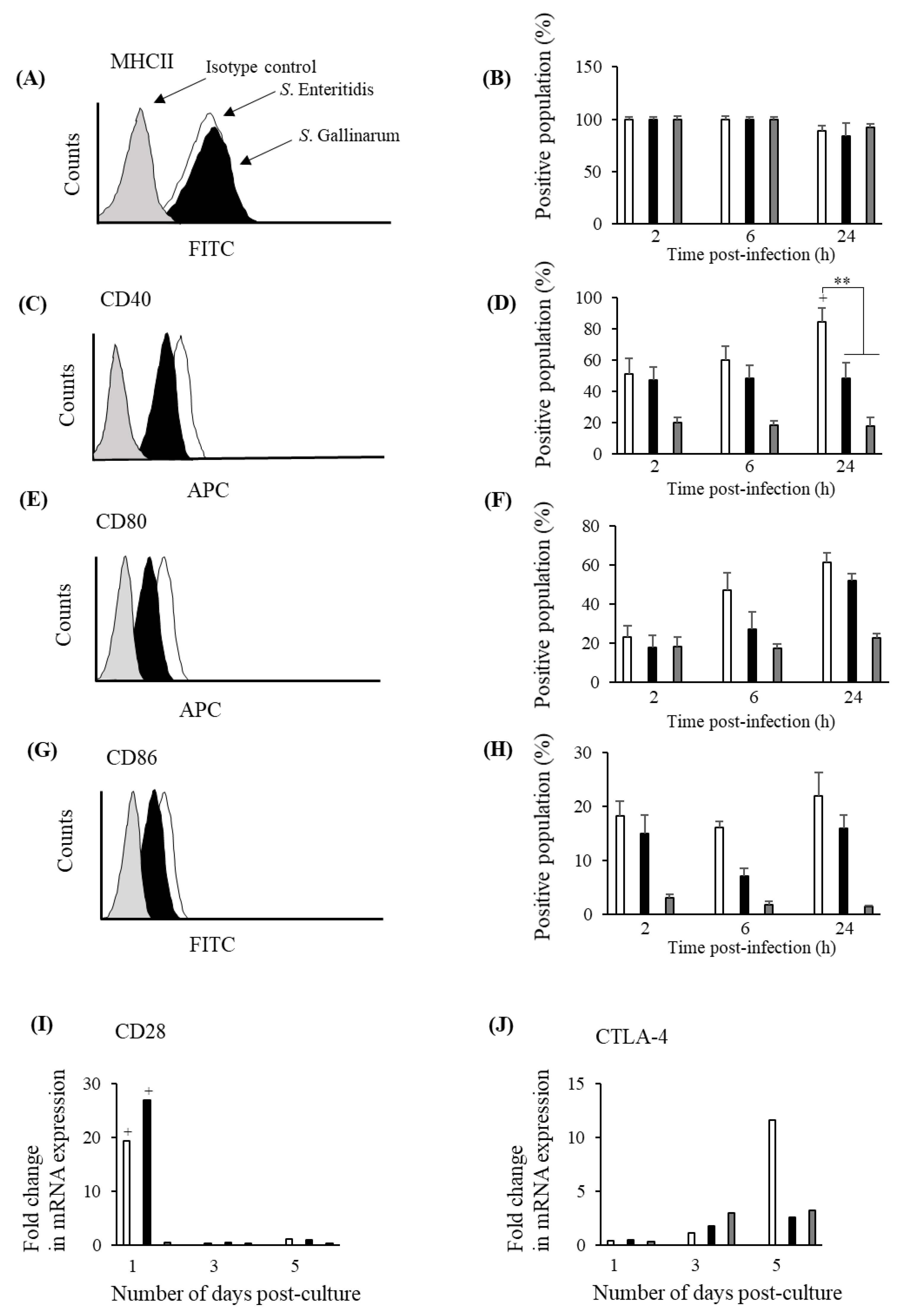
3.1. S. Gallinarum and S. Enteritidis Show Differential Infection and Survival Patterns in chMDMs
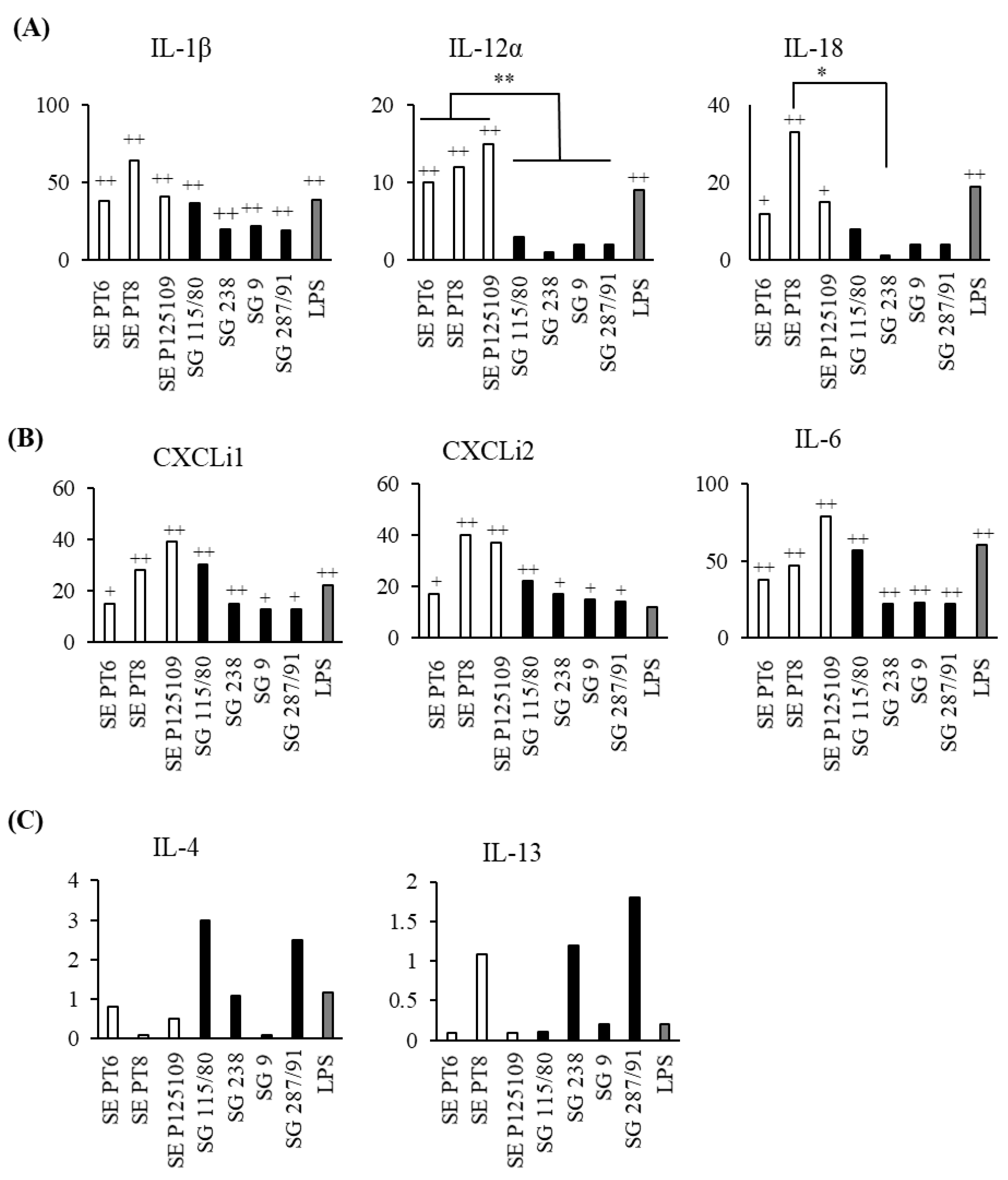
3.2. S. Gallinarum Induces Lower Expression of Inflammatory Cytokines and Chemokines by chMDMs Compared to S. Enteritidis
3.3. Different Strains of S. Gallinarum and S. Enteritidis Induced Similar Cytokine mRNA Expression Profiles in chMDMs
3.4. S. Enteritidis Induces a Dominant Th17/IFN-γ Response in CD4+ Lymphocytes Compared to S. Gallinarum
3.5. S. Gallinarum Stimulates Lower Levels of Co-Stimulatory Molecules on the Surface of chMDMs Compared to S. Enteritidis but a Similar Expression Profile for CD28 on CD4+ Lymphocytes
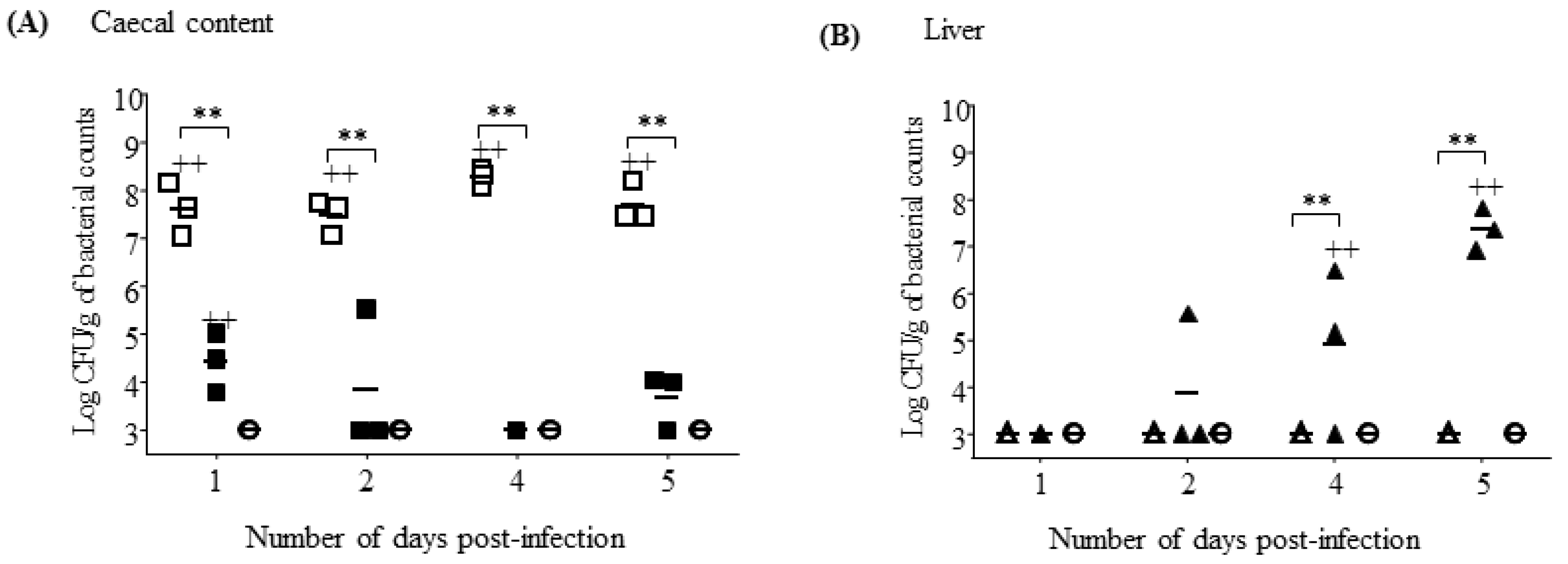
3.6. S. Gallinarum Effectively Disseminates from the Gastro-Intestinal Tract to Colonise Deeper Tissues
3.7. S. Gallinarum Suppresses the Chemokine Response in the Caecal Tonsil In Vivo
3.8. S. Gallinarum Suppresses Expression of Inflammatory Mediators in the Caecal Tonsil and Spleen and Promotes an Anti-Inflammatory IL-10 Response in the Spleen
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Hornick, R.B.; Greisman, S.E.; Woodward, T.E.; Dupont, H.L.; Dawkins, A.T.; Snyder, M.J. Typhoid Fever: Pathogenesis and Immunologic Control. N. Engl. J. Med. 1970, 283, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P.; Rothwell, L.; Galyov, E.E.; Barrow, P.A.; Burnside, J.; Wigley, P. Differential cytokine expression in avian cells in response to invasion by Salmonella typhimurium, Salmonella enteritidis and Salmonella gallinarum. Microbiology 2000, 146, 3217–3226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, L.; Kaiser, P.; Barrow, P.; Jones, M.A.; Johnston, C.; Wigley, P. The immunobiology of avian systemic salmonellosis. Veter- Immunol. Immunopathol. 2009, 128, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Langridge, G.C.; Fookes, M.; Connor, T.R.; Feltwell, T.; Feasey, N.; Parsons, B.N.; Seth-Smith, H.M.B.; Barquist, L.; Stedman, A.; Humphrey, T.; et al. Patterns of genome evolution that have accompanied host adaptation inSalmonella. Proc. Natl. Acad. Sci. USA 2014, 112, 863–868. [Google Scholar] [CrossRef] [Green Version]
- Crump, J.A.; Mintz, E.D. Global Trends in Typhoid and Paratyphoid Fever. Clin. Infect. Dis. 2010, 50, 241–246. [Google Scholar] [CrossRef]
- Mogasale, V.; Maskery, B.; Ochiai, R.L.; Lee, M.J.S.; Mogasale, V.V.; Ramani, M.E.; Kim, Y.-E.; Park, J.K.; Wierzba, T.F. Burden of typhoid fever in low-income and middle-income countries: A systematic, literature-based update with risk-factor adjustment. Lancet Glob. Heal. 2014, 2, e570–e580. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.M.; O Tacket, C.; Sztein, M.B. Host–Salmonella interaction: Human trials. Microbes Infect. 2001, 3, 1271–1279. [Google Scholar] [CrossRef]
- Salerno-Gonçalves, R.; Fernandez-Viña, M.; Lewinsohn, D.M.; Sztein, M.B. Identification of a Human HLA-E-Restricted CD8+ T Cell Subset in Volunteers Immunized with Salmonella enterica Serovar Typhi Strain Ty21a Typhoid Vaccine. J. Immunol. 2004, 173, 5852–5862. [Google Scholar] [CrossRef] [Green Version]
- Sztein, M.B. Cell-Mediated Immunity and Antibody Responses Elicited by Attenuated Salmonella enterica Serovar Typhi Strains Used as Live Oral Vaccines in Humans. Clin. Infect. Dis. 2007, 45, S15–S19. [Google Scholar] [CrossRef] [Green Version]
- Bhuiyan, S.; Sayeed, A.; Khanam, F.; Leung, D.T.; Bhuiyan, T.R.; Sheikh, A.; Salma, U.; Larocque, R.C.; Harris, J.B.; Pacek, M.; et al. Cellular and cytokine responses to Salmonella enterica serotype Typhi proteins in patients with typhoid fever in Bangladesh. Am. J. Trop. Med. Hyg. 2014, 90, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- Looney, R.J.; Steigbigel, R.T. Role of the Vi antigen of Salmonella typhi in resistance to host defense in vitro. J. Lab. Clin. Med. 1986, 108, 498–505. [Google Scholar]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Dusold, R.; Rubino, S.; Bäumler, A.J. The Vi Capsular Antigen of Salmonella enterica Serotype Typhi Reduces Toll-Like Receptor-Dependent Interleukin-8 Expression in the Intestinal Mucosa. Infect. Immun. 2005, 73, 3367–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, A.M.; Hall, L.J.; Clare, S.; Goulding, D.; Holt, K.E.; Grant, A.J.; Mastroeni, P.; Dougan, G.; Kingsley, R.A. A Salmonella Typhimurium-Typhi Genomic Chimera: A Model to Study Vi Polysaccharide Capsule Function In Vivo. PLoS Pathog. 2011, 7, e1002131. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A. Experimental infection of chickens withSalmonella enteritidis. Avian Pathol. 1991, 20, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrow, P.A.; Lovell, M.A.; Berchieri, A. The use of two live attenuated vaccines to immunize egg-laying hens againstSalmonella enteritidisphage type 4. Avian Pathol. 1991, 20, 681–692. [Google Scholar] [CrossRef]
- Smith, H.W. Observations on Experimental Fowl Typhoid. J. Comp. Pathol. Ther. 1955, 65, 37–IN5. [Google Scholar] [CrossRef]
- Ewigley, P.; Hulme, S.D.; Bumstead, N.; Barrow, P.A. In vivo and in vitro studies of genetic resistance to systemic salmonellosis in the chicken encoded by the SAL1 locus. Microbes Infect. 2002, 4, 1111–1120. [Google Scholar] [CrossRef]
- Tang, Y.; Foster, N.; Jones, M.A.; Barrow, P. Model of Persistent Salmonella Infection: Salmonella enterica Serovar Pullorum Modulates the Immune Response of the Chicken from a Th17-Type Response towards a Th2-Type Response. Infect. Immun. 2018, 86, 00307-18. [Google Scholar] [CrossRef] [Green Version]
- Hughes, S.; Poh, T.-Y.; Bumstead, N.; Kaiser, P. Re-evaluation of the chicken MIP family of chemokines and their receptors suggests that CCL5 is the prototypic MIP family chemokine, and that different species have developed different repertoires of both the CC chemokines and their receptors. Dev. Comp. Immunol. 2007, 31, 72–86. [Google Scholar] [CrossRef]
- Jones, M.A.; Wigley, P.; Page, K.L.; Hulme, S.D.; Barrow, P.A. Salmonella enterica Serovar Gallinarum Requires the Salmonella Pathogenicity Island 2 Type III Secretion System but Not the Salmonella Pathogenicity Island 1 Type III Secretion System for Virulence in Chickens. Infect. Immun. 2001, 69, 5471–5476. [Google Scholar] [CrossRef] [Green Version]
- Bravo-Blas, A.; Utriainen, L.; Clay, S.L.; Kästele, V.; Cerovic, V.; Cunningham, A.F.; Henderson, I.R.; Wall, D.M.; Milling, S. Salmonella enterica Serovar Typhimurium Travels to Mesenteric Lymph Nodes Both with Host Cells and Autonomously. J. Immunol. 2019, 202, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Mastroeni, P.; Vazquez-Torres, A.; Fang, F.C.; Xu, Y.; Khan, S.; Hormaeche, C.E.; Dougan, G. Antimicrobial actions of the NADPH phagocyte oxidase and inducible nitric oxide synthase in experimental salmonellosis. II. Effects on microbial proliferation and host survival in vivo. J. Exp. Med. 2000, 192, 237–248. [Google Scholar] [CrossRef]
- Hulme, S.D.; Barrow, P.A.; Foster, N. Inhibited Production of iNOS by Murine J774 Macrophages Occurs via a phoP-Regulated Differential Expression of NFκB and AP-1. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 483170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, N.; Lovell, M.A.; Marston, K.L.; Hulme, S.D.; Frost, A.J.; Bland, P.; Barrow, P. Rapid Protection of Gnotobiotic Pigs against Experimental Salmonellosis following Induction of Polymorphonuclear Leukocytes by Avirulent Salmonella enterica. Infect. Immun. 2003, 71, 2182–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaggerty, C.L.; Pevzner, I.Y.; Kogut, M.H. Selection for pro-inflammatory mediators yields chickens with increased resistance against Salmonella enterica serovar Enteritidis. Poult. Sci. 2014, 93, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J. Host-Microbe Interactions and Gut Health in Poultry-Focus on Innate Responses. Microorganisms 2019, 7, 139. [Google Scholar] [CrossRef] [Green Version]
- Berchieri, A.; Murphy, C.K.; Marston, K.; Barrow, P.A. Observations on the persistence and vertical transmission of Salmonella enterica serovars Pullorum and Gallinarum in chickens: Effect of bacterial and host genetic background. Avian Pathol. 2001, 30, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Ewigley, P.; Berchieri, A.; Page, K.L.; Smith, A.L.; Barrow, P.A. Salmonella enterica Serovar Pullorum Persists in Splenic Macrophages and in the Reproductive Tract during Persistent, Disease-Free Carriage in Chickens. Infect. Immun. 2001, 69, 7873–7879. [Google Scholar] [CrossRef] [Green Version]
- Barrow, P.A.; Lovell, M.A. Experimental infection of egg-laying hens with Salmonella enteritidis phage type 4. Avian Pathol. 1991, 20, 335–348. [Google Scholar] [CrossRef] [Green Version]
- Keuter, M.; Dharmana, E.; Gasem, M.H.; Van Der Ven-Jongekrijg, J.; Djokomoeljanto, R.; Dolmans, W.M.V.; Demacker, P.; Sauerwein, R.; Gallati, H.; Van Der Meer, J.W.M. Patterns of Proinflammatory Cytokines and Inhibitors during Typhoid Fever. J. Infect. Dis. 1994, 169, 1306–1311. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Jones, M.; Barrow, P.A.; Foster, N. Immune Modulation and the Development of Fowl Typhoid: A Model of Human Disease? Pathogens 2020, 9, 843. https://doi.org/10.3390/pathogens9100843
Tang Y, Jones M, Barrow PA, Foster N. Immune Modulation and the Development of Fowl Typhoid: A Model of Human Disease? Pathogens. 2020; 9(10):843. https://doi.org/10.3390/pathogens9100843
Chicago/Turabian StyleTang, Ying, Michael Jones, Paul A. Barrow, and Neil Foster. 2020. "Immune Modulation and the Development of Fowl Typhoid: A Model of Human Disease?" Pathogens 9, no. 10: 843. https://doi.org/10.3390/pathogens9100843