In Vivo Antiviral Effects of U18666A Against Type I Feline Infectious Peritonitis Virus


Abstract
:1. Introduction
2. Results
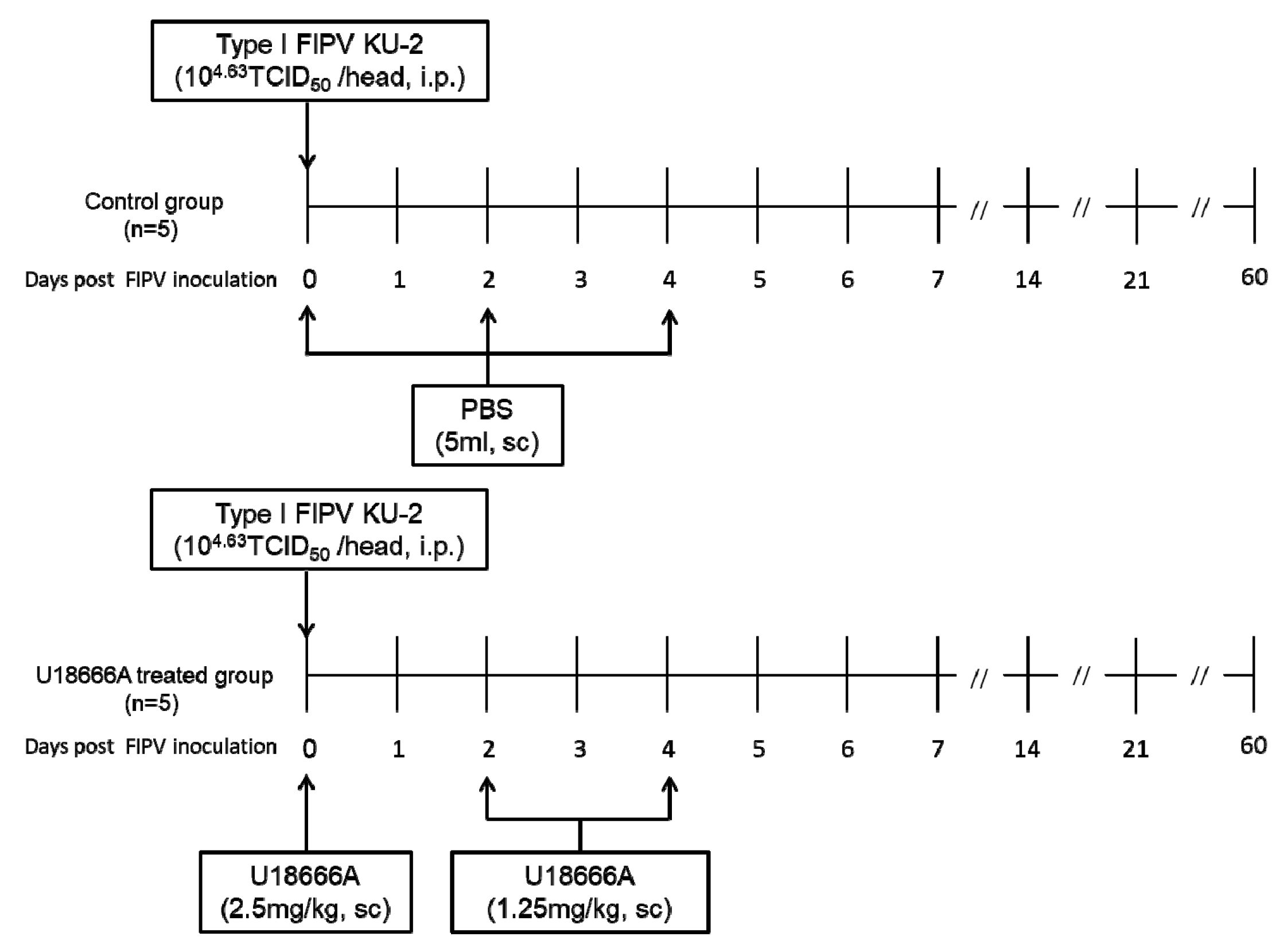
2.1. Experimental Schedule
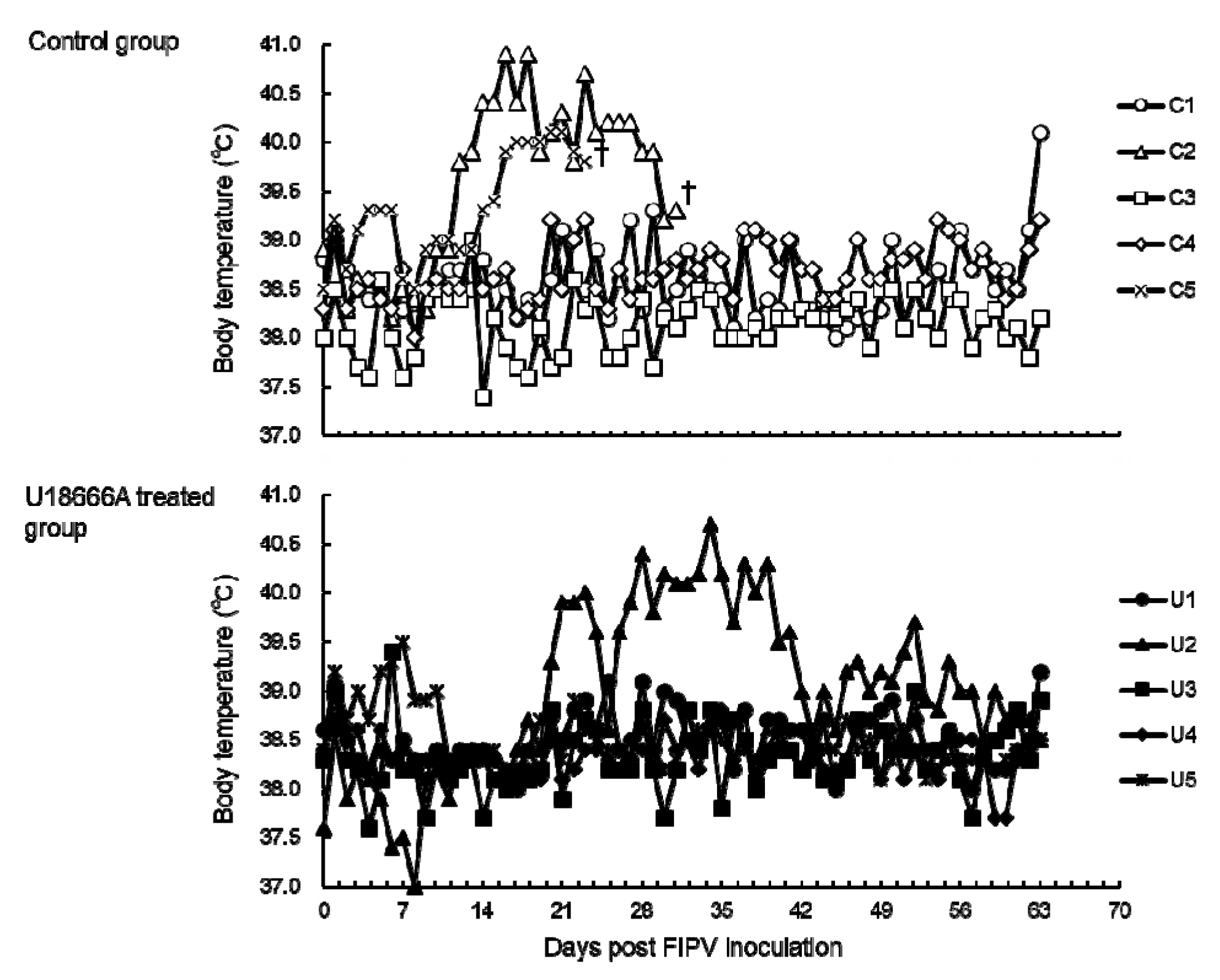
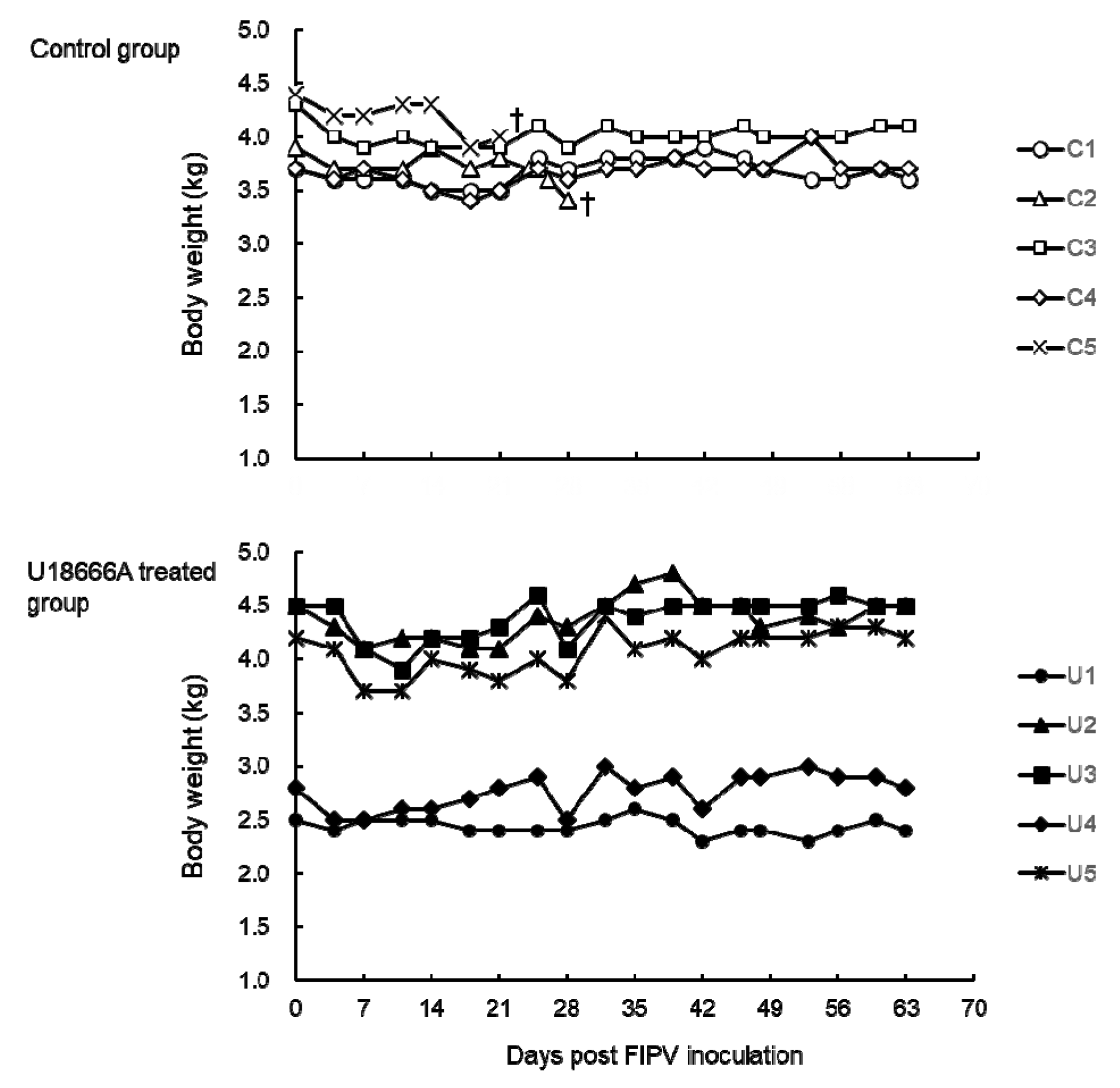
2.2. Changes in Body Temperature and Body Weight
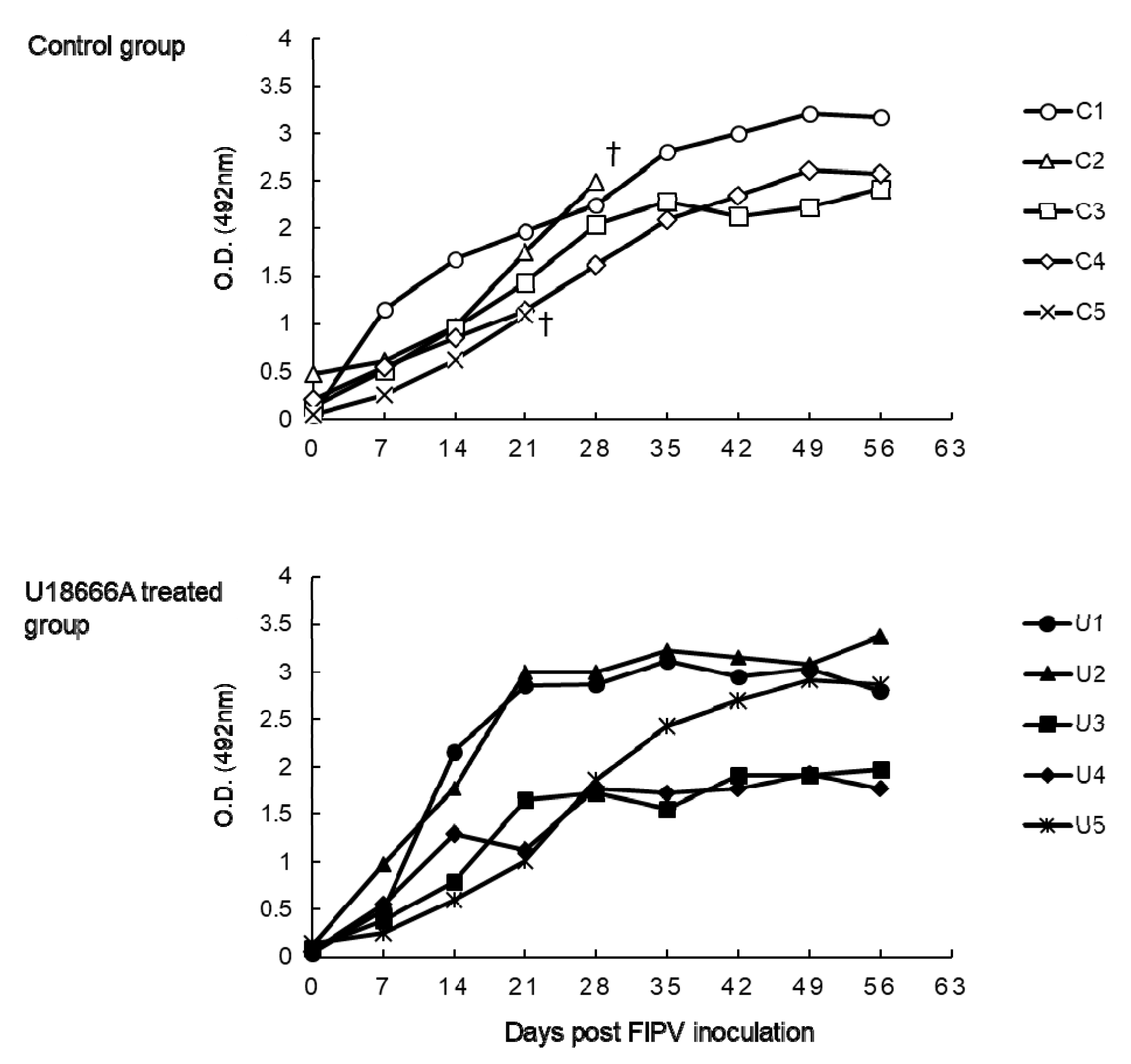
2.3. The Changes on Anti-FCoV Antibody of FIPV-Infected Cats
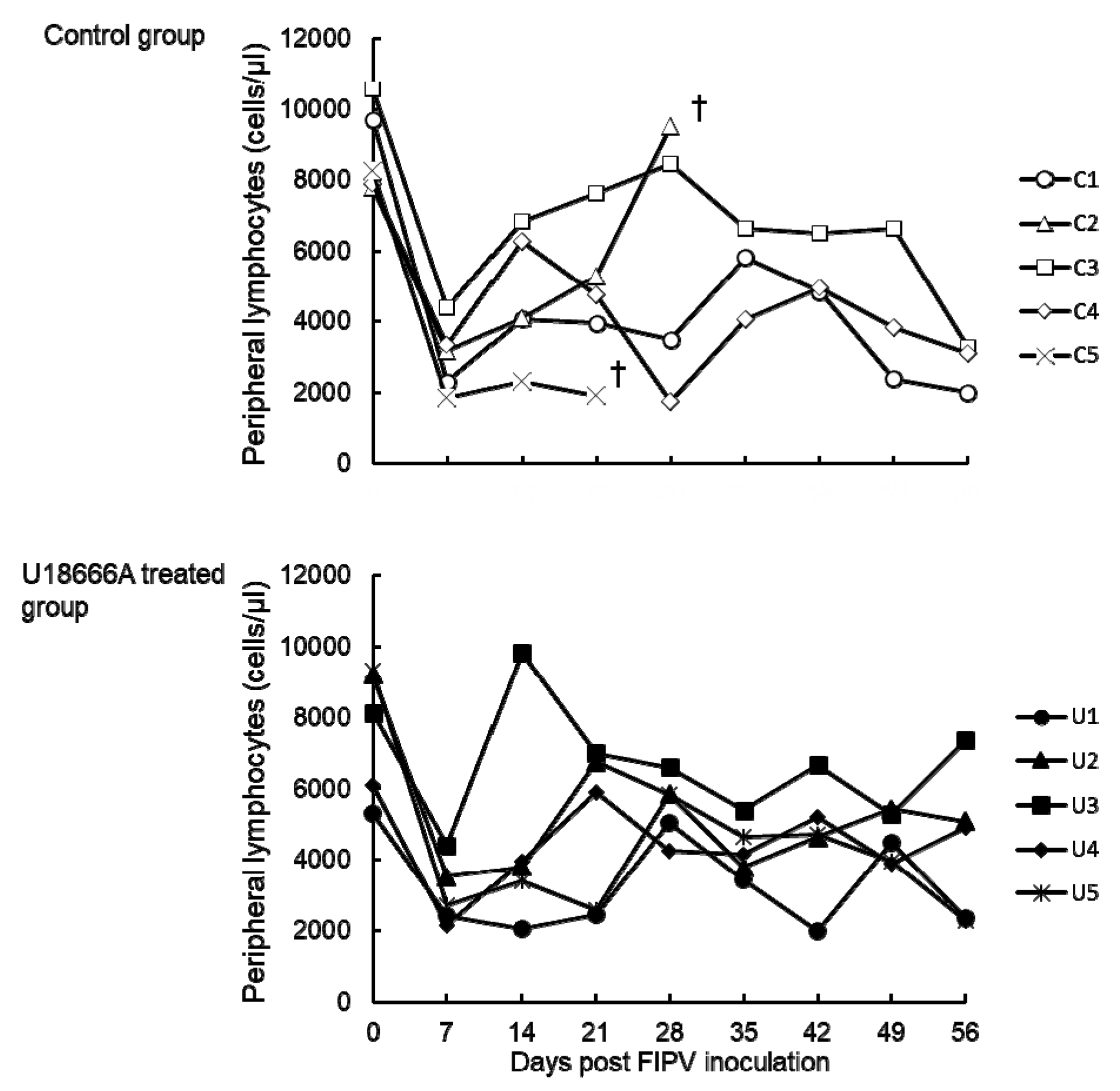
2.4. Changes in the Peripheral Blood Lymphocyte Count
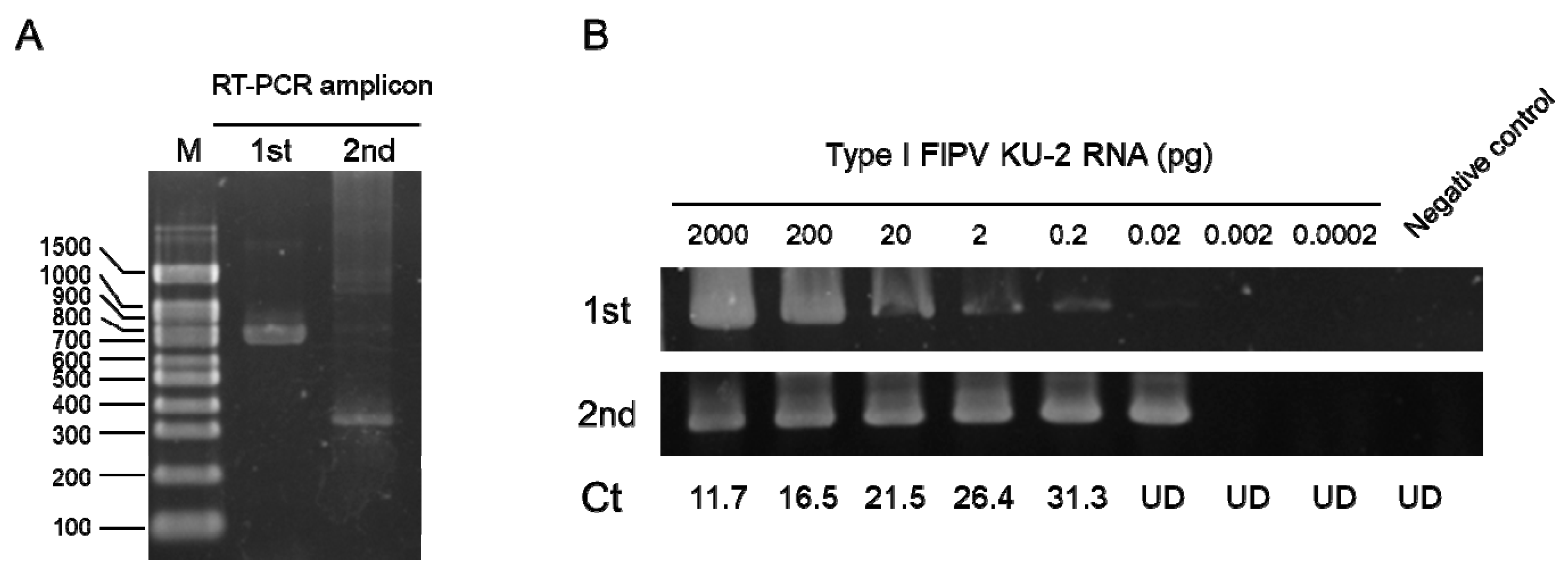
2.5. Detection of the FIPV N Gene in Feces, Saliva, and Plasma Samples
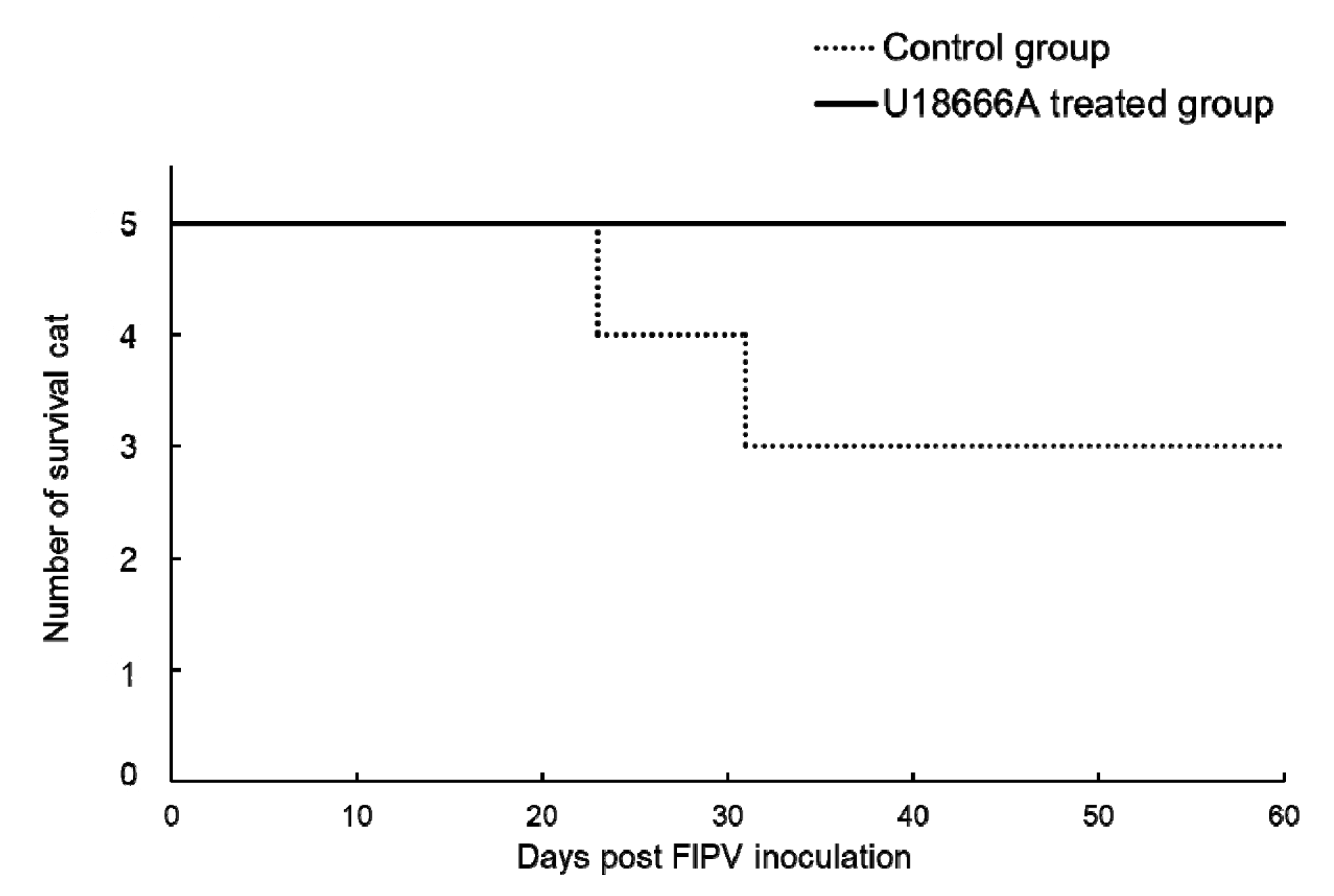
2.6. Survival Rate and Incidence of FIP
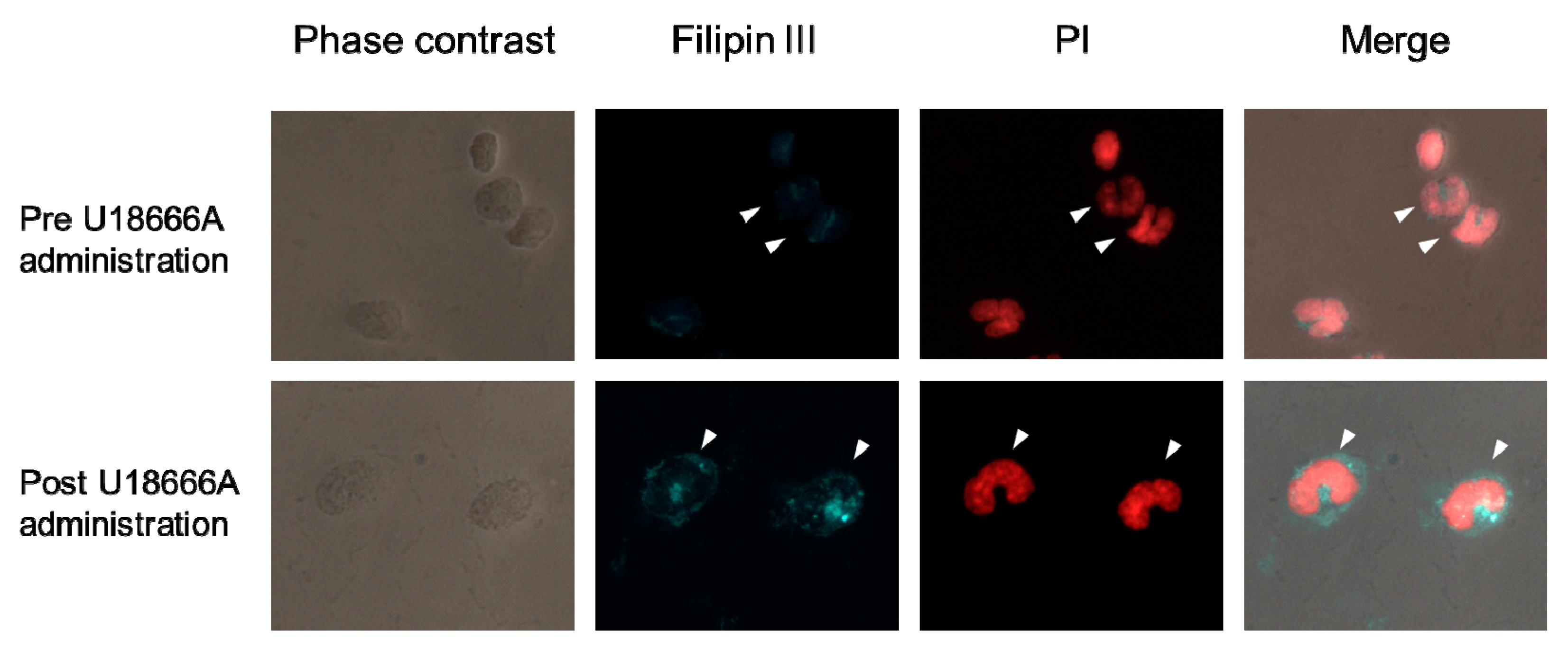
2.7. Localization of Intracellular Cholesterol in Peripheral Blood Monocytes of SPF Cat Administered a Single Dose of U18666A
3. Discussion
4. Materials and Methods
4.1. Virus
4.2. U18666A
4.3. Animal Experiments
4.4. ELISA
4.5. RNA Isolation and cDNA Preparation
4.6. Detection of Feline FIPV N Gene
4.7. Single Administration of U18666A and Staining of Cellular Cholesterol in Monocytes
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groot, R.; Cowley, J.; Enjuanes, L.; Faaberg, K.; Perlman, S.; Rottier, P.; Snijder, E.; Ziebuhr, J.; Gorbalenya, A. Order-Nidovirales. In Virus Taxonomy; King, A., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 784–794. ISBN 9780123846846. [Google Scholar]
- Whittaker, G.R.; André, N.M.; Millet, J.K. Improving Virus Taxonomy by Recontextualizing Sequence-Based Classification with Biologically Relevant Data: The Case of the Alphacoronavirus 1 Species. MSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekes, G.; Thiel, H.J. Feline Coronaviruses: Pathogenesis of Feline Infectious Peritonitis. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2016; pp. 193–218. ISBN 9780128047361. [Google Scholar]
- Jaimes, J.A.; Whittaker, G.R. Feline coronavirus: Insights into viral pathogenesis based on the spike protein structure and function. Virology 2018, 517, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Terada, Y.; Matsui, N.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Soma, T.; Mochizuki, M.; Maeda, K. Emergence of pathogenic coronaviruses in cats by homologous recombination between feline and canine coronaviruses. PLoS ONE 2014, 9, e106534. [Google Scholar] [CrossRef] [Green Version]
- Decaro, N.; Mari, V.; Campolo, M.; Lorusso, A.; Camero, M.; Elia, G.; Martella, V.; Cordioli, P.; Enjuanes, L.; Buonavoglia, C. Recombinant Canine Coronaviruses Related to Transmissible Gastroenteritis Virus of Swine Are Circulating in Dogs. J. Virol. 2009, 83, 1532–1537. [Google Scholar] [CrossRef] [Green Version]
- Herrewegh, A.A.; Smeenk, I.; Horzinek, M.C.; Rottier, P.J.; de Groot, R.J. Feline coronavirus type II strains 79-1683 and 79-1146 originate from a double recombination between feline coronavirus type I and canine coronavirus. J. Virol. 1998, 72, 4508–4514. [Google Scholar] [CrossRef] [Green Version]
- Terada, Y.; Shiozaki, Y.; Shimoda, H.; Youssef, H.; Mahmoud, A.H.; Noguchi, K.; Nagao, Y.; Shimojima, M.; Iwata, H.; Mizuno, T.; et al. Feline infectious peritonitis virus with a large deletion in the 59-terminal region of the spike gene retains its virulence for cats. J. Gen. Virol. 2012, 93, 1930–1934. [Google Scholar] [CrossRef]
- Addie, D.D.; Schaap, I.A.T.; Nicolson, L.; Jarrett, O. Persistence and transmission of natural type I feline coronavirus infection. J. Gen. Virol. 2003, 84, 2735–2744. [Google Scholar] [CrossRef]
- Kummrow, M.; Meli, M.L.; Haessig, M.; Goenczi, E.; Poland, A.; Pedersen, N.C.; Hofmann-Lehmann, R.; Lutz, H. Feline coronavirus serotypes 1 and 2: Seroprevalence and association with disease in Switzerland. Clin. Diagn. Lab. Immunol. 2005, 12, 1209–1215. [Google Scholar] [CrossRef] [Green Version]
- Hohdatsu, T.; Okada, S.; Ishizuka, Y.; Yamada, H.; Koyama, H. The Prevalence of Types I and II Feline Coronavirus Infections in Cats. J. Vet. Med. Sci. 1992, 54, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, N.C. An update on feline infectious peritonitis: Diagnostics and therapeutics. Vet. J. 2014, 201, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasker, S. Diagnosis of feline infectious peritonitis: Update on evidence supporting available tests. J. Feline Med. Surg. 2018, 20, 228–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Lovell, S.; Tiew, K.-C.; Mandadapu, S.R.; Alliston, K.R.; Battaile, K.P.; Groutas, W.C.; Chang, K.-O. Broad-spectrum antivirals against 3C or 3C-like proteases of picornaviruses, noroviruses, and coronaviruses. J. Virol. 2012, 86, 11754–11762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Liu, H.; Galasiti Kankanamalage, A.C.; Weerasekara, S.; Hua, D.H.; Groutas, W.C.; Chang, K.O.; Pedersen, N.C. Reversal of the Progression of Fatal Coronavirus Infection in Cats by a Broad-Spectrum Coronavirus Protease Inhibitor. PLoS Pathog. 2016, 12, e1005531. [Google Scholar]
- Pedersen, N.C.; Kim, Y.; Liu, H.; Galasiti Kankanamalage, A.C.; Eckstrand, C.; Groutas, W.C.; Bannasch, M.; Meadows, J.M.; Chang, K.O. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J. Feline Med. Surg. 2018, 20, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.G.; Perron, M.; Murakami, E.; Bauer, K.; Park, Y.; Eckstrand, C.; Liepnieks, M.; Pedersen, N.C. The nucleoside analog GS-441524 strongly inhibits feline infectious peritonitis (FIP) virus in tissue culture and experimental cat infection studies. Vet. Microbiol. 2018, 219, 226–233. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Perron, M.; Bannasch, M.; Montgomery, E.; Murakami, E.; Liepnieks, M.; Liu, H. Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis. J. Feline Med. Surg. 2019, 21, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Ko, D.C.; Gordon, M.D.; Jin, J.Y.; Scott, M.P. Dynamic movements of organelles containing Niemann-Pick C1 protein: NPC1 involvement in late endocytic events. Mol. Biol. Cell 2001, 12, 601–614. [Google Scholar] [CrossRef]
- Poh, M.K.; Shui, G.; Xie, X.; Shi, P.Y.; Wenk, M.R.; Gu, F. U18666A, an intra-cellular cholesterol transport inhibitor, inhibits dengue virus entry and replication. Antivir. Res. 2012, 93, 191–198. [Google Scholar] [CrossRef]
- Sabino, C.; Basic, M.; Bender, D.; Elgner, F.; Himmelsbach, K.; Hildt, E.; Sabino, C.; Basic, M.; Bender, D.; Elgner, F.; et al. Bafilomycin A1 and U18666A Efficiently Impair ZIKV Infection. Viruses 2019, 11, 524. [Google Scholar] [CrossRef] [Green Version]
- Wichit, S.; Hamel, R.; Bernard, E.; Talignani, L.; Diop, F.; Ferraris, P.; Liegeois, F.; Ekchariyawat, P.; Luplertlop, N.; Surasombatpattana, P.; et al. Imipramine Inhibits Chikungunya Virus Replication in Human Skin Fibroblasts through Interference with Intracellular Cholesterol Trafficking. Sci. Rep. 2017, 7, 3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgner, F.; Ren, H.; Medvedev, R.; Ploen, D.; Himmelsbach, K.; Boller, K.; Hildt, E. The Intracellular Cholesterol Transport Inhibitor U18666A Inhibits the Exosome-Dependent Release of Mature Hepatitis C Virus. J. Virol. 2016, 90, 11181–11196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, T.; Endoh, M.; Fukatsu, H.; Sakurada, H.; Doki, T.; Hohdatsu, T. The cholesterol transport inhibitor U18666A inhibits type I feline coronavirus infection. Antivir. Res. 2017, 145, 96–102. [Google Scholar] [CrossRef]
- Cenedella, R.J.; Sarkar, C.P.; Towns, L. Studies on the Mechanism of the Epileptiform Activity Induced by U18666A. II. Concentration, Half-Life and Distribution of Radiolabeled U18666A in the Brain. Epilepsia 1982, 23, 257–268. [Google Scholar] [CrossRef]
- Takano, T.; Akiyama, M.; Doki, T.; Hohdatsu, T. Antiviral activity of itraconazole against type I feline coronavirus infection. Vet. Res. 2019, 50, 5. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Yamada, S.; Doki, T.; Hohdatsu, T. Pathogenesis of oral type I feline infectious peritonitis virus (FIPV) infection: Antibody-dependent enhancement infection of cats with type I FIPV via the oral route. J. Vet. Med. Sci. 2019, 81, 911–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, T.; Kawakami, C.; Yamada, S.; Satoh, R.; Hohdatsu, T. Antibody-dependent enhancement occurs upon re-infection with the identical serotype virus in feline infectious peritonitis virus infection. J. Vet. Med. Sci. 2008, 70, 1315–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.C.; Eckstrand, C.; Liu, H.; Leutenegger, C.; Murphy, B. Levels of feline infectious peritonitis virus in blood, effusions, and various tissues and the role of lymphopenia in disease outcome following experimental infection. Vet. Microbiol. 2015, 175, 157–166. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cat No. | Gender | Age (Months) | |
|---|---|---|---|
| Control group | C1 | Female | 13 |
| C2 | Male | 13 | |
| C3 | Male | 13 | |
| C4 | Male | 12 | |
| C5 | Female | 8 | |
| U18666A treated group | U1 | Female | 13 |
| U2 | Male | 13 | |
| U3 | Male | 12 | |
| U4 | Female | 12 | |
| U5 | Male | 8 |
| Days Post FIPV Inoculation | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cat No. | 0 | 1–3 | 4–6 | 7–9 | 10–12 | 13–15 | 16–18 | 19–21 | 22–24 | 25–27 | 28–30 | |
| Control group | C1 | − | − | − | − | − | − | − | − | − | − | − |
| C2 | − | − | − | − | − | − | + | − | − | − | − | |
| C3 | − | − | − | − | − | + | + | − | − | − | + | |
| C4 | − | − | − | − | − | − | − | − | − | − | − | |
| C5 | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | |
| U18666A treated group | U1 | − | − | + | − | − | − | − | + | − | − | − |
| U2 | − | − | − | − | − | − | − | − | − | − | − | |
| U3 | − | − | − | − | − | − | − | − | − | − | − | |
| U4 | − | − | − | − | − | − | − | − | − | − | − | |
| U5 | − | − | − | − | − | − | − | + | − | − | + | |
| Days Post FIPV Inoculation | ||||||||
|---|---|---|---|---|---|---|---|---|
| Cat No. | 0 | 2 | 4 | 6 | 8 | 10 | 14 | |
| Control group | C1 | − | − | − | − | − | + | − |
| C2 | − | − | − | − | − | − | − | |
| C3 | − | − | − | − | − | − | − | |
| C4 | − | − | − | − | − | − | − | |
| C5 | − | − | − | − | + | − | − | |
| U18666A treated group | U1 | − | − | − | − | − | − | + |
| U2 | − | − | − | − | − | − | − | |
| U3 | − | − | − | − | − | − | − | |
| U4 | − | − | − | − | − | + | − | |
| U5 | − | − | − | + | + | − | − | |
| Orientation | Nucleotide Sequence | Location | Length (bp) | Reference | |
|---|---|---|---|---|---|
| 1st | Forward | 5′-CAACTGGGGAGATGAACCTT-3′ | 23–42 | 788 | KC461235 |
| Reverse | 5′-GGTAGCATTTGGCAGCGTTA-3′ | 791–810 | |||
| 2nd | Forward | 5′-ATTGATGGAGTCTTCTGGGTTG-3′ | 324–345 | 362 | KC461235 |
| Reverse | 5′-TTGGCATTCTTAGGTGTTGTGTC-3′ | 663–685 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doki, T.; Tarusawa, T.; Hohdatsu, T.; Takano, T. In Vivo Antiviral Effects of U18666A Against Type I Feline Infectious Peritonitis Virus. Pathogens 2020, 9, 67. https://doi.org/10.3390/pathogens9010067
Doki T, Tarusawa T, Hohdatsu T, Takano T. In Vivo Antiviral Effects of U18666A Against Type I Feline Infectious Peritonitis Virus. Pathogens. 2020; 9(1):67. https://doi.org/10.3390/pathogens9010067
Chicago/Turabian StyleDoki, Tomoyoshi, Tomoyo Tarusawa, Tsutomu Hohdatsu, and Tomomi Takano. 2020. "In Vivo Antiviral Effects of U18666A Against Type I Feline Infectious Peritonitis Virus" Pathogens 9, no. 1: 67. https://doi.org/10.3390/pathogens9010067