Antibiotic and Antibiofilm Activities of Salvadora persica L. Essential Oils against Streptococcus mutans: A Detailed Comparative Study with Chlorhexidine Digluconate



Abstract
:1. Introduction
2. Results
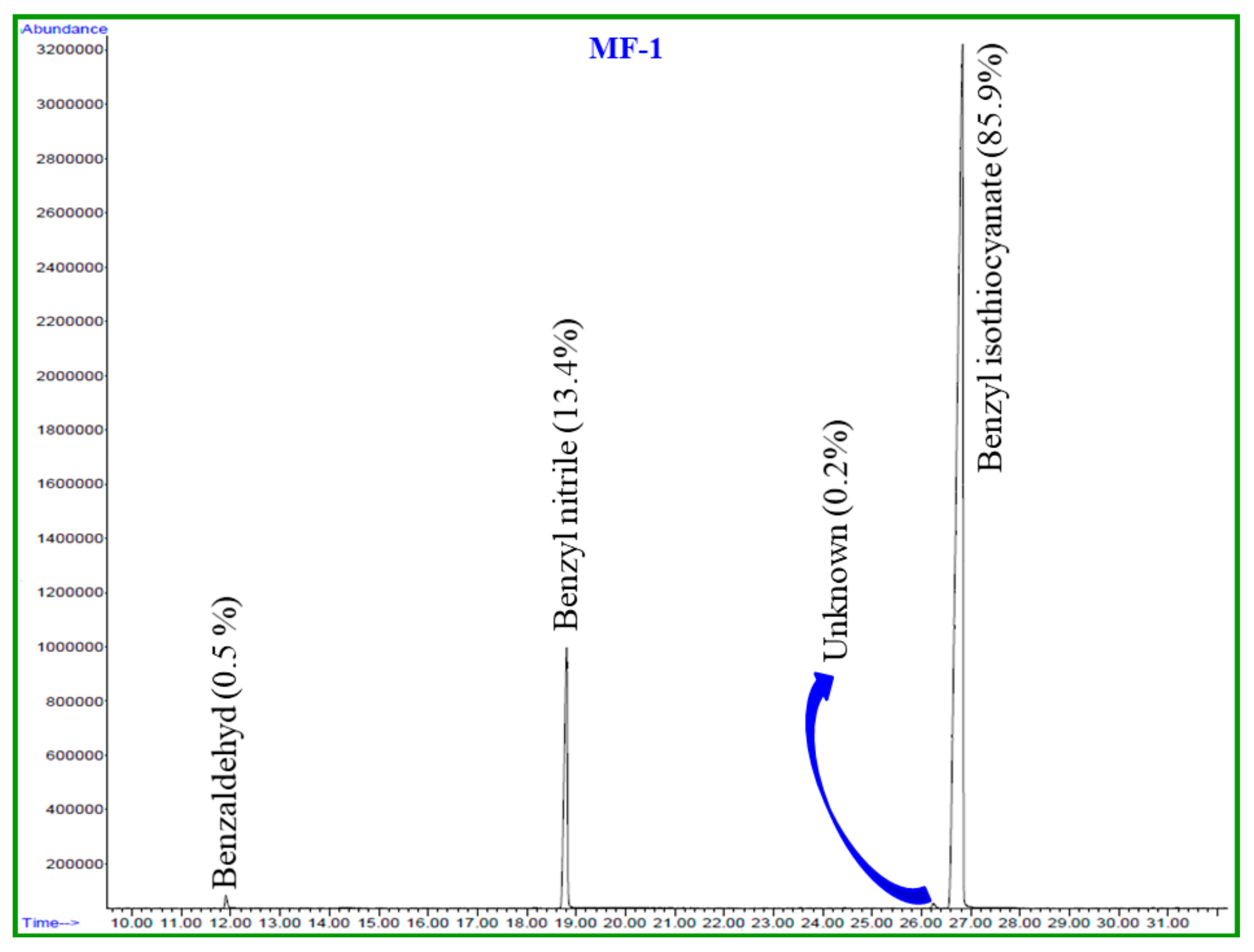
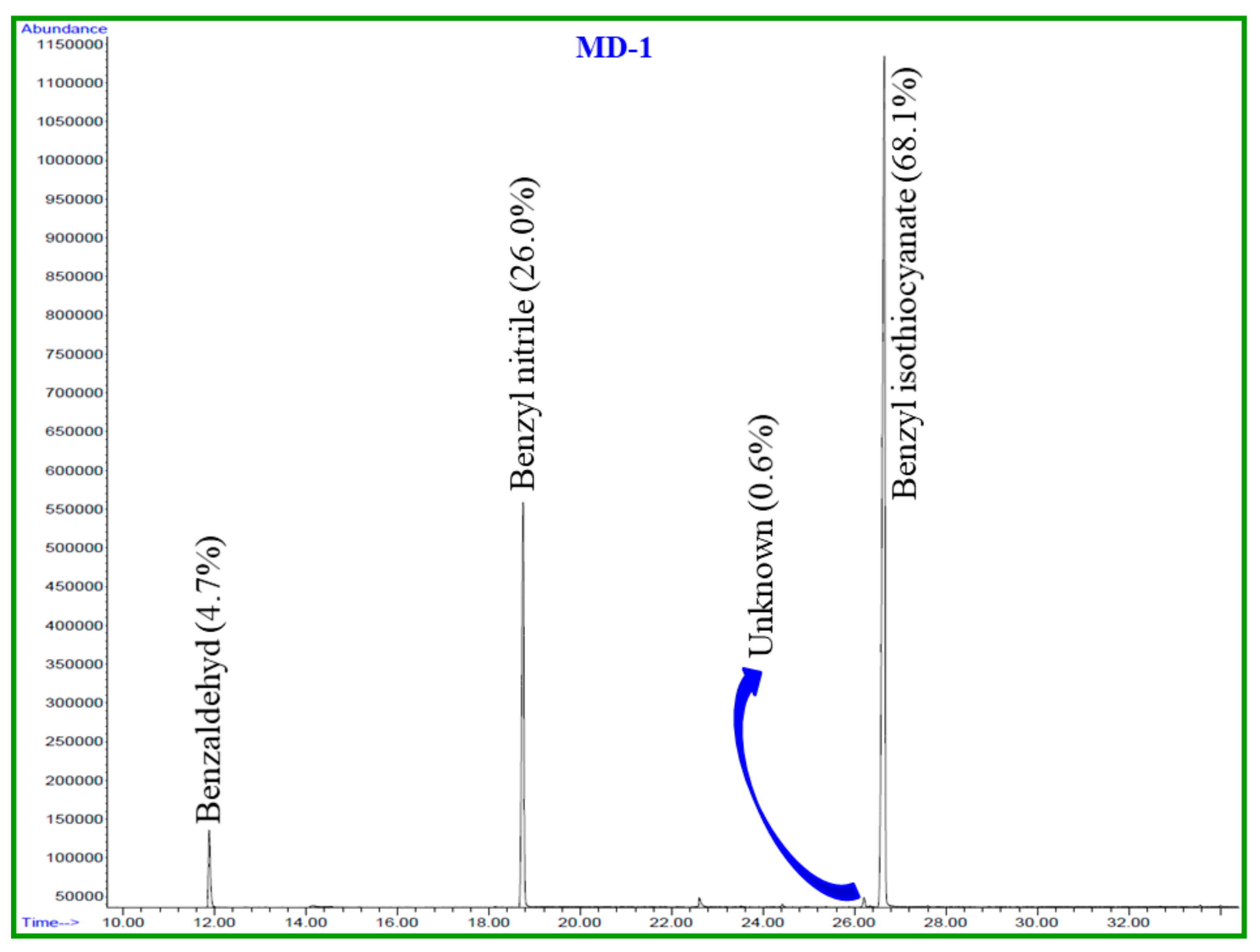
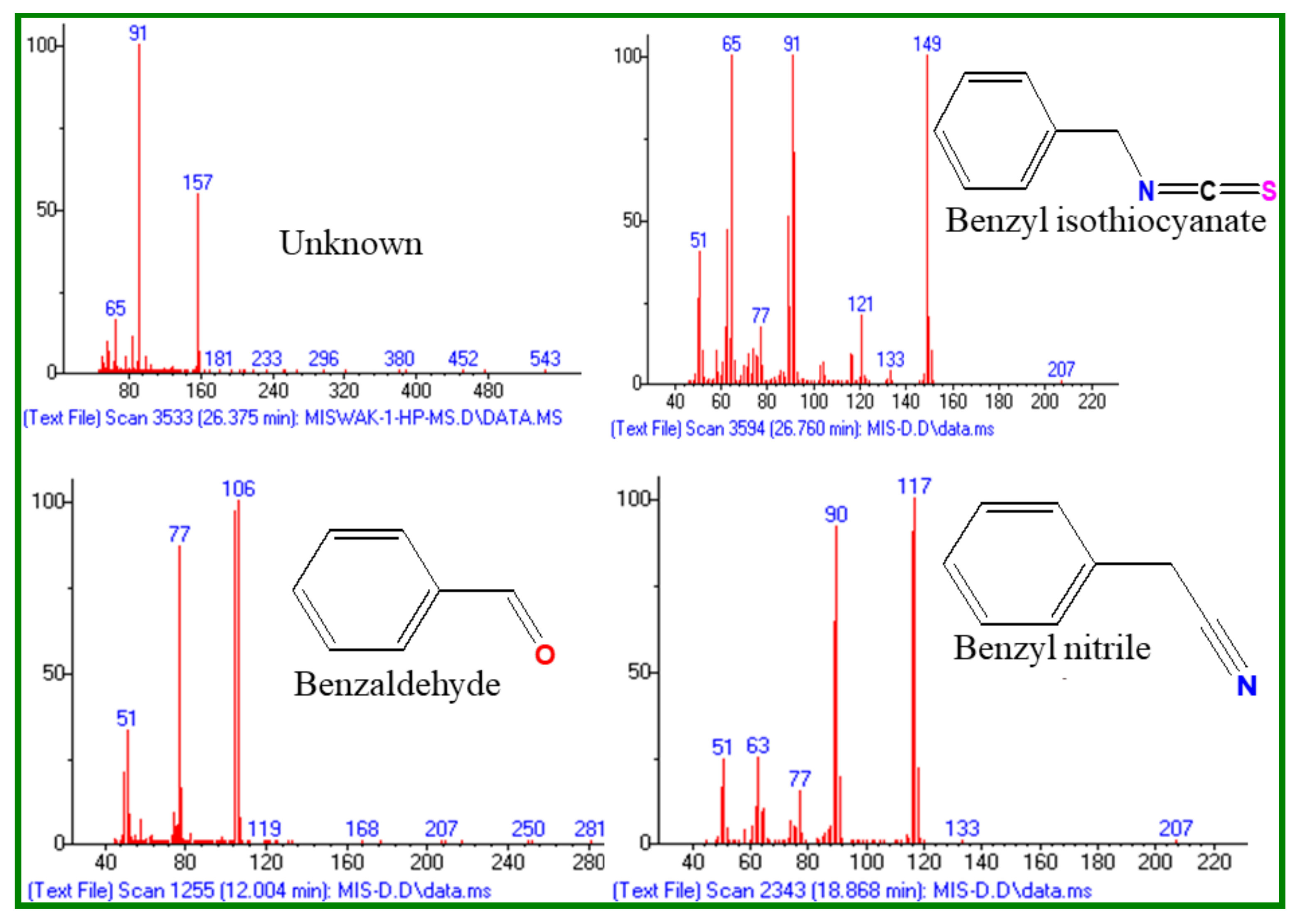
2.1. Chemical Composition of Essential Oils
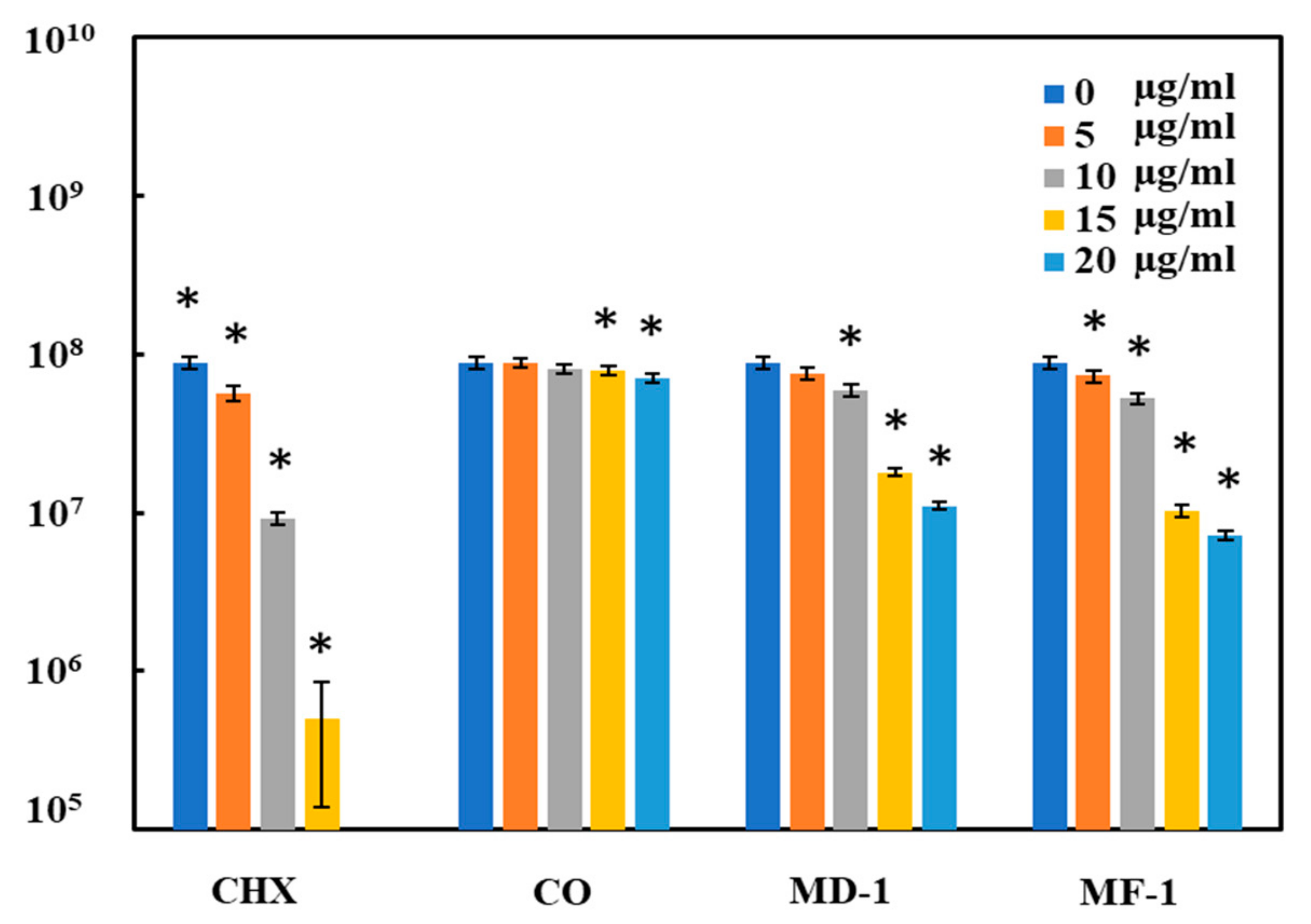
2.2. Antimicrobial Activity of Compounds
2.2.1. Microdilution Method
2.2.2. Spread Plate Method
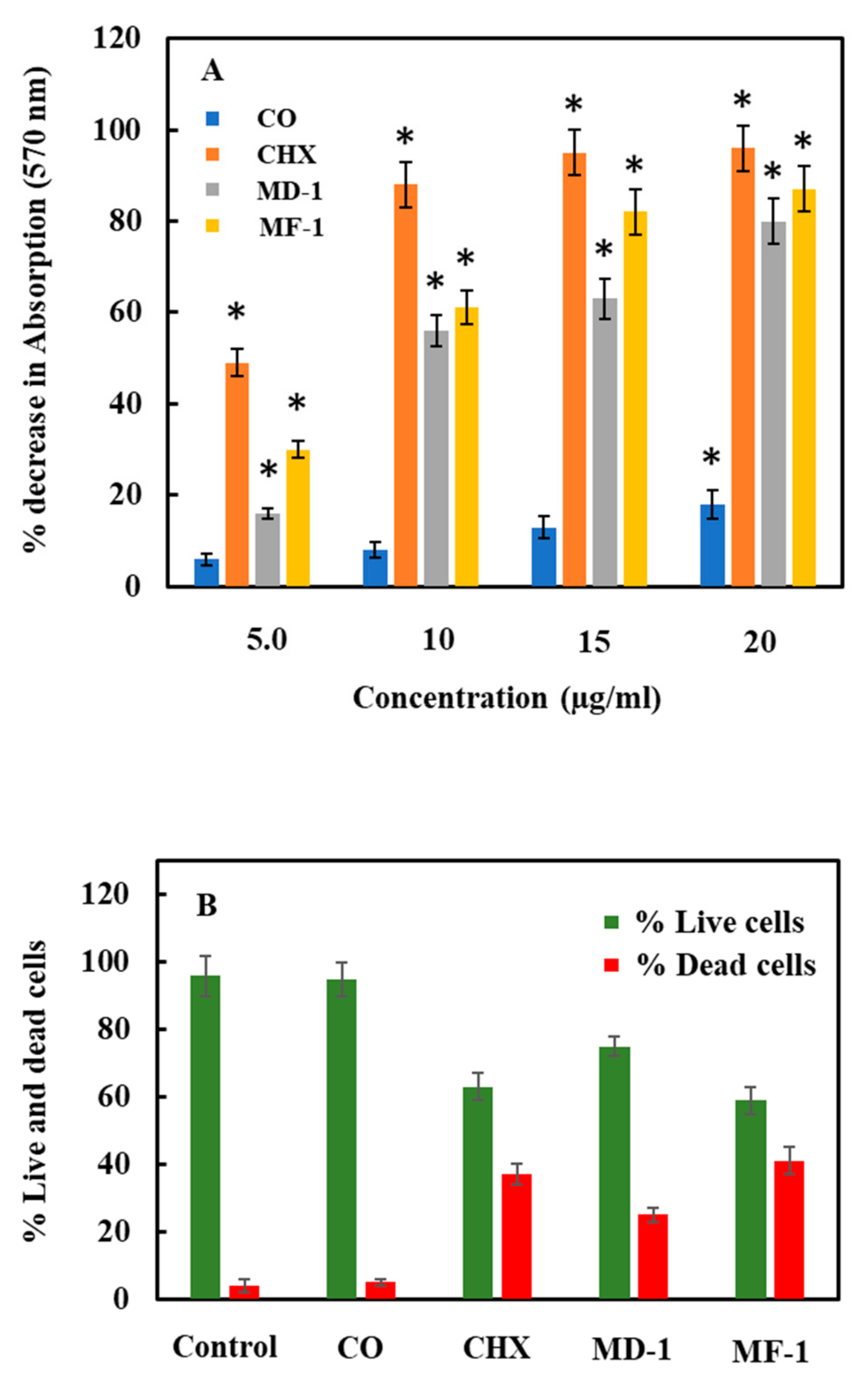
2.3. Change in Bacterial Viability
2.3.1. Viability Assay Using MTT
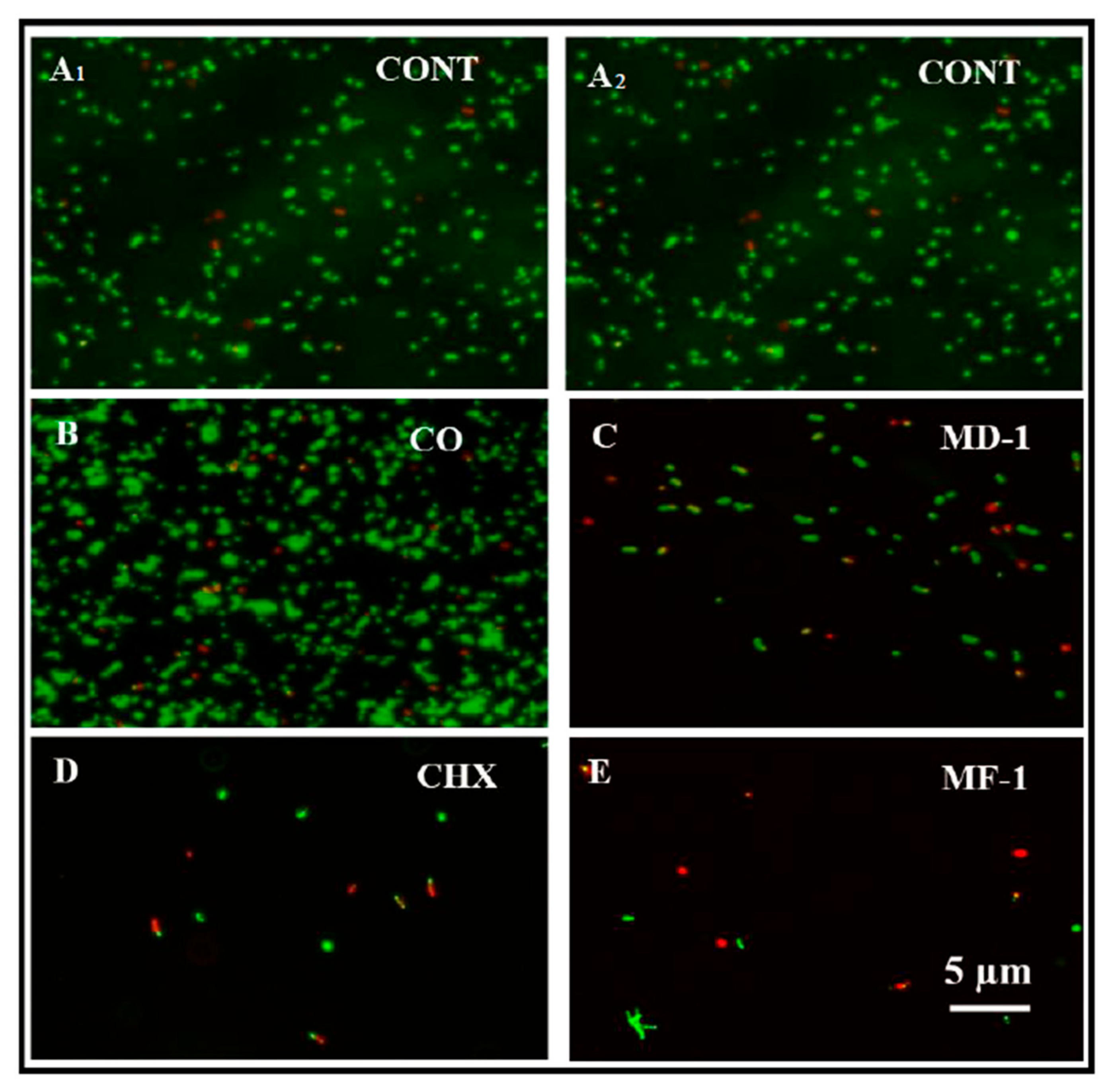
2.3.2. Live and Dead Staining
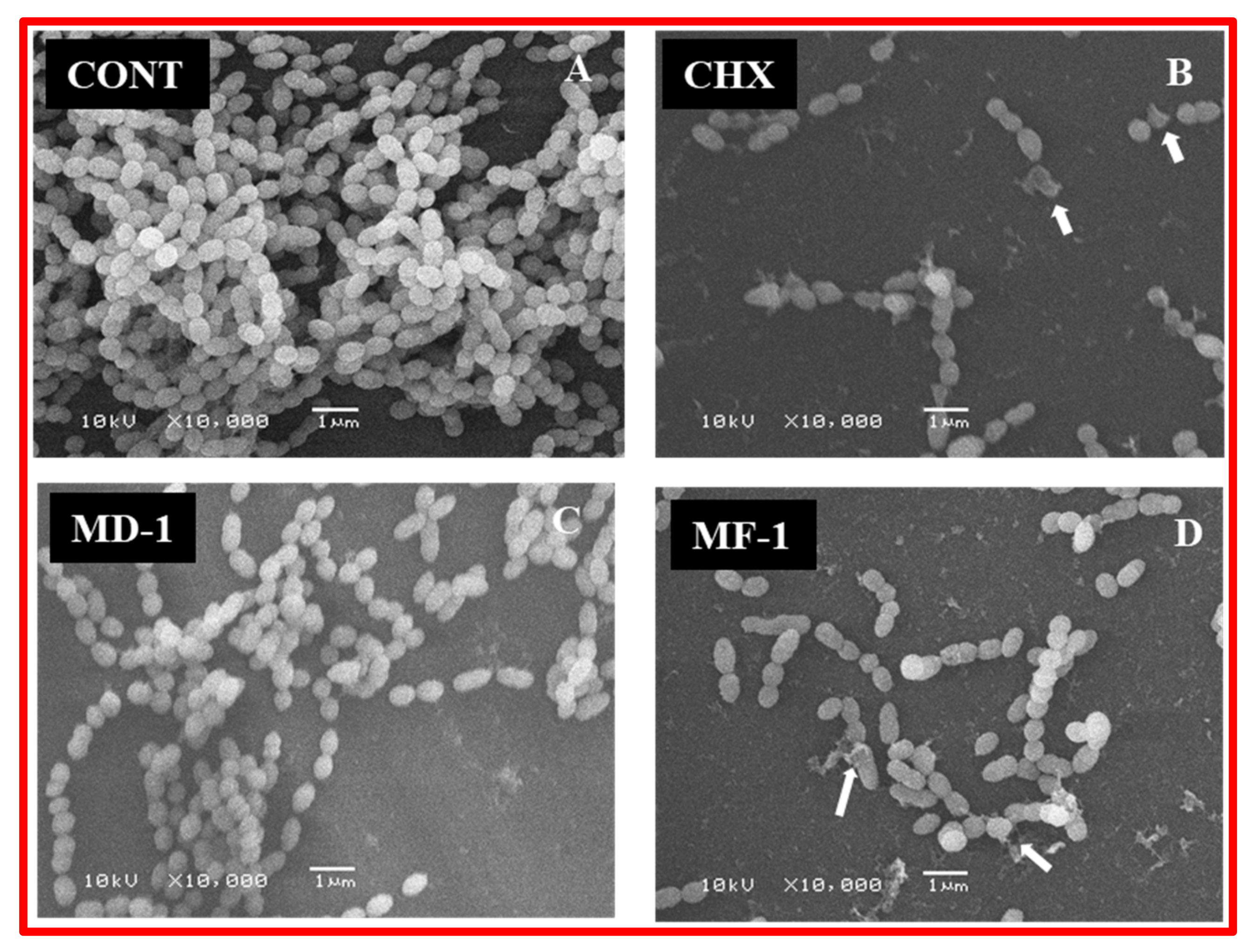
2.4. Change in Biofilm Formation Activity through Quantitative and Qualitative Assay
2.5. Real-Time PCR Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation of EOs
4.3. Characterization of EOs Constituents
4.3.1. GC and GC-MS Analysis of EOs
4.3.2. Linear Retention Indices (LRIs)
4.3.3. Identification of EOs Constituents
4.4. Culture Media and Bacterial Strains
4.5. Antimicrobial Activity against S. mutans
4.6. Bacterial Viability
4.7. Antibiofilm Activity
4.7.1. Crystal Violet Assay
4.7.2. Scanning Electron Microscopy of Biofilms
4.8. Real-Time PCR Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mabberley, D.J. Mabberley’s Plant-Book: A Portable Dictionary of Plants, Their Classification and Uses, 4th ed.; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar] [CrossRef]
- Collenette, S. An Illustrated Guide to the Flowers of Saudi Arabia; Scorpion Publishing: London, UK, 1985. [Google Scholar]
- Aumeeruddy, M.Z.; Zengin, G.; Mahomoodally, M.F. A review of the traditional and modern uses of Salvadora persica L. (Miswak): Toothbrush tree of Prophet Muhammad. J. Ethnopharmacol. 2018, 213, 409–444. [Google Scholar] [CrossRef] [PubMed]
- Khatak, M.; Khatak, S.; Siddqui, A.A.; Vasudeva, N.; Aggarwal, A.; Aggarwal, P. Salvadora persica. Pharm. Rev. 2010, 4, 209–214. [Google Scholar] [CrossRef]
- Haque, M.M.; Alsareii, S.A. A review of the therapeutic effects of using miswak (Salvadora persica) on oral health. Saudi Med. J. 2015, 36, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Halawany, H.S. A review on miswak (Salvadora persica) and its effect on various aspects of oral health. Saudi Dent. J. 2012, 24, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Al-Marri, A.H.; Khan, M.; Shaik, M.R.; Mohri, N.; Adil, S.F.; Kuniyil, M.; Alkhathlan, H.Z.; Al-Warthan, A.; Tremel, W. Green approach for the effective reduction of graphene oxide using Salvadora persica L. root (Miswak) extract. Nanoscale Res. Lett. 2015, 10, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Albalawi, G.H.; Shaik, M.R.; Khan, M.; Adil, S.F.; Kuniyil, M.; Alkhathlan, H.Z.; Al-Warthan, A.; Siddiqui, M.R.H. Miswak mediated green synthesized palladium nanoparticles as effective catalysts for the Suzuki coupling reactions in aqueous media. J. Saudi Chem. Soc. 2017, 21, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Balto, H.; Al-Sanie, I.; Al-Beshri, S.; Aldrees, A. Effectiveness of Salvadora persica extracts against common oral pathogens. Saudi Dent. J. 2017, 29, 1–6. [Google Scholar] [CrossRef]
- Al-Bayati, F.A.Y.; Sulaiman, K. In Vitro Antimicrobial Activity of Salvadora persica L. Extracts Against Some Isolated Oral Pathogens in Iraq. Turk. J. Biol. 2008, 32, 57–62. [Google Scholar]
- Sofrata, A.; Santangelo, E.M.; Azeem, M.; Borg-Karlson, A.-K.; Gustafsson, A.; Pütsep, K. Benzyl Isothiocyanate, a Major Component from the Roots of Salvadora Persica Is Highly Active against Gram-Negative Bacteria. PLoS ONE 2011, 6, e23045. [Google Scholar] [CrossRef]
- Han, Y.W.; Wang, X. Mobile microbiome: Oral bacteria in extra-oral infections and inflammation. J. Dent. Res. 2013, 92, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Högberg, L.D.; Heddini, A.; Cars, O. The global need for effective antibiotics: Challenges and recent advances. Trends Pharm. Sci. 2010, 31, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.T.; Khan, M.; Ahmad, J.; Wahab, R.; Abd-Elkader, O.H.; Musarrat, J.; Alkhathlan, H.Z.; Al-Kedhairy, A.A. Thymol and carvacrol induce autolysis, stress, growth inhibition and reduce the biofilm formation by Streptococcus mutans. Amb Express 2017, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eden, B.D. Chapter 16—Prevention Strategies for Periodontal Diseases. In Prevention in Clinical Oral Health Care; Cappelli, D.P., Mobley, C.C., Eds.; Mosby: St. Louis, MO, USA, 2008; pp. 213–229. [Google Scholar] [CrossRef]
- Leistevuo, J.; Järvinen, H.; Osterblad, M.; Leistevuo, T.; Huovinen, P.; Tenovuo, J. Resistance to mercury and antimicrobial agents in Streptococcus mutans isolates from human subjects in relation to exposure to dental amalgam fillings. Antimicrob. Agents Chemother. 2000, 44, 456–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeffler, J.M.; Garbino, J.; Lew, D.; Harbarth, S.; Rohner, P. Antibiotic consumption, bacterial resistance and their correlation in a Swiss university hospital and its adult intensive care units. Scand. J. Infect. Dis. 2003, 35, 843–850. [Google Scholar] [CrossRef]
- Li, X.; Kolltveit, K.M.; Tronstad, L.; Olsen, I. Systemic diseases caused by oral infection. Clin. Microbiol. Rev. 2000, 13, 547–558. [Google Scholar] [CrossRef]
- Khan, S.; Al-Khedhairy, A.; Musarrat, J. ZnO and TiO2 nanoparticles as novel antimicrobial agents for oral hygiene: A review. J. Nanopart. Res. 2015, 17. [Google Scholar] [CrossRef]
- Khan, S.T.; Al-Khedhairy, A.A.; Musarrat, J.; Ahamed, M. Application of nanoparticles in oral hygiene. Biomater. Tissue Eng. Bull. 2016, 3, 35–49. [Google Scholar] [CrossRef]
- Ajdić, D.; McShan, W.M.; McLaughlin, R.E.; Savić, G.; Chang, J.; Carson, M.B.; Primeaux, C.; Tian, R.; Kenton, S.; Jia, H.; et al. Genome sequence of Streptococcus mutans UA159, a cariogenic dental pathogen. Proc. Natl. Acad. Sci. USA 2002, 99, 14434–14439. [Google Scholar] [CrossRef] [Green Version]
- Dufour, V.; Stahl, M.; Rosenfeld, E.; Stintzi, A.; Baysse, C. Insights into the mode of action of benzyl isothiocyanate on Campylobacter jejuni. Appl. Environ. Microbiol. 2013, 79, 6958–6968. [Google Scholar] [CrossRef] [Green Version]
- Nowicki, D.; Rodzik, O.; Herman-Antosiewicz, A.; Szalewska-Pałasz, A. Isothiocyanates as effective agents against enterohemorrhagic Escherichia coli: Insight to the mode of action. Sci. Rep. 2016, 6, 22263. [Google Scholar] [CrossRef] [Green Version]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, F.; Huang, Z.; Ma, L.; Chen, L.; Pan, Y.; Xu, J.; Kim, S.; Kinane, D.; Koo, H.; et al. Meeting report: A close look at oral biofilms and microbiomes. Int. J. Oral Sci. 2018, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Romeo, L.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. Isothiocyanates: An Overview of Their Antimicrobial Activity against Human Infections. Molecules 2018, 23, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Mahmood, A.; Alkhathlan, H.Z. Characterization of leaves and flowers volatile constituents of Lantana camara growing in central region of Saudi Arabia. Arab. J. Chem. 2016, 9, 764–774. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Al-Saleem, M.S.; Alkhathlan, H.Z. A detailed study on chemical characterization of essential oil components of two Plectranthus species grown in Saudi Arabia. J. Saudi Chem. Soc. 2016, 20, 711–721. [Google Scholar] [CrossRef]
- Mshana, R.N.; Tadesse, G.; Abate, G.; Miorner, H. Use of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide for rapid detection of rifampin-resistant Mycobacterium tuberculosis. J. Clin. Microbiol. 1998, 36, 1214–1219. [Google Scholar] [CrossRef] [Green Version]
- Burton, E.; Yakandawala, N.; LoVetri, K.; Madhyastha, M.S. A microplate spectrofluorometric assay for bacterial biofilms. J. Ind. Microbiol. Biotechnol. 2007, 34, 1–4. [Google Scholar] [CrossRef]
- Ahn, S.J.; Burne, R.A. The atlA operon of Streptococcus mutans: Role in autolysin maturation and cell surface biogenesis. J. Bacteriol. 2006, 188, 6877–6888. [Google Scholar] [CrossRef] [Green Version]
- Carabetta, V.J.; Tanner, A.W.; Greco, T.M.; Defrancesco, M.; Cristea, I.M.; Dubnau, D. A complex of YlbF, YmcA and YaaT regulates sporulation, competence and biofilm formation by accelerating the phosphorylation of Spo0A. Mol. Microbiol. 2013, 88, 283–300. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Higuchi, M.; Poole, L.B.; Kamio, Y. Role of the dpr product in oxygen tolerance in Streptococcus mutans. J. Bacteriol. 2000, 182, 3740–3747. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | MF | MW | RT (min) | LRILit | LRIExpa | MF-1 (%) | MD-1 (%) |
|---|---|---|---|---|---|---|---|---|
| 1 | Benzaldehyde | C7H6O | 106 | 11.9 | 952 | 958 | 0.5 | 4.7 |
| 2 | Benzyl nitrile | C8H7N | 117 | 18.8 | - | 1140 | 13.4 | 26.0 |
| 3 | Unknown | - | - | 26.2 | - | 1355 | 0.2 | 0.6 |
| 4 | Benzyl Isothiocyanate | C8H7NS | 149 | 26.8 | - | 1373 | 85.8 | 68.1 |
| Total identified | 99.7 | 98.8 | ||||||
| Oil Yield | 0.2 | 0.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.; Alkhathlan, H.Z.; Khan, S.T. Antibiotic and Antibiofilm Activities of Salvadora persica L. Essential Oils against Streptococcus mutans: A Detailed Comparative Study with Chlorhexidine Digluconate. Pathogens 2020, 9, 66. https://doi.org/10.3390/pathogens9010066
Khan M, Alkhathlan HZ, Khan ST. Antibiotic and Antibiofilm Activities of Salvadora persica L. Essential Oils against Streptococcus mutans: A Detailed Comparative Study with Chlorhexidine Digluconate. Pathogens. 2020; 9(1):66. https://doi.org/10.3390/pathogens9010066
Chicago/Turabian StyleKhan, Merajuddin, Hamad Z. Alkhathlan, and Shams Tabrez Khan. 2020. "Antibiotic and Antibiofilm Activities of Salvadora persica L. Essential Oils against Streptococcus mutans: A Detailed Comparative Study with Chlorhexidine Digluconate" Pathogens 9, no. 1: 66. https://doi.org/10.3390/pathogens9010066