
Genome Characteristics of Two Ranavirus Isolates from Mandarin Fish and Largemouth Bass

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
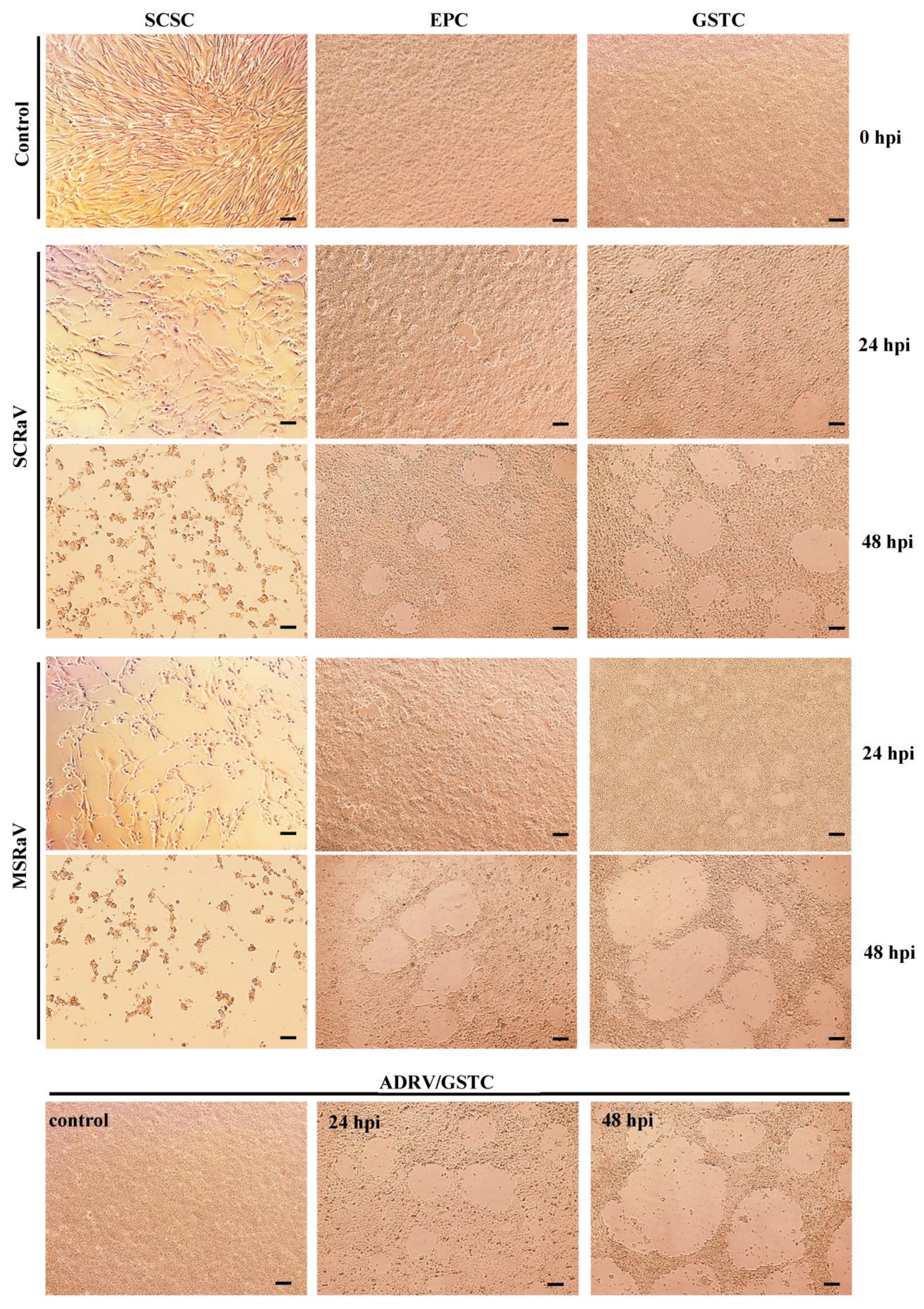
2.2. Virus Isolation
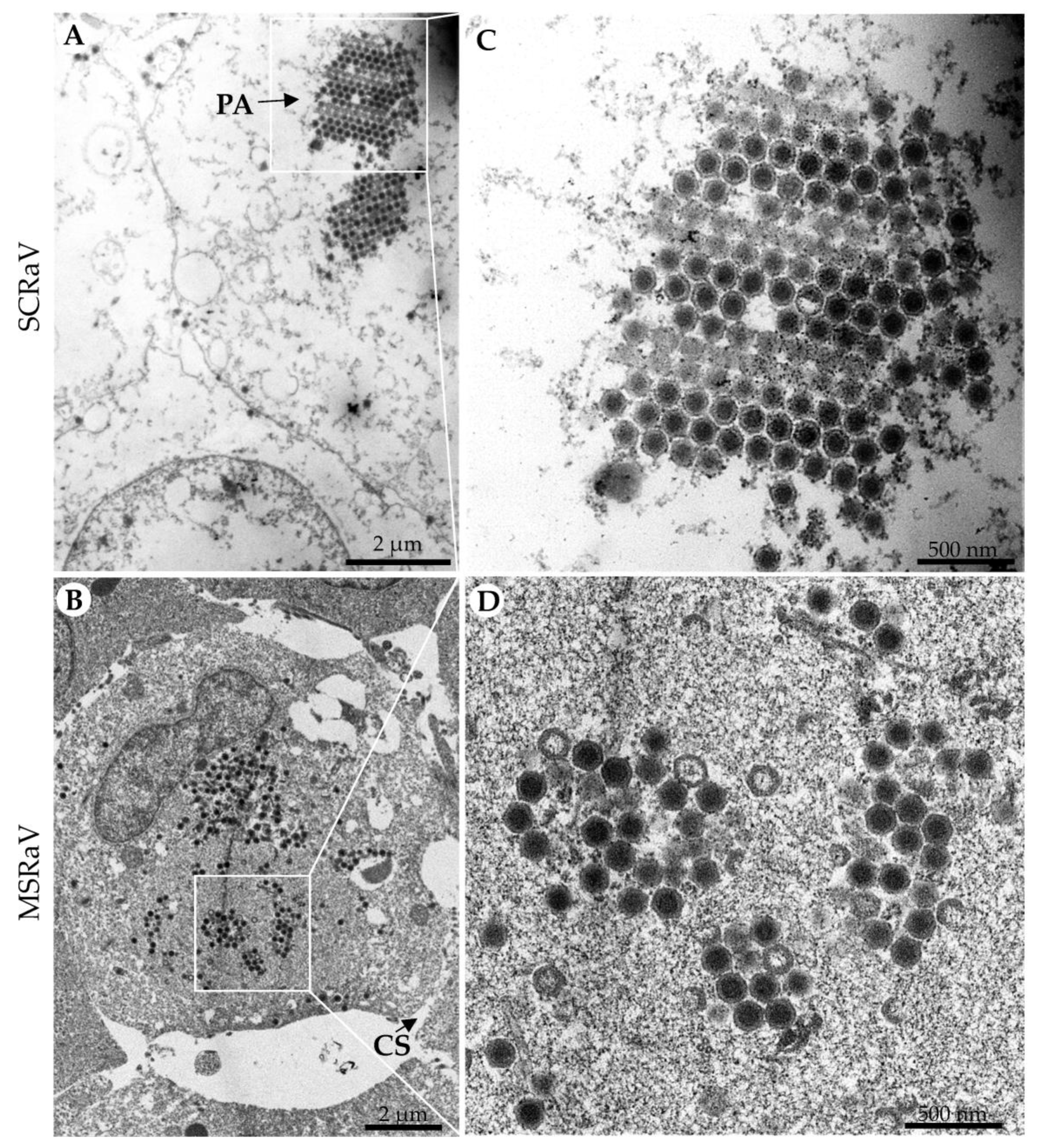
2.3. Electron Microscopy
2.4. Genomic DNA Extraction and Sequencing
2.5. Genome Annotation and Analysis
3. Results
3.1. Virus Isolation and Identification
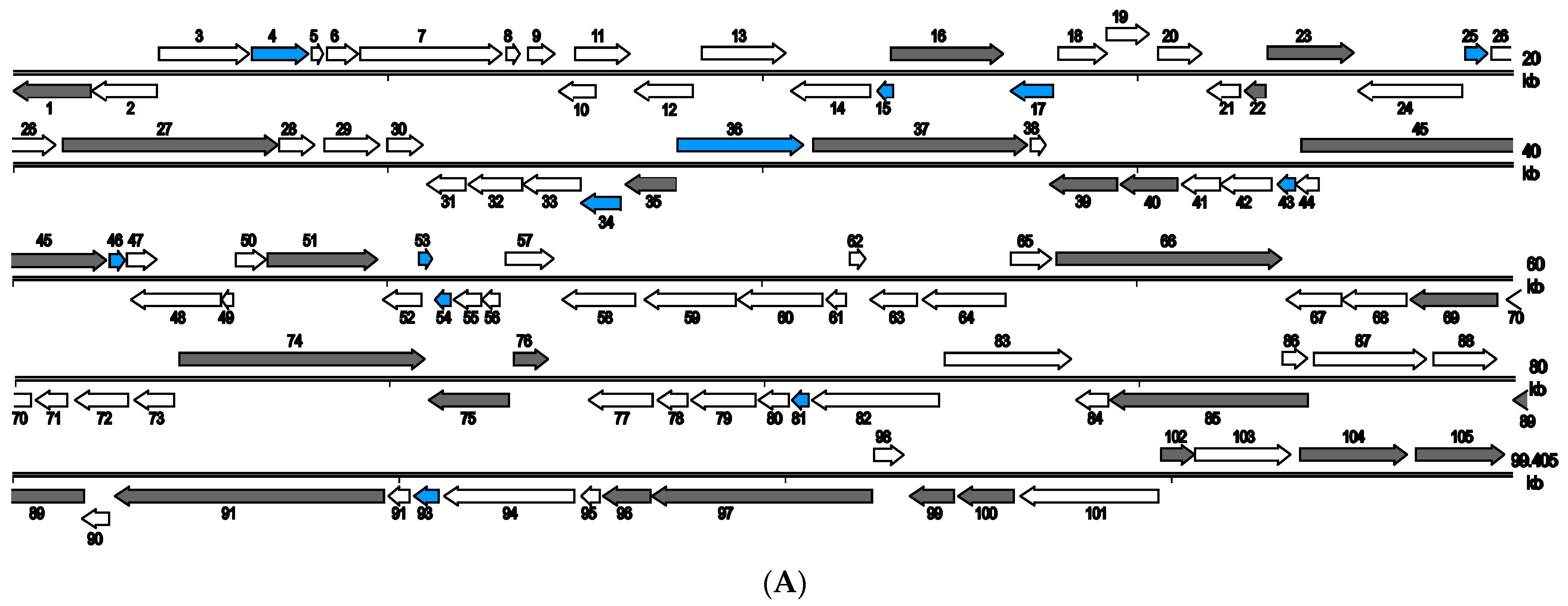
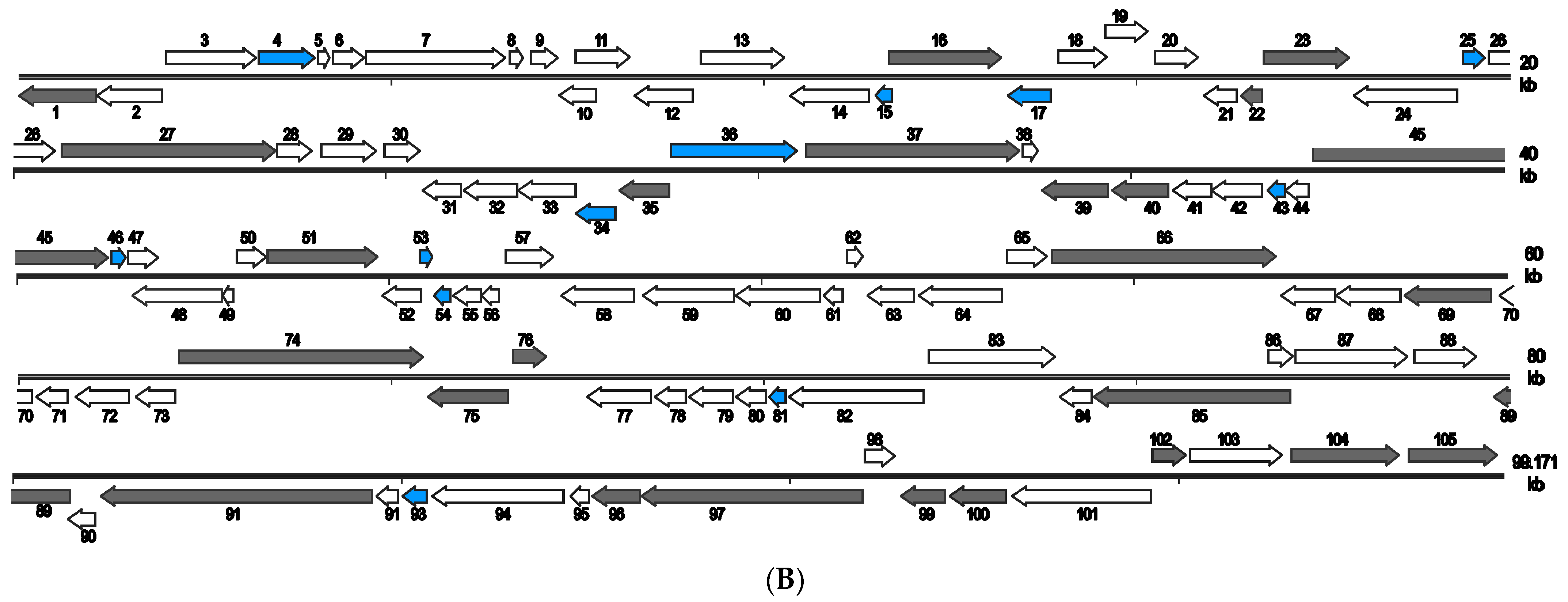
3.2. Architecture and General Features of the Two Virus Genomes
3.3. Structural Proteins
3.4. Nucleotide Metabolism Related Genes
3.5. DNA Replication- and Transcription-Related Genes
3.6. Virus–Host Interaction Related Genes
3.7. SCRaV- and MSRaV-Specific Genes
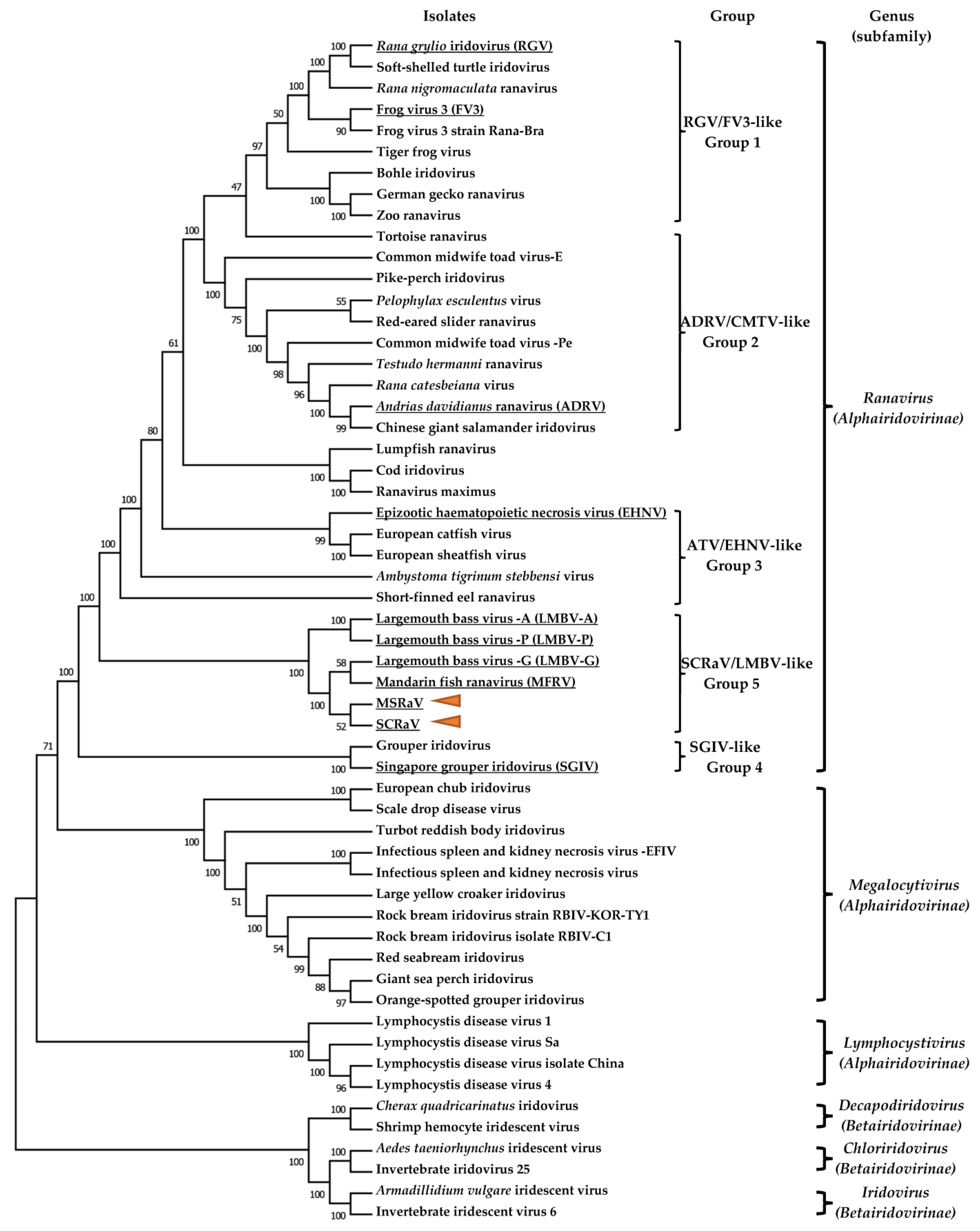
3.8. Phylogenetic Analysis
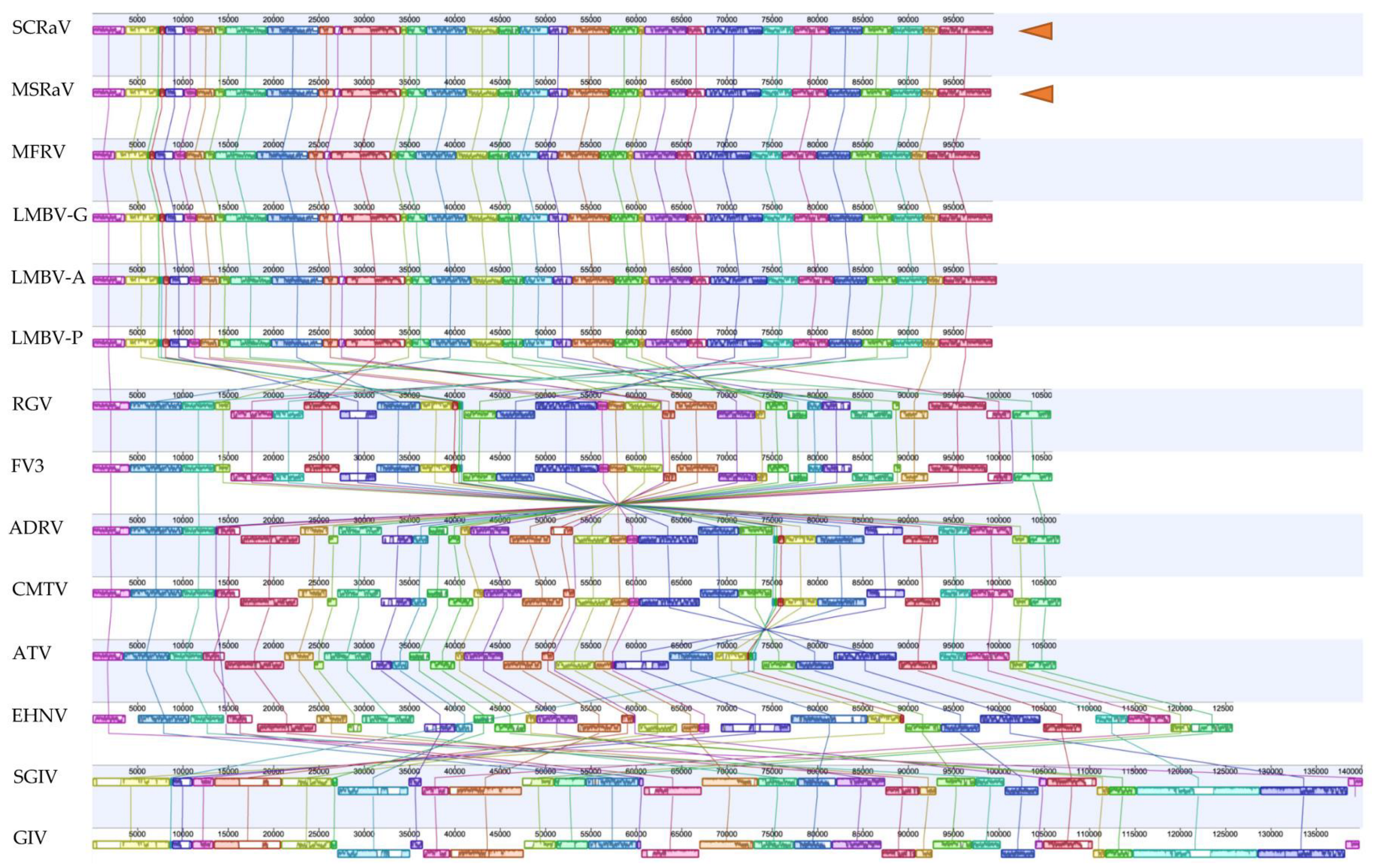
3.9. Genome Comparison
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinchar, V.G.; Hick, P.; Ince, I.A.; Jancovich, J.K.; Marschang, R.; Qin, Q.; Subramaniam, K.; Waltzek, T.B.; Whittington, R.; Williams, T.; et al. ICTV Virus Taxonomy Profile: Iridoviridae. J. Gen. Virol. 2017, 98, 890–891. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.W.; Chang, S.F.; Ngoh-Lim, G.H.; Gibson-Kueh, S.; Shi, C.; Lam, T.J. Characterization of a novel ranavirus isolated from grouper Epinephelus tauvina. Dis. Aquat. Org. 2003, 53, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jancovich, J.K.; Bremont, M.; Touchman, J.W.; Jacobs, B.L. Evidence for multiple recent host species shifts among the Ranaviruses (family Iridoviridae). J. Virol. 2010, 84, 2636–2647. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Geng, Y.; Zhao, R.; Gray, M.J.; Wang, K.; Ouyang, P.; Chen, D.; Huang, X.; Chen, Z.; Huang, C.; et al. CMTV-like ranavirus infection associated with high mortality in captive catfish-like loach, Triplophysa siluorides, in China. Transbound. Emerg. Dis. 2020, 67, 1330–1335. [Google Scholar] [PubMed]
- Zhang, Q.Y.; Xiao, F.; Li, Z.Q.; Gui, J.F.; Mao, J.; Chinchar, V.G. Characterization of an iridovirus from the cultured pig frog Rana grylio with lethal syndrome. Dis. Aquat. Organ. 2001, 48, 27–36. [Google Scholar]
- Chen, Z.Y.; Gui, J.F.; Gao, X.C.; Pei, C.; Hong, Y.J.; Zhang, Q.Y. Genome architecture changes and major gene variations of Andrias davidianus ranavirus (ADRV). Vet. Res. 2013, 44, 101. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Waltzek, T.B.; Subramaniam, K. Ranaviruses and other members of the family Iridoviridae: Their place in the virosphere. Virology 2017, 511, 259–271. [Google Scholar]
- Huang, Y.; Huang, X.; Liu, H.; Gong, J.; Ouyang, Z.; Cui, H.; Cao, J.; Zhao, Y.; Wang, X.; Jiang, Y.; et al. Complete sequence determination of a novel reptile iridovirus isolated from soft-shelled turtle and evolutionary analysis of Iridoviridae. BMC Genom. 2009, 10, 224. [Google Scholar] [CrossRef]
- Borzym, E.; Stachnik, M.; Reichert, M.; Rzezutka, A.; Jasik, A.; Waltzek, T.B.; Subramaniam, K. Genome Sequence of a Ranavirus Isolated from a Red-Eared Slider (Trachemys scripta elegans) in Poland. Microbiol. Resour. Announc. 2020, 9, e00781-20. [Google Scholar] [CrossRef]
- Claytor, S.C.; Subramaniam, K.; Landrau-Giovannetti, N.; Chinchar, V.G.; Gray, M.J.; Miller, D.L.; Mavian, C.; Salemi, M.; Wisely, S.; Waltzek, T.B. Ranavirus phylogenomics: Signatures of recombination and inversions among bullfrog ranaculture isolates. Virology 2017, 511, 330–343. [Google Scholar] [CrossRef]
- Conrad, C.R.K.; Subramaniam, K.; Chinchar, V.G.; Waltzek, T.B. Genomic Sequencing of Ranavirus Isolates from a Three-Spined Stickleback (Gasterosteus aculeatus) and a Red-Legged Frog (Rana aurora). Microbiol. Resour. Announc. 2021, 10, e0090221. [Google Scholar] [CrossRef] [PubMed]
- He, J.G.; Lu, L.; Deng, M.; He, H.H.; Weng, S.P.; Wang, X.H.; Zhou, S.Y.; Long, Q.X.; Wang, X.Z.; Chan, S.M. Sequence analysis of the complete genome of an iridovirus isolated from the tiger frog. Virology 2002, 292, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.Y.; Ou, T.; Zhu, R.L.; Zhang, Q.Y. Sequencing and analysis of the complete genome of Rana grylio virus (RGV). Arch. Virol. 2012, 157, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Mavian, C.; Lopez-Bueno, A.; Balseiro, A.; Casais, R.; Alcami, A.; Alejo, A. The genome sequence of the emerging common midwife toad virus identifies an evolutionary intermediate within ranaviruses. J. Virol. 2012, 86, 3617–3625. [Google Scholar] [CrossRef]
- Tan, W.G.; Barkman, T.J.; Gregory Chinchar, V.; Essani, K. Comparative genomic analyses of frog virus 3, type species of the genus Ranavirus (family Iridoviridae). Virology 2004, 323, 70–84. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2016. Available online: http://www.fao.org/3/a-i5555e.pdf (accessed on 11 April 2023).
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef]
- Gui, J.-F.; Zhou, L.; Li, X.-Y. Rethinking fish biology and biotechnologies in the challenge era for burgeoning genome resources and strengthening food security. Water Biol. Secur. 2022, 1, 100002. [Google Scholar] [CrossRef]
- Yu, X.J.; Xu, L.J.; Wu, F.X.; Song, D.D.; Gao, H.Q. China Fishery Statistical Yearbook. 2020. Available online: https://www.cafs.ac.cn/system/_content/download.jsp?urltype=news.DownloadAttachUrl&owner=1323872515&wbfileid=2829712 (accessed on 11 April 2023).
- Dong, C.F.; Wang, Z.M.; Weng, S.P.; He, J.G. Occurrence of a lethal ranavirus in hybrid mandarin (Siniperca scherzeri × Siniperca chuatsi) in Guangdong, South China. Vet. Microbiol. 2017, 203, 28–33. [Google Scholar] [CrossRef]
- Grizzle, J.M.; Altinok, I.; Fraser, W.A.; Francis-Floyd, R. First isolation of largemouth bass virus. Dis. Aquat. Organ. 2002, 50, 233–235. [Google Scholar] [CrossRef]
- Jin, Y.; Bergmann, S.M.; Mai, Q.; Yang, Y.; Liu, W.; Sun, D.; Chen, Y.; Yu, Y.; Liu, Y.; Cai, W.; et al. Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides). Viruses 2022, 14, 1643. [Google Scholar] [CrossRef]
- Ke, F.; Gui, L.; Li, T.; Li, F.H.; Zhao, X.; Chen, Z.Y.; Lei, C.K.; Zhang, Q.Y. The features of an emerging whispovirus isolate from freshwater crayfish. Aquacult. Rep. 2021, 20, 100728. [Google Scholar]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ke, F.; Huang, Y.H.; Zhao, J.G.; Gui, J.F.; Zhang, Q.Y. Identification and characterization of a novel envelope protein in Rana grylio virus. J. Gen. Virol. 2008, 89, 1866–1872. [Google Scholar] [CrossRef] [PubMed]
- He, L.B.; Ke, F.; Wang, J.; Gao, X.C.; Zhang, Q.Y. Rana grylio virus (RGV) envelope protein 2L: Subcellular localization and essential roles in virus infectivity revealed by conditional lethal mutant. J. Gen. Virol. 2014, 95, 679–690. [Google Scholar] [CrossRef]
- Ke, F.; Yu, X.-D.; Wang, Z.-H.; Gui, J.-F.; Zhang, Q.-Y. Replication and transcription machinery for ranaviruses: Components, correlation, and functional architecture. Cell Biosci. 2022, 12, 6. [Google Scholar] [CrossRef]
- Zeng, X.T.; Zhang, Q.Y. Interaction between two iridovirus core proteins and their effects on ranavirus (RGV) replication in cells from different species. Viruses 2019, 11, 416. [Google Scholar] [CrossRef]
- Ke, F.; Zhang, Q.Y. ADRV 12L: A Ranaviral Putative Rad2 Family Protein Involved in DNA Recombination and Repair. Viruses 2022, 14, 908. [Google Scholar] [CrossRef]
- Vilaca, S.T.; Bienentreu, J.F.; Brunetti, C.R.; Lesbarreres, D.; Murray, D.L.; Kyle, C.J. Frog virus 3 genomes reveal prevalent recombination between ranavirus lineages and their origins in Canada. J. Virol. 2019, 93, e00765-19. [Google Scholar] [CrossRef]
- Xu, Z.Q.; Dixon, N.E. Bacterial replisomes. Curr. Opin. Struct. Biol. 2018, 53, 159–168. [Google Scholar] [CrossRef]
- Kimura, M.; Ishihama, A. Involvement of multiple subunit-subunit contacts in the assembly of RNA polymerase II. Nucleic. Acids. Res. 2000, 28, 952–959. [Google Scholar] [CrossRef]
- Grimm, C.; Hillen, H.S.; Bedenk, K.; Bartuli, J.; Neyer, S.; Zhang, Q.; Huttenhofer, A.; Erlacher, M.; Dienemann, C.; Schlosser, A.; et al. Structural Basis of Poxvirus Transcription: Vaccinia RNA Polymerase Complexes. Cell 2019, 179, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zheng, C. The Race between Host Antiviral Innate Immunity and the Immune Evasion Strategies of Herpes Simplex Virus 1. Microbiol. Mol. Biol. Rev. 2020, 84, e00099-20. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.G.; Morgans, S.; Smith, E.; Aron, M.M.; Jancovich, J.K. The Ambystoma tigrinum virus (ATV) RNase III gene can modulate host PKR activation and interferon production. Virology 2017, 511, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Rothenburg, S.; Chinchar, V.G.; Dever, T.E. Characterization of a ranavirus inhibitor of the antiviral protein kinase PKR. BMC Microbiol. 2011, 11, 56. [Google Scholar] [CrossRef]
- Jancovich, J.K.; Jacobs, B.L. Innate immune evasion mediated by the Ambystoma tigrinum virus eukaryotic translation initiation factor 2alpha homologue. J. Virol. 2011, 85, 5061–5069. [Google Scholar] [CrossRef]
- Huynh, T.P.; Jancovich, J.K.; Tripuraneni, L.; Heck, M.C.; Langland, J.O.; Jacobs, B.L. Characterization of a PKR inhibitor from the pathogenic ranavirus, Ambystoma tigrinum virus, using a heterologous vaccinia virus system. Virology 2017, 511, 290–299. [Google Scholar] [CrossRef]
- Huang, X.H.; Huang, Y.H.; Cai, J.; Wei, S.N.; Gao, R.; Qin, Q.W. Identification and characterization of a tumor necrosis factor receptor like protein encoded by Singapore grouper iridovirus. Virus Res. 2013, 178, 340–348. [Google Scholar] [CrossRef]
- Andino Fde, J.; Grayfer, L.; Chen, G.; Chinchar, V.G.; Edholm, E.S.; Robert, J. Characterization of Frog Virus 3 knockout mutants lacking putative virulence genes. Virology 2015, 485, 162–170. [Google Scholar] [CrossRef]
- Yu, Y.; Huang, Y.; Ni, S.; Zhou, L.; Liu, J.; Zhang, J.; Zhang, X.; Hu, Y.; Huang, X.; Qin, Q. Singapore grouper iridovirus (SGIV) TNFR homolog VP51 functions as a virulence factor via modulating host inflammation response. Virology 2017, 511, 280–289. [Google Scholar] [CrossRef]
- Yan, Y.; Cui, H.; Guo, C.; Li, J.; Huang, X.; Wei, J.; Qin, Q. An insulin-like growth factor homologue of Singapore grouper iridovirus modulates cell proliferation, apoptosis and enhances viral replication. J. Gen. Virol. 2013, 94, 2759–2770. [Google Scholar] [CrossRef]
- Altindis, E.; Cai, W.; Sakaguchi, M.; Zhang, F.; GuoXiao, W.; Liu, F.; De Meyts, P.; Gelfanov, V.; Pan, H.; DiMarchi, R.; et al. Viral insulin-like peptides activate human insulin and IGF-1 receptor signaling: A paradigm shift for host-microbe interactions. Proc. Natl. Acad. Sci. USA 2018, 115, 2461–2466. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, A.P.; Willems, R.J.; Bonten, M.J.; van Schaik, W. LPxTG surface proteins of enterococci. Trends Microbiol. 2009, 17, 423–430. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF/aa | Nucleotide Position | Predicted Function/Conserved Domain | kDa | MSRaV c (OQ267587) | MFRV c (MG941005) | LMBV-G c (MW630113) | LMBV-A c (MK681855) | LMBV-P c (MK681856) | ADRV c (KC865735) | RGV c (JQ654586) | FV3 c (AY548484) | EHNV c (MT510742) | SGIV c (NC_006549) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ORF/AA | %ID d | ||||
| 1L/345 b | 1–1038 | myristylated membrane protein | 37.2 | 1L/345 | 100 | 123L/354 | 97.5 | 4L/345 | 100 | 1L/354 | 96.1 | 1L/354 | 96.1 | 2L/325 | 75.2 | 2L/323 | 75.2 | 2L/320 | 75.2 | 1L/350 | 74.4 | 19R/342 | 70.5 |
| 2L/290 | 1044–1916 | hypothetical protein | 33.2 | 2L/290 | 100 | 124L/290 | 100 | 5L/290 | 100 | 2L/290 | 98.6 | 2L/290 | 98.6 | 3L/291 | 45.1 | 3L/292 | 44 | 3L/279 | 44 | 2L/279 | 44.7 | 18R/285 | 36.7 |
| 3R/404 | 1943–3157 | hypothetical protein | 44.7 | 3R/404 | 100 | NA/NA | NA | 6R/404 | 100 | 3R/404 | 99.5 | 3R/404 | 99.5 | 4R/404 | 51.4 | 4R/404 | 51.2 | 3R/438 | 51.2 | 3R/404 | 50.8 | 16L/413 | 39 |
| 4R/253 | 3184–3945 | N-terminal immunoglobulin (Ig)-like domain | 27.3 | 4R/253 | 100 | 1R/253 | 100 | 7R/253 | 100 | 4R/253 | 98.4 | 4R/165 | 98.2 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 5R/54 | 3982–4146 | TM | 6.1 | 5R/54 | 100 | 2R/54 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 5R/60 | 45.5 | 5R/60 | 47.7 | 4R/60 | 47.7 | 4R/60 | 48.8 | 15L/59 | 45.7 |
| 6R/141 | 4187–4612 | hypothetical protein | 15.5 | 6R/141 | 100 | 3R/141 | 100 | 8R/141 | 100 | 5R/141 | 97.9 | 5R/141 | 97.9 | 80L/139 | 37.6 | 33R/104 | 35.4 | 31R/139 | 37.6 | 67R/139 | 37.6 | 14L/141 | 33.1 |
| 7R/634 | 4627–6531 | transcription termination factor Rho | 67.7 | 7R/624 | 98.4 | 4R/598 | 94.2 | 9R/396 | 89.4 | 6R/633 | 96.3 | 6R/621 | 94.4 | 79L/640 | 51 | 34R/644 | 51 | 32R/629 | 50 | 68R/658 | 49 | 12L/1024 | 31.1 |
| 8R/62 | 6581–6769 | TM | 6.8 | 8R/62 | 100 | 6R/62 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 78L/63 | 62.1 | 35R/63 | 62.1 | 33R/63 | 63.8 | 69R/63 | 63.8 | 11L/62 | 53.2 |
| 9R/119 | 6872–7231 | L protein-like protein, TM | 12.8 | 9R/119 | 100 | 8R/119 | 100 | 11R/99 | 100 | 7R/99 | 99 | 7R/99 | 99 | 77L/106 | 55.2 | 36R/106 | 53.1 | 34R/106 | 53.1 | 70R/107 | 52.5 | 9L/154 | 42.5 |
| 10L/163 | 7277–7768 | hypothetical protein | 16.2 | 10L/163 | 100 | 9L/166 | 98.2 | 12R/244 | 26.3 | 54/288 | 42.9 | NA/NA | NA | NA/NA | NA | 72R/115 | 47.7 | 36L/207 | 44.8 | NA/NA | NA | 7L/307 | 29.5 |
| 11R/245 | 7500–8237 | LPXTG-anchored collagen-like adhesin | 24.4 | 11R/245 | 100 | 10R/248 | 98.8 | 12R/244 | 98.8 | 9R/102 | 46.6 | 9R/102 | 46.6 | 75L/144 | 54.6 | 38R/91 | 39.2 | 65L/54 | 54 | 47L/112 | 51.4 | 8L/230 | 40.5 |
| 12L/261 | 8285–9070 | p31K protein | 29.3 | 12L/261 | 100 | 11L/261 | 100 | 13L/261 | 100 | 10L/262 | 99.2 | 10L/262 | 99.2 | 85L/261 | 77 | 27R/261 | 77 | 25R/262 | 77 | 60R/304 | 77 | 6R/259 | 64.4 |
| 13R/374 | 9190–10,314 | hypothetical protein | 42.3 | 13R/374 | 100 | 12R/374 | 100 | 14R/374 | 100 | 11R/374 | 98.9 | 11R/374 | 98.9 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | 4L/365 | 22.3 |
| 14L/354 | 10,373–11,437 | hypothetical protein | 39.2 | 14L/354 | 100 | 14L/354 | 100 | 15L/354 | 100 | 12L/354 | 99.4 | 12L/354 | 99.4 | 60R/237 | 65.9 | 52L/355 | 64.5 | 52L/355 | 64.8 | 54R/355 | 64.8 | 3R/381 | 55.8 |
| 15L/71 | 11,525–11,740 | hypothetical protein | 7.6 | 15L/71 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 16R/503 b | 11,706–13,217 | myristylated membrane protein | 53.1 | 16R/503 | 100 | 17R/503 | 100 | 16R/503 | 99.8 | 13R/503 | 99.2 | 13R/503 | 99.2 | 58L/522 | 63.7 | 53R/522 | 63.3 | 53R/522 | 63.5 | 53L/523 | 63.9 | 88L/506 | 55.4 |
| 17L/191 | 13,299–13,874 | Tumor necrosis factor receptor | 20.5 | 17L/191 | 100 | 19L/191 | 100 | 17L/191 | 100 | 14L/191 | 96.3 | 14L/191 | 96.3 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 18R/220 | 13,940–14,602 | DNA methyltransferase | 25.4 | 18R/220 | 100 | 20R/220 | 100 | 18R/220 | 100 | 15R/220 | 97.7 | 15R/220 | 97.7 | 24L/214 | 65.3 | 90R/214 | 64.8 | 83R/214 | 65.3 | 20L/214 | 65.3 | NA/NA | NA |
| 19R/192 | 14,587–15,165 | Methylase of polypeptide chain release factors | 20.5 | 19R/192 | 100 | 21R/177 | 100 | 19R/177 | 100 | 16R/177 | 94.9 | 16R/177 | 94.9 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 20R/197 | 15,271–15,864 | hypothetical protein | 22 | 20R/197 | 100 | 22R/197 | 100 | 20R/197 | 100 | 17R/196 | 97 | 17R/196 | 97 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 21L/147 | 15,925–16,368 | immediate early protein ICP-18 | 16.4 | 21L/147 | 100 | 23L/147 | 100 | 21L/147 | 100 | 18L/147 | 98.6 | 18L/147 | 98.6 | 26L/157 | 39.9 | 89R/157 | 40.5 | 82R/157 | 40.5 | 22L/157 | 39.2 | 86R/154 | 49 |
| 22L/91 b | 16,431–16,706 | transcription elongation factor S-II | 10.3 | 22L/91 | 100 | 24L/91 | 98.9 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 27L/92 | 56.3 | 88R/92 | 56.3 | 81R/92 | 56.3 | 23L/92 | 55.2 | 85R/92 | 51.7 |
| 23R/385 b | 16,736–17,893 | RNAseIII | 42.2 | 23R/385 | 100 | 25R/385 | 100 | 22R/385 | 100 | 19R/370 | 99.2 | 19R/385 | 99.2 | 28R/372 | 60.7 | 87L/371 | 61.7 | 80L/371 | 61.7 | 24R/372 | 61.4 | 84L/375 | 52.3 |
| 24L/464 | 17,939–19,333 | ATPase-dependent protease | 52.4 | 24L/464 | 100 | 26L/464 | 100 | 23L/464 | 99.8 | 20L/464 | 99.6 | 20L/464 | 99.6 | 29L/558 | 52.3 | 86R/572 | 52.6 | 79R/572 | 52.8 | 25L/645 | 52.8 | 83R/445 | 41.6 |
| 25R/101 | 19,415–19,720 | hypothetical protein | 11.3 | 25R/101 | 100 | 27R/101 | 100 | 24R/101 | 100 | 21R/101 | 100 | 21R/101 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 26R/270 | 19,764–20,576 | putative eIF-2 alpha-like protein | 30.6 | 26R/270 | 100 | 28R/270 | 100 | 25R/270 | 100 | 22R/270 | 98.2 | 22R/270 | 97.8 | 84L/233 | 30.7 | 28R/69 | 31.1 | 26R/76 | 31.4 | 61R/259 | 30.7 | NA/NA | NA |
| 27R/957 b | 20,666–23,539 | tyrosine kinase | 107.2 | 27R/957 | 100 | 29R/957 | 100 | 26R/957 | 100 | 23R/957 | 99.7 | 23R/957 | 99.7 | 83L/837 | 54.6 | 29R/970 | 53.6 | 27R/970 | 53 | 62R/970 | 54.1 | 78L/790 | 45.9 |
| 28R/159 | 23,550–24,029 | DNA-directed RNA polymerase II subunit | 18 | 28R/159 | 100 | 30R/159 | 100 | 27R/159 | 100 | 24R/159 | 100 | 24R/159 | 100 | 82L/175 | 45.1 | 30R/162 | 43.8 | 28R/162 | 43.8 | 63R/169 | 44.1 | 160L/162 | 39.7 |
| 29R/248 | 24,149–24,895 | capsid maturation protease | 27.8 | 29R/248 | 100 | 32R/248 | 100 | 28R/248 | 100 | 25R/248 | 99.2 | 25R/248 | 99.2 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 65R/274 | 27 | 156L/270 | 23.9 |
| 30R/159 | 24,994–25,473 | hypothetical protein | 17.9 | 30R/159 | 98.7 | 33R/159 | 100 | 29R/159 | 100 | 26R/158 | 96.6 | 26R/158 | 96.6 | 81R/98 | 32.1 | 31L/98 | 32.7 | 29L/98 | 33.7 | 66R/161 | 36.3 | 158L/138 | 38.2 |
| 31L/173 | 25,518–26,039 | hypothetical protein | 19.1 | 31L/173 | 100 | 34L/173 | 100 | 30L/173 | 100 | 27L/114 | 89.3 | 27L/173 | 99.4 | 50L/184 | 67.4 | 61R/184 | 67.4 | NA/NA | NA | NA/NA | NA | 157R/174 | 59.2 |
| 32L/240 | 26,072–26,794 | LPXTG-anchored collagen-like adhesin Scl2/SclB | 22.5 | 32L/240 | 100 | 35L/240 | 99.6 | 31L/240 | 99.6 | 28L/240 | 97.5 | 28L/240 | 97.1 | 75L/144 | 36.7 | NA/NA | NA | 65L/54 | 51.1 | 40R/240 | 40.5 | 56R/246 | 35.8 |
| 33L/257 | 26,802–27,575 | LPXTG-anchored collagen-like adhesin Scl2/SclB | 24.8 | 33L/257 | 100 | 36L/257 | 100 | 32L/257 | 99.6 | 29L/257 | 98.8 | 29L/257 | 98.8 | 75L/144 | 35.1 | 38R/91 | 45 | 65L/54 | 55.1 | 39R/183 | 40.8 | 45L/242 | 36.1 |
| 34L/177 | 27,575–28,108 | hypothetical protein | 18.7 | 34L/177 | 100 | 37L/177 | 100 | 33L/177 | 100 | 30L/177 | 93.8 | 30L/177 | 93.2 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 35L/223 b | 28,163–28,834 | hypothetical protein | 25.7 | 35L/223 | 100 | 38L/223 | 100 | 34L/223 | 100 | 31L/223 | 100 | 31L/223 | 100 | 89R/219 | 72.9 | 23L/219 | 72.9 | 21L/219 | 72.9 | 86R/219 | 72.4 | 54R/215 | 68.9 |
| 36R/563 | 28,859–30,550 | hypothetical protein | 63.8 | 36R/563 | 100 | 39R/563 | 100 | 35R/539 | 100 | 32R/539 | 98.7 | 32R/539 | 98.7 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 37R/955 b | 30,673–33,540 | hypothetical protein | 107.5 | 37R/955 | 99.9 | 40R/955 | 99.8 | 36R/955 | 99.6 | 33R/955 | 99.5 | 33R/955 | 99.5 | 88L/975 | 75 | 24R/975 | 75.2 | 22R/973 | 76.1 | 85L/973 | 76 | 52L/968 | 68 |
| 38R/70 | 33,573–33,785 | TM | 7.7 | 38R/70 | 100 | 42R/70 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 99L/70 | 52.5 | 12R/70 | 52.5 | 11R/70 | 54.2 | 96L/70 | 52.5 | 103R/97 | 38.8 |
| 39L/297 b | 33,832–34,725 | hypothetical protein | 33 | 39L/297 | 100 | 44L/297 | 100 | 37L/297 | 100 | 34L/297 | 99.7 | 34L/297 | 99.7 | 98R/297 | 65.3 | 13L/297 | 66 | 12L/297 | 65.7 | 95R/297 | 66.7 | 118R/319 | 59.7 |
| 40L/253 b | 34,770–35,531 | replicating factor | 29.6 | 40L/253 | 100 | 45L/253 | 99.6 | 38L/253 | 99.2 | 35L/253 | 99.6 | 35L/253 | 99.6 | 1R/256 | 61.5 | 1R/256 | 61.5 | 1R/256 | 61.9 | 100R/256 | 62.4 | 116R/258 | 53.5 |
| 41L/171 | 35,589–36,104 | myeloid cell leukemia protein | 18.7 | 41L/171 | 100 | 46L/171 | 100 | 39L/150 | 100 | 36L/150 | 99.3 | 36L/150 | 99.3 | 101R/137 | 30.6 | 105R/137 | 30.8 | 97R/137 | 30.8 | 99R/137 | 30.5 | 115R/152 | 24.5 |
| 42L/226 | 36,107–36,787 | hypothetical protein | 24.7 | 42L/226 | 100 | 47L/226 | 100 | 40L/214 | 100 | 37L/214 | 99.1 | 37L/214 | 99.1 | 100R/228 | 39.6 | 104R/223 | 40.5 | 96R/223 | 39.6 | 98R/228 | 40.5 | 111R/255 | 41 |
| 43L/78 | 36,866–37,102 | TM | 8.7 | 43L/78 | 100 | 48L/78 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 44L/103 | 37,104–37,415 | TM | 11 | 44L/103 | 100 | 49L/103 | 100 | 38L/253 | 100 | 38L/103 | 100 | 38L/103 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 45R/1354b | 37,201–41,265 | DNA-dependent RNA polymerase a subunit | 147.1 | 45R/1354 | 100 | 50R/1354 | 99.9 | 41R/1353 | 99.9 | 39R/1263 | 99.7 | 39R/1263 | 99.7 | 9R/1294 | 64.9 | 9R/1294 | 64.7 | 8R/1293 | 64.8 | 7R/1303 | 64.9 | 104L/1268 | 62.4 |
| 46R/69 | 41,298–41,507 | hypothetical protein | 6.2 | 46R/55 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 47R/136 | 41,525–41,935 | hypothetical protein | 14.9 | 47R/136 | 100 | 51R/136 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 55L/379 | 52.6 | 57R/379 | 51.6 | 55R/379 | 52.4 | NA/NA | NA | NA/NA | NA |
| 48L/401 | 41,582–42,787 | helicase-like protein | 44.6 | 48L/400 | 100 | 52L/401 | 100 | 42L/401 | 100 | 40L/401 | 98.5 | 40L/401 | 98.5 | 54R/431 | 55 | 56L/431 | 55.2 | 55L/431 | 55 | 51R/431 | 54.7 | 152R/412 | 48 |
| 49L/49 | 42,794–42,943 | TM | 5.2 | 49L/49 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 50R/136 | 42,982–43,392 | hypothetical protein | 15.5 | 50R/136 | 100 | 53R/136 | 100 | 43R/136 | 100 | 41R/136 | 98.5 | 41R/136 | 98.5 | 52L/134 | 37.8 | 59R/134 | 38.6 | NA/NA | NA | 49L/134 | 37.8 | 151L/195 | 39.3 |
| 51R/492 b | 43,401–44,879 | Serine/threonine protein kinases | 53.8 | 51R/492 | 100 | 54R/492 | 99.8 | 44R/492 | 99.8 | 42R/492 | 99.2 | 42R/492 | 99.2 | 51L/498 | 45.2 | 60R/498 | 45.6 | 57R/498 | 45.6 | 48L/498 | 46.3 | 150L/508 | 35.4 |
| 52L/173 | 44,939–45,460 | hypothetical protein | 19.6 | 52L/173 | 100 | 56L/173 | 100 | 45L/172 | 100 | 43L/172 | 99.4 | 43L/172 | 99.4 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 33L/160 | 49 | 148R/159 | 40.8 |
| 53R/60 | 45,433–45,615 | hypothetical protein | 6.5 | 53R/60 | 100 | 57R/60 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 54L/71 | 45,634–45,849 | hypothetical protein | 7.9 | 54L/71 | 100 | 58L/71 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 55L/123 | 45,885–46,256 | hypothetical protein | 13.7 | 55L/123 | 100 | 59L/123 | 100 | 46L/123 | 100 | 44L/123 | 99.2 | 44L/123 | 99.2 | 38L/124 | 40 | 77R/124 | 39.2 | 70R/124 | 40 | 35L/122 | 41.1 | NA/NA | NA |
| 56L/79 | 46,264–46,503 | TM | 8.5 | 56L/79 | 100 | 60L/79 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 39L/88 | 64.8 | 76R/88 | 63.4 | 69R/88 | 63.4 | 36L/88 | 63.4 | 143L/79 | 51.9 |
| 57R/216 | 46,591–47,241 | hypothetical protein | 24 | 57R/216 | 100 | 61R/216 | 100 | 47R/216 | 99.5 | 45R/216 | 100 | 45R/216 | 100 | 40R/107 | 41.1 | 75L/88 | 43.1 | NA/NA | NA | 37R/234 | 35.9 | 145R/165 | 20.5 |
| 58L/324 | 47,338–48,312 | NTPase helicase-like protein | 36.1 | 58L/324 | 100 | 63L/324 | 100 | 48L/324 | 100 | 46L/324 | 99.7 | 46L/324 | 99.7 | 35R/350 | 44.3 | 80L/324 | 44.6 | 73L/324 | 44.6 | 31R/324 | 44.9 | 146L/324 | 47.2 |
| 59L/405 | 48,433–49,650 | hypothetical protein | 44.6 | 59L/405 | 100 | 65L/405 | 100 | 49L/405 | 100 | 47L/405 | 100 | 47L/405 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | 147L/344 | 24.5 |
| 60L/377 | 49,677–50,810 | hypothetical protein | 41.3 | 60L/377 | 100 | 67L/377 | 100 | 50L/377 | 100 | 48L/377 | 98.4 | 48L/377 | 98.4 | 34R/447 | 41.8 | 81L/364 | 42.7 | 74L/370 | 42.3 | 30R/393 | 64.6 | 137R/461 | 41.1 |
| 61L/85 | 50,859–51,116 | lipopolysaccharide-induced TNF-alpha factor-like protein | 9.5 | 61L/85 | 100 | 68L/85 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 33R/84 | 67.1 | 82L/84 | 66.3 | 75L/84 | 65.9 | NA/NA | NA | 136R/104 | 66.7 |
| 62R/73 | 51,175–51,396 | hypothetical protein | 8.2 | 62R/73 | 100 | 69R/73 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 32L/73 | 60.3 | 83R/73 | 58.9 | 76R/73 | 58.9 | 28L/73 | 60.3 | 119R/83 | 30.4 |
| 63L/208 | 51,446–52,072 | hypothetical protein | 23.7 | 63L/208 | 100 | 70L/208 | 100 | 51L/208 | 100 | 49L/208 | 98.6 | 49L/208 | 98.6 | 30R/212 | 39.1 | 85L/224 | 39.5 | 78L/212 | 39.5 | 26R/255 | 38.6 | 122L/210 | 29.1 |
| 64L/371 | 52,139–53,254 | hypothetical protein | 42.6 | 64L/371 | 100 | 71L/371 | 100 | 52L/371 | 100 | 50L/371 | 99.5 | 50L/371 | 99.2 | 48R/352 | 30.9 | 62L/352 | 32 | 59L/352 | 32 | 45R/352 | 32.3 | 123L/362 | 30.5 |
| 65R/182 | 53,321–53,869 | hypothetical protein | 20.4 | 65R/182 | 100 | 72R/182 | 100 | 53R/182 | 100 | 51R/182 | 98.9 | 51R/182 | 98.9 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | 126R/185 | 44.4 |
| 66R/1004 b | 53,929–56,943 | DNA polymerase | 113.7 | 66R/1004 | 100 | 74R/1004 | 100 | 54R/1004 | 100 | 52R/1004 | 99.8 | 52R/1004 | 99.8 | 47L/1013 | 75.8 | 63R/1013 | 75.9 | 60R/1013 | 75.8 | 44L/1013 | 75.6 | 128R/109 | 68.8 |
| 67L/243 | 57,000–57,731 | LPXTG-anchored collagen-like adhesin Scl2/SclB | 23.4 | 67L/243 | 100 | 75L/243 | 100 | 55L/238 | 100 | 53L/238 | 97.5 | 53L/238 | 97.5 | 75L/144 | 42.9 | 38R/91 | 55.6 | NA/NA | NA | 47L/112 | 46.4 | 55R/240 | 59.6 |
| 68L/288 | 57,737–58,603 | LPXTG-anchored collagen-like adhesin Scl2/SclB | 28.9 | 68L/288 | 99.7 | 76L/288 | 100 | 56L/288 | 100 | 54L/288 | 96.5 | 54L/173 | 89.5 | 75L/144 | 40 | 38R/91 | 56.3 | NA/NA | NA | 39R/183 | 58.8 | 112R/355 | 53.1 |
| 69L/387 b | 58,652–59,815 | ribonucleotide reductase beta subunit | 44.1 | 69L/387 | 100 | 77L/387 | 100 | 57L/387 | 100 | 55L/387 | 100 | 55L/387 | 100 | 42R/387 | 77.9 | 73L/387 | 77.7 | 67L/387 | 77.7 | 38R/387 | 77.7 | 47L/384 | 74.9 |
| 70L/91 | 59,932–60,207 | caspase recruitment domain protein | 10.2 | 70L/91 | 100 | 78L/91 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 43L/95 | 39.8 | 68R/95 | 42.1 | 64R/95 | 42.1 | 41L/95 | 42.1 | 48L/91 | 37.7 |
| 71L/141 | 60,269–60,694 | deoxyuridine 5’-triphosphate nucleotidohydrolase | 15.1 | 71L/141 | 99.3 | 80L/141 | 100 | 58L/141 | 100 | 56L/141 | 100 | 56L/141 | 100 | 44L/164 | 57.3 | 67R/164 | 57.3 | 63R/164 | 57.3 | 42L/164 | 57.3 | 49L/155 | 57.9 |
| 72L/237 | 60,795–61,508 | tumor necrosis factor receptor, TM | 25.4 | 72L/237 | 100 | 81L/237 | 100 | 59L/237 | 100 | 57L/237 | 99.6 | 57L/237 | 99.6 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | 51L/231 | 36.7 |
| 73L/178 | 61,588–62,124 | hypothetical protein | 20 | 73L/178 | 100 | 83L/178 | 100 | 60L/178 | 100 | 58L/178 | 99.4 | 58L/178 | 99.4 | 45L/178 | 41.5 | 66R/178 | 40.6 | NA/NA | NA | NA/NA | NA | 75R/178 | 38.9 |
| 74R/1094 b | 62,188–65,472 | DNA-dependent RNA polymerase II | 120.8 | 74R/1094 | 100 | 84R/1094 | 100 | 61R/1094 | 100 | 59R/1094 | 99.6 | 59R/1094 | 99.6 | 46R/1221 | 70.4 | 65L/1221 | 69.9 | 62L/1221 | 69.9 | 43R/1227 | 70 | 73L/1103 | 65.8 |
| 75L/356 b | 65,516–66,586 | DNA repair enzyme RAD2 | 40 | 75L/356 | 100 | 85L/356 | 100 | 62L/356 | 100 | 60L/356 | 98.9 | 60L/356 | 98.9 | 12L/363 | 59.9 | 102R/363 | 59.9 | 95R/363 | 59.9 | 10L/364 | 59.5 | 97L/382 | 59.4 |
| 76R/154 b | 66,655–67,119 | hypothetical protein | 17.7 | 76R/154 | 100 | 86R/154 | 100 | 63R/154 | 99.4 | 61R/154 | 100 | 61R/154 | 100 | 13R/155 | 76.6 | 101L/155 | 76.6 | 94L/155 | 76.6 | 11R/155 | 76.6 | 98R/267 | 67.6 |
| 77L/284 | 67,653–68,507 | DNA polymerase III subunits gamma and tau | 29.1 | 77L/284 | 100 | 89L/284 | 100 | 64L/284 | 99.7 | 62L/263 | 91.1 | 62L/263 | 91.4 | 67R/290 | 33 | 45L/383 | 37.3 | 42L/85 | 34.7 | 78L/285 | 33 | 20L/322 | 35.9 |
| 78L/136 | 68,563–68,973 | hypothetical protein | 15.6 | 78L/136 | 100 | 9L/136 | 100 | 65L/136 | 100 | 63L/136 | 98.5 | 63L/136 | 98.5 | 66R/136 | 62.9 | 46L/136 | 62.1 | 45L/136 | 62.1 | NA/NA | NA | 21L/139 | 60.3 |
| 79L/287 | 69,019–69,882 | neurofilament triplet H1-like protein | 30.8 | 79L/197 | 86 | 92L/191 | 86 | 66L/136 | 86 | 64L/311 | 97 | 64L/239 | 82.6 | 65R/169 | 53.4 | 47L/144 | 52.9 | 46L/81 | 37.1 | 80L/203 | 55.3 | 22L/166 | 44.3 |
| 80L/136 | 69,916–70,326 | hypothetical protein | 16 | 80L/136 | 99.3 | 93L/136 | 98.5 | 67L/136 | 98.5 | 65L/136 | 99.3 | 65L/136 | 99.3 | 64R/138 | 36 | 48L/138 | 38.1 | 47L/138 | 38.1 | 81L/138 | 34.5 | 24L/151 | 27.9 |
| 81L/73 | 70,366–70,587 | hypothetical protein | 8.6 | 81L/73 | 100 | 94L/73 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 82L/566 | 70,628–72,328 | SAP domain-containing protein | 61.7 | 82L/602 | 92.5 | 95L/604 | 93.2 | 68L/320 | 90.4 | 66L/560 | 88.6 | 67L/383 | 95.6 | 62R/508 | 48.8 | 50L/499 | 47.7 | 49L/249 | 49.2 | 83L/541 | 50.8 | 25L/510 | 54.5 |
| 83R/566 | 72,401–74,101 | hypothetical protein | 63.2 | 83R/566 | 100 | 96R/566 | 100 | 70R/566 | 100 | 67R/566 | 99.3 | 68R/566 | 99.1 | 61L/561 | 46.2 | 51R/561 | 46.6 | 51R/561 | 46.6 | 84R/561 | 46.6 | 26R/566 | 39.9 |
| 84L/144 | 74,152–74,586 | hypothetical protein | 15.8 | 84L/144 | 100 | 97L/144 | 100 | 71L/144 | 100 | 68L/144 | 100 | 69L/144 | 100 | 90L/148 | 60.1 | 22R/172 | 60.8 | 20R/148 | 60.1 | 88L/149 | 61.8 | 38L/170 | 44.7 |
| 85L/879 b | 74,614–77,253 | TM | 96.3 | 86R/113 | 99.9 | 99R/113 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | 91L/960 | 41.6 | 21R/852 | 41.8 | 19R/851 | 40.4 | 89L/907 | 41.6 | 39L/1051 | 38.7 |
| 86R/113 | 76,955–77,296 | 2-cysteine adaptor domain protein | 12 | 85L/879 | 100 | 98L/879 | 99.9 | 72L/879 | 99.9 | 69L/879 | 99.2 | 70L/879 | 99.3 | 92R/78 | 42.4 | 20L/79 | 46.2 | 18L/78 | 48.5 | NA/NA | NA | NA/NA | NA |
| 87R/503 | 77,329–78,840 | hypothetical protein | 54.3 | 87R/503 | 100 | 100R/503 | 100 | 73R/503 | 100 | 70R/503 | 99.8 | 71R/503 | 99.8 | 93R/502 | 54.5 | 19L/502 | 54.9 | 17L/502 | 55.1 | 90R/502 | 54.7 | 43R/667 | 39.3 |
| 88R/281 | 78,924–79,769 | hypothetical protein | 31 | 88R/281 | 100 | 101R/281 | 100 | 74R/281 | 100 | 71R/281 | 100 | 72R/281 | 100 | 95L/216 | 45.7 | 17R/275 | 46.7 | 16R/275 | 46.1 | 91R/291 | 47.4 | 132R/275 | 39.7 |
| 89L/315 b | 79,982–80,929 | ABC-ATPase | 35.8 | 89L/315 | 100 | 103L/293 | 100 | 75L/293 | 100 | 72L/300 | 99.3 | 73L/300 | 99.3 | 96L/315 | 80.9 | 16R/315 | 80.9 | 15R/322 | 80.9 | 92L/308 | 80.9 | 134L/323 | 71.2 |
| 90L/119 | 80,896–81,255 | hypothetical protein | 13.7 | 90L/119 | 100 | 104L/119 | 100 | 76L/119 | 100 | 73L/119 | 99.2 | 74L/119 | 99.2 | 97L/119 | 52.1 | 15R/84 | 60.8 | 14R/119 | 52.1 | 93L/118 | 52.1 | 135L/112 | 46.7 |
| 91L/1162 b | 81,325–84,813 | TM | 129.7 | 91L/1162 | 100 | 105L/1162 | 100 | 77L/1162 | 99.9 | 74L/1162 | 99.8 | 75L/1162 | 99.8 | 68L/1165 | 60 | 44R/1165 | 60.1 | 41R/1165 | 59.8 | 77R/1165 | 60 | 57L/1168 | 52 |
| 92L/91 | 84,867–85,142 | TM | 10.2 | 92L/91 | 100 | 107L/91 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | 40R/128 | 44 | 75R/268 | 44 | NA/NA | NA |
| 93L/106 | 85,201–85,521 | hypothetical protein | 12.3 | 93L/106 | 100 | 108L/106 | 100 | 78L/101 | 100 | 75L/106 | 100 | 76L/106 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA |
| 94L/562 | 85,585–87,273 | ribonucleotide reductase alpha subunit | 62.9 | 94L/562 | 100 | 109L/562 | 99.8 | 79L/562 | 100 | 76L/562 | 99.6 | 77L/562 | 99.6 | 71L/565 | 78.8 | 41R/565 | 78.5 | 38R/565 | 78.1 | 73R/254 | 78.9 | 64R/572 | 70.3 |
| 95L/79 | 87,364–87,603 | insulin-like growth factor | 8.4 | 95L/79 | 100 | 110L/79 | 100 | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | NA/NA | NA | 62R/256 | 38.7 |
| 96L/205 b | 87,643–88,260 | NIF/NLI interacting factor | 23.4 | 96L/205 | 100 | 111L/205 | 100 | 80L/205 | 100 | 77L/205 | 99.5 | 78L/205 | 99.5 | 72L/213 | 59.2 | 40R/213 | 59.2 | 37R/209 | 59.2 | 72R/211 | 59.2 | 61R/204 | 50.5 |
| 97L/949 b | 88,278–91,127 | NTPase | 106.4 | 97L/949 | 100 | 112L/949 | 100 | 81L/949 | 100 | 78L/949 | 99.4 | 79L/949 | 99.4 | 10L/948 | 68.2 | 10L/948 | 68.2 | 9L/948 | 68.2 | 8L/948 | 68.1 | 60R/970 | 60.6 |
| 98R/132 | 91,149–91,547 | TM | 14.6 | 98R/132 | 100 | 113R/132 | 100 | 82R/132 | 100 | 79R/132 | 100 | 80R/132 | 100 | 11R/137 | 46 | 11R/137 | 43.1 | 10R/137 | 43.1 | 9R/137 | 47.5 | 59L/146 | 32.4 |
| 99L/189 b | 91,617–92,186 | deoxyribonucleoside kinase | 20.8 | 99L/189 | 100 | 115L/189 | 100 | 83L/189 | 100 | 80L/189 | 99.5 | 81L/189 | 99.5 | 22L/195 | 52.7 | 92R/195 | 52.2 | 85R/195 | 52.2 | 18L/195 | 55.2 | 67L/191 | 53.6 |
| 100L/242 b | 92,238–92,966 | hypothetical protein | 25.8 | 100L/242 | 100 | 116L/242 | 100 | 84L/242 | 100 | 81L/242 | 99.2 | 82L/242 | 99.2 | 23L/245 | 60.5 | 91R/245 | 61.3 | 84R/245 | 60.9 | 19L/260 | 60.9 | 68L/272 | 50 |
| 101L/594 | 93,049–94,833 | hypothetical protein | 66.4 | 101L/594 | 99.8 | 117L/594 | 100 | 85L/594 | 100 | 82L/594 | 99.5 | 83L/594 | 99.5 | 20R/605 | 34.9 | 94L/593 | 33.6 | 87L/605 | 34.9 | 17R/617 | 34.6 | 69L/548 | 32 |
| 102R/147 b | 94,867–95,310 | thiol oxidoreductase | 16.6 | 102R/147 | 100 | 118R/147 | 100 | 86R/147 | 100 | 83R/147 | 99.3 | 84R/147 | 99.3 | 19L/150 | 62.3 | 95R/150 | 61.6 | 88R/150 | 61.6 | 16L/150 | 61 | 70R/152 | 54.6 |
| 103R/413 | 95,316–96,557 | hypothetical protein | 48.5 | 103R/398 | 96.4 | 119R/368 | 88.9 | 1R/387 | 93.7 | 84R/390 | 81.8 | 85R/390 | 83.8 | 18L/414 | 36 | 96R/381 | 37.4 | 89R/388 | 38.1 | 15L/368 | 37.1 | 71R/274 | 38.8 |
| 104R/463 b | 96,670–98,061 | major capsid protein | 50.1 | 104R/463 | 100 | 120R/463 | 100 | 2R/463 | 100 | 85R/463 | 99.4 | 86R/463 | 99.4 | 17L/463 | 84 | 97R/463 | 83.6 | 90R/463 | 83.2 | 14L/463 | 83.4 | 72R/463 | 73.7 |
| 105R/382 b | 98,174–99,322 | immediate early protein ICP-46 | 43.9 | 105R/382 | 100 | 121R/382 | 100 | 3R/382 | 100 | 86R/382 | 99.7 | 87R/382 | 99.7 | 16L/395 | 57.1 | 98R/395 | 57.6 | 91R/395 | 57.9 | 13L/395 | 57.6 | 162L/382 | 50.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.-D.; Ke, F.; Zhang, Q.-Y.; Gui, J.-F. Genome Characteristics of Two Ranavirus Isolates from Mandarin Fish and Largemouth Bass. Pathogens 2023, 12, 730. https://doi.org/10.3390/pathogens12050730
Yu X-D, Ke F, Zhang Q-Y, Gui J-F. Genome Characteristics of Two Ranavirus Isolates from Mandarin Fish and Largemouth Bass. Pathogens. 2023; 12(5):730. https://doi.org/10.3390/pathogens12050730
Chicago/Turabian StyleYu, Xue-Dong, Fei Ke, Qi-Ya Zhang, and Jian-Fang Gui. 2023. "Genome Characteristics of Two Ranavirus Isolates from Mandarin Fish and Largemouth Bass" Pathogens 12, no. 5: 730. https://doi.org/10.3390/pathogens12050730