
Neutralization Sensitivity and Evolution of Virus in a Chronic HIV-1 Clade B Infected Patient with Neutralizing Activity against Membrane-Proximal External Region
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Source and Background
2.2. Viral RNA Extraction, cDNA Synthesis, and SGA
2.3. Functional Plasmid Cloning and Screening
2.4. Pseudovirus Preparation and Titration
2.5. Neutralization Assays
2.6. DNA Sequencing, Alignment, and Analysis
2.7. Amino Acid Analysis
2.8. Simulation of MPER-Antibody Interactions
2.9. Cells
3. Results
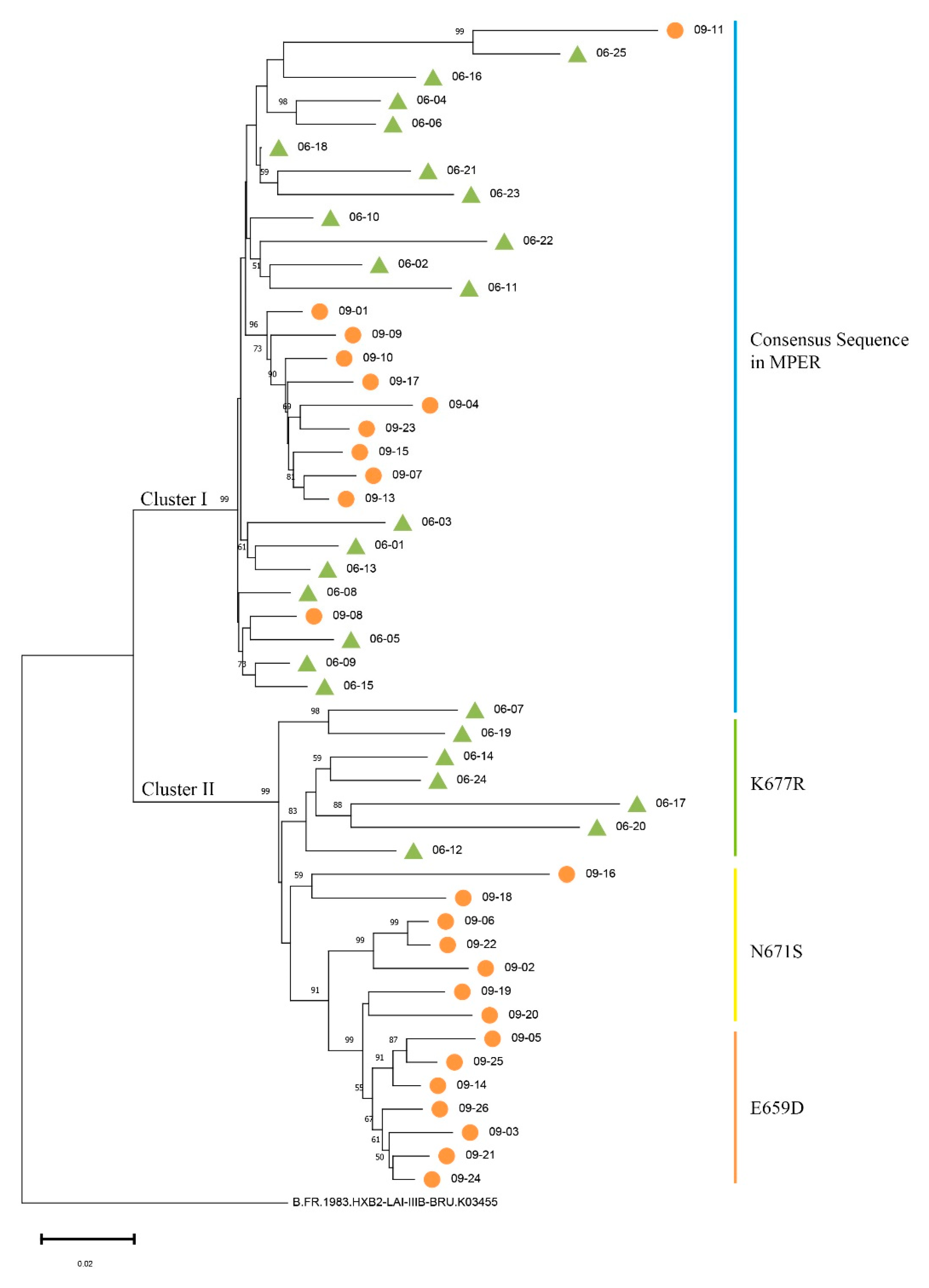
3.1. Analyses of the SGA-Derived Env Genes
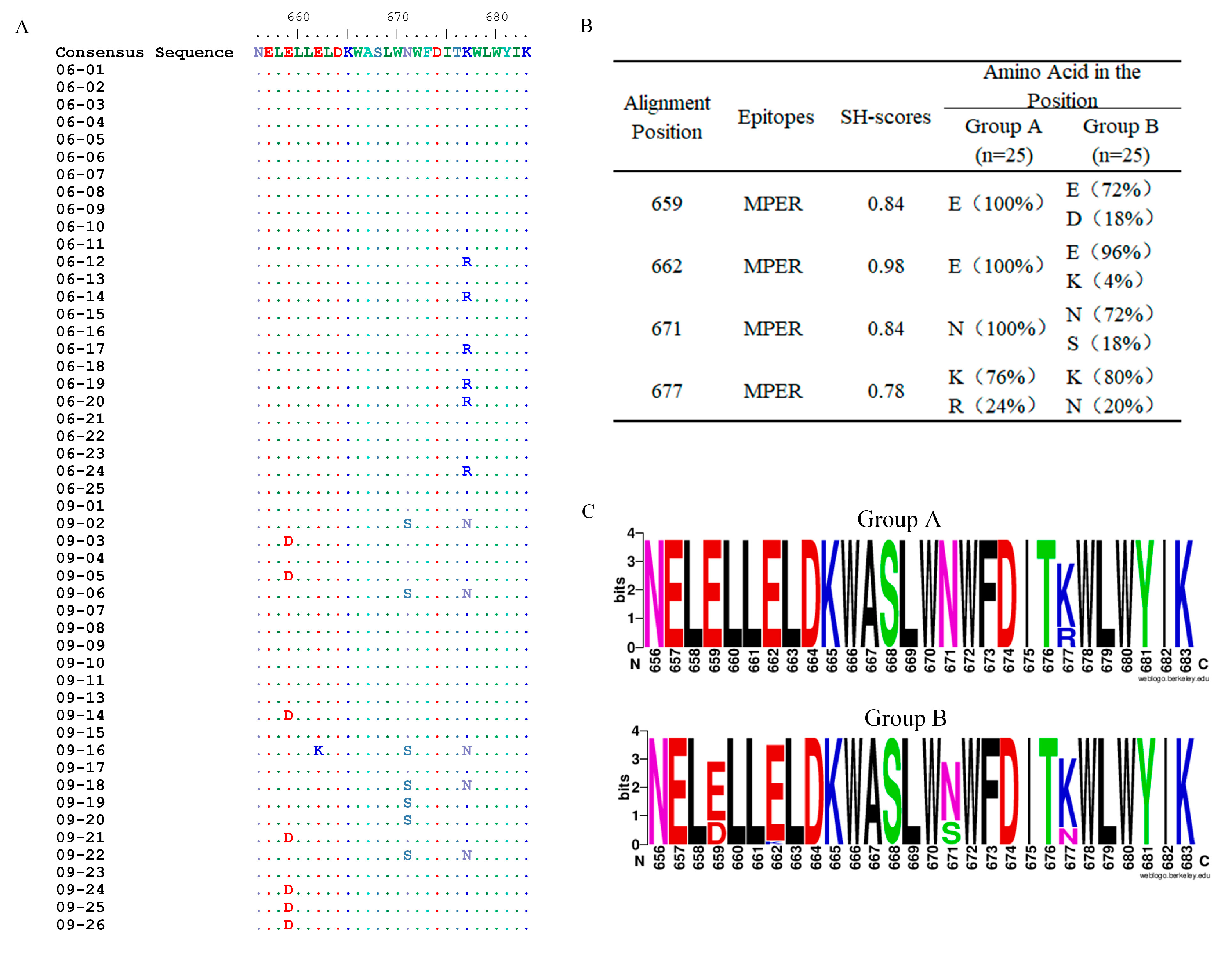
3.2. Amino Acid Mutations in the MPER
3.3. Mutations of 659D and 677R Decreased Neutralization Sensitivity of Pseudovirus to mAbs
3.4. Sensitivity of Pseudoviruses to Autologous Plasma
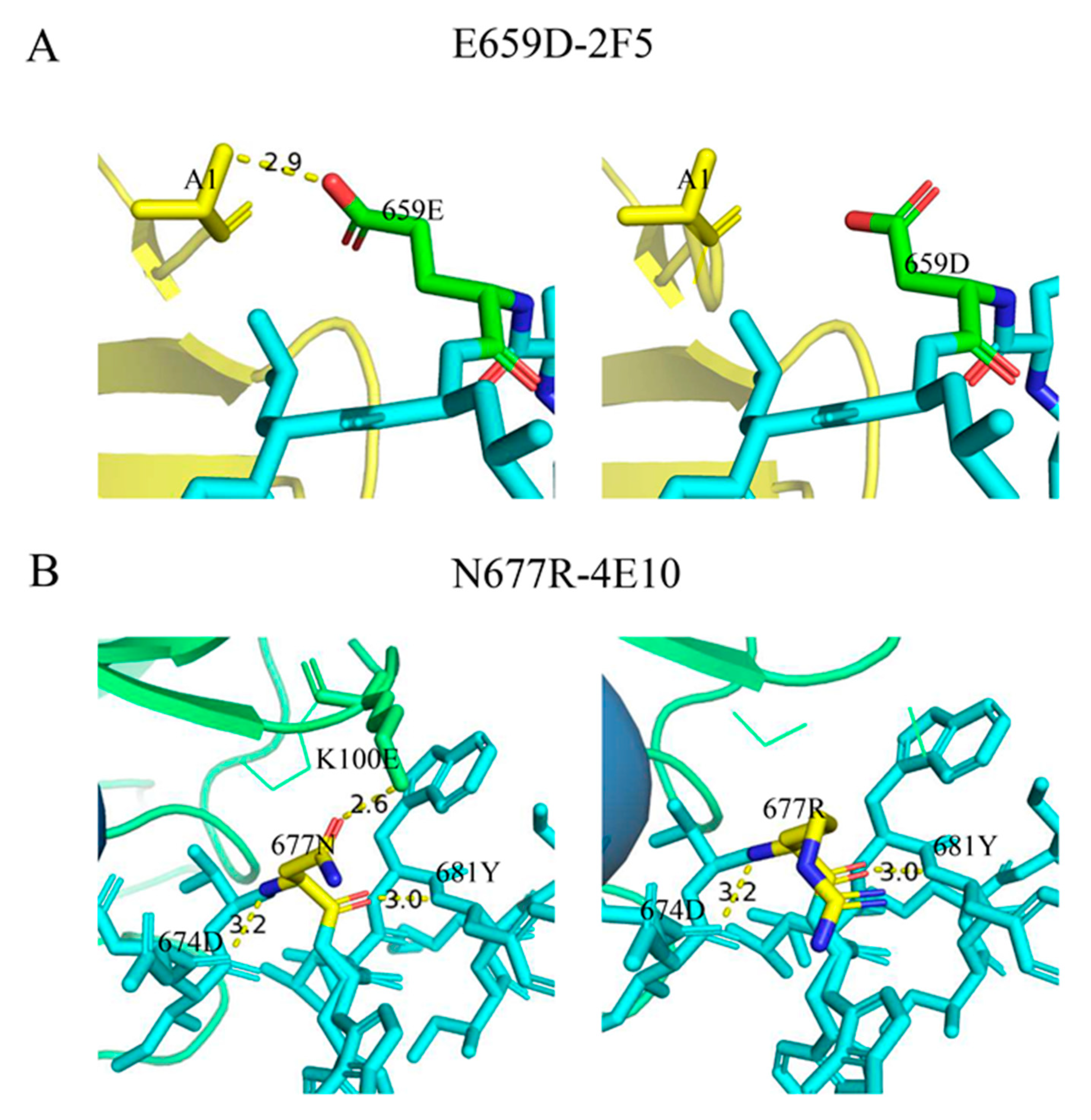
3.5. Effects of Mutations E659D and N677R on the Binding of the MPER Epitope to Antibodies in Simulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Julg, B.; Barouch, D.H. Neutralizing antibodies for HIV-1 prevention. Curr. Opin. HIV AIDS 2019, 14, 318–324. [Google Scholar] [CrossRef]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockerham, L.R.; Hatano, H. Elite control of HIV: Is this the right model for a functional cure? Trends Microbiol. 2015, 23, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Y.J.; Oh, K.J.; Kim, M.; Yu, J.; Brusic, V.; Song, L.; Qiao, Z.; Wang, J.; Wagner, G.; Reinherz, E.L. HIV-1 Broadly Neutralizing Antibody Extracts Its Epitope from a Kinked gp41 Ectodomain Region on the Viral Membrane. Immunity 2008, 28, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Bonsignori, M.; Zhou, T.; Sheng, Z.; Chen, L.; Gao, F.; Joyce, M.G.; Ozorowski, G.; Chuang, G.Y.; Schramm, C.A.; Wiehe, K.; et al. Maturation Pathway from Germline to Broad HIV-1 Neutralizer of a CD4-Mimic Antibody. Cell 2016, 165, 449–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Zhang, D.; Qi, Z.; Hu, X.; Hu, Y.; Ren, L.; Liu, Y.; Ma, L.; Shao, Y.; Hong, K. Characteristics of HIV-1 Env Genes from Chinese Chronically Infected Donors with Highly Broad Cross-Neutralizing Activity. Virology 2020, 551, 16–25. [Google Scholar] [CrossRef]
- Zwick, M.B.; Labrijn, A.F.; Wang, M.; Spenlehauer, C.; Saphire, E.O.; Binley, J.M.; Moore, J.P.; Stiegler, G.; Katinger, H.; Burton, D.R.; et al. Broadly neutralizing antibodies targeted to the membrane-proximal external region of human immunodeficiency virus type 1 glycoprotein gp41. J. Virol. 2001, 75, 10892–10905. [Google Scholar] [CrossRef] [Green Version]
- Hessell, A.J.; Rakasz, E.G.; Tehrani, D.M.; Huber, M.; Weisgrau, K.L.; Landucci, G.; Forthal, D.N.; Koff, W.C.; Poignard, P.; Watkins, D.I.; et al. Broadly neutralizing monoclonal antibodies 2F5 and 4E10 directed against the human immunodeficiency virus type 1 gp41 membrane-proximal external region protect against mucosal challenge by simian-human immunodeficiency virus SHIVBa-L. J. Virol. 2010, 84, 1302–1313. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Ofek, G.; Laub, L.; Louder, M.K.; Doria-Rose, N.A.; Longo, N.S.; Imamichi, H.; Bailer, R.T.; Chakrabarti, B.; Connors, M.; et al. Broad and potent neutralization of HIV-1 by a gp41-specific human antibody. Nature 2012, 491, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Ofek, G.; Zirkle, B.; Yang, Y.; Zhu, Z.; McKee, K.; Zhang, B.; Chuang, G.Y.; Georgiev, I.S.; O’Dell, S.; Kwong, P.D.; et al. Structural basis for HIV-1 neutralization by 2F5-like antibodies m66 and m66.6. J. Virol. 2014, 88, 2426–2441. [Google Scholar] [CrossRef] [Green Version]
- Binley, J.M.; Wrin, T.; Korber, B.; Zwick, M.B.; Wang, M.; Chappey, C.; Stiegler, G.; Kunert, R.; Zolla-Pazner, S.; Katinger, H.; et al. Comprehensive cross-clade neutralization analysis of a panel of anti-human immunodeficiency virus type 1 monoclonal antibodies. J. Virol. 2004, 78, 13232–13252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnett, K.; Barton, F.H. Host controls of HIV broadly neutralizing antibody development. Immunol. Rev. 2017, 275, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Verkoczy, L.; Diaz, M. Autoreactivity in HIV-1 broadly neutralizing antibodies: Implications for their function and induction by vaccination. Curr. Opin. HIV AIDS 2014, 9, 224–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Holl, T.M.; Liu, Y.; Li, Y.; Lu, X.; Nicely, N.I.; Kepler, T.B.; Alam, S.M.; Liao, H.X.; Cain, D.W.; et al. Identification of autoantigens recognized by the 2F5 and 4E10 broadly neutralizing HIV-1 antibodies. J. Exp. Med. 2013, 210, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Su, X.; Si, L.; Lu, L.; Jiang, S. The development of HIV vaccines targeting gp41 membrane-proximal external region (MPER): Challenges and prospects. Protein Cell 2018, 9, 596–615. [Google Scholar] [CrossRef]
- Salzwedel, K.; West, J.T.; Hunter, E. A conserved tryptophan-rich motif in the membrane-proximal region of the human immunodeficiency virus type 1 gp41 ectodomain is important for Env-mediated fusion and virus infectivity. J. Virol. 1999, 73, 2469–2480. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wang, C.; O’Dell, S.; Li, Y.; Keele, B.F.; Yang, Z.; Imamichi, H.; Doria-Rose, N.; Hoxie, J.A.; Connors, M.; et al. Selection pressure on HIV-1 envelope by broadly neutralizing antibodies to the conserved CD4-binding site. J. Virol. 2012, 86, 5844–5856. [Google Scholar] [CrossRef] [Green Version]
- Gray, E.S.; Moore, P.L.; Choge, I.A.; Decker, J.M.; Bibollet-Ruche, F.; Li, H.; Leseka, N.; Treurnicht, F.; Mlisana, K.; Shaw, G.M.; et al. Neutralizing antibody responses in acute human immunodeficiency virus type 1 subtype C infection. J. Virol. 2007, 81, 6187–6196. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Zou, S.; Hu, Y.; Hou, J.; Hu, X.; Ren, L.; Ma, L.; He, X.; Shao, Y.; Hong, K. Characteristics of Envelope Genes in a Chinese Chronically HIV-1 Infected Patient With Broadly Neutralizing Activity. Front. Microbiol. 2019, 10, 1096. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Gonzalez, J.F.; Bailes, E.; Pham, K.T.; Salazar, M.G.; Guffey, M.B.; Keele, B.F.; Derdeyn, C.A.; Farmer, P.; Hunter, E.; Allen, S.; et al. Deciphering human immunodeficiency virus type 1 transmission and early envelope diversification by single-genome amplification and sequencing. J. Virol. 2008, 82, 3952–3970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Zou, S.; Wang, Z.; Liu, Y.; Ren, L.; Hao, Y.; Sun, S.; Hu, X.; Ruan, Y.; Ma, L.; et al. Virus Evolution and Neutralization Sensitivity in an HIV-1 Subtype B’ Infected Plasma Donor with Broadly Neutralizing Activity. Vaccines 2021, 9, 311. [Google Scholar] [CrossRef] [PubMed]
- David, C.M. Measuring HIV neutralization in a luciferase reporter gene assay. Methods Mol. Biol. 2009, 485, 395–405. [Google Scholar] [CrossRef]
- Sarzotti-Kelsoe, M.; Bailer, R.T.; Turk, E.; Lin, C.L.; Bilska, M.; Greene, K.M.; Gao, H.; Todd, C.A.; Ozaki, D.A.; Seaman, M.S.; et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1. J. Immunol. Methods 2014, 409, 131–146. [Google Scholar] [CrossRef] [Green Version]
- Gaschen, B.; Kuiken, C.; Korber, B.; Foley, B. Retrieval and on-the-fly alignment of sequence fragments from the HIV database. Bioinformatics 2001, 17, 415–418. [Google Scholar] [CrossRef] [Green Version]
- Bricault, C.A.; Yusim, K.; Seaman, M.S.; Yoon, H.; Theiler, J.; Giorgi, E.E.; Wagh, K.; Theiler, M.; Hraber, P.; Macke, J.; et al. HIV-1 Neutralizing Antibody Signatures and Application to Epitope-Targeted Vaccine Design. Cell Host Microbe 2019, 25, 59–72.e8. [Google Scholar] [CrossRef] [Green Version]
- Bradley, T.; Trama, A.; Tumba, N.; Gray, E.; Lu, X.; Madani, N.; Jahanbakhsh, F.; Eaton, A.; Xia, S.M.; Parks, R.; et al. Amino Acid Changes in the HIV-1 gp41 Membrane Proximal Region Control Virus Neutralization Sensitivity. EBioMedicine 2016, 12, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Ofek, G.; Tang, M.; Sambor, A.; Katinger, H.; Mascola, J.R.; Wyatt, R.; Kwong, P.D. Structure and mechanistic analysis of the anti-human immunodeficiency virus type 1 antibody 2F5 in complex with its gp41 epitope. J. Virol. 2004, 78, 10724–10737. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Irimia, A.; He, L.; Landais, E.; Rantalainen, K.; Leaman, D.P.; Vollbrecht, T.; Stano, A.; Sands, D.I.; Zwick, M.B.; et al. An MPER antibody neutralizes HIV-1 using germline features shared among donors. Nat. Commun. 2019, 10, 5389. [Google Scholar] [CrossRef] [Green Version]
- West, A.P., Jr.; Scharf, L.; Scheid, J.F.; Klein, F.; Bjorkman, P.J.; Nussenzweig, M.C. Structural insights on the role of antibodies in HIV-1 vaccine and therapy. Cell 2014, 156, 633–648. [Google Scholar] [CrossRef] [Green Version]
- Schapiro, H.M.; Khasnis, M.D.; Ahn, K.; Karagiaridi, A.; Hayden, S.; Cilento, M.E.; Root, M.J. Regulation of epitope exposure in the gp41 membrane-proximal external region through interactions at the apex of HIV-1 Env. PLoS Pathog. 2022, 18, e1010531. [Google Scholar] [CrossRef]
- West, A.P., Jr.; Scharf, L.; Horwitz, J.; Klein, F.; Nussenzweig, M.C.; Bjorkman, P.J. Computational analysis of anti-HIV-1 antibody neutralization panel data to identify potential functional epitope residues. Proc. Natl. Acad. Sci. USA 2013, 110, 10598–10603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.X.; Lynch, R.; Zhou, T.; Gao, F.; Alam, S.M.; Boyd, S.D.; Fire, A.Z.; Roskin, K.M.; Schramm, C.A.; Zhang, Z.; et al. Co-evolution of a broadly neutralizing HIV-1 antibody and founder virus. Nature 2013, 496, 469–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunnik, E.M.; Pisas, L.; Van Nuenen, A.C.; Schuitemaker, H. Autologous neutralizing humoral immunity and evolution of the viral envelope in the course of subtype b human immunodeficiency virus type 1 infection. J. Virol. 2008, 82, 7932–7941. [Google Scholar] [CrossRef] [Green Version]
- Irimia, A.; Sarkar, A.; Stanfield, R.L.; Wilson, I.A. Crystallographic Identification of Lipid as an Integral Component of the Epitope of HIV Broadly Neutralizing Antibody 4E10. Immunity 2016, 44, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, R.; Zwick, M.B.; Stanfield, R.L.; Kunert, R.; Binley, J.M.; Katinger, H.; Burton, D.R.; Wilson, I.A. Broadly Neutralizing Anti-HIV Antibody 4E10 Recognizes a Helical Conformation of a Highly Conserved Fusion-Associated Motif in gp41. Immunity 2005, 22, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.P.; Hock, M.; Radzimanowski, J.; Tonks, P.; Hulsik, D.L.; Effantin, G.; Seilly, D.J.; Dreja, H.; Kliche, A.; Wagner, R.; et al. A fusion intermediate gp41 immunogen elicits neutralizing antibodies to HIV-1. J. Biol. Chem. 2014, 289, 29912–29926. [Google Scholar] [CrossRef] [Green Version]
- McGaughey, G.B.; Citron, M.; Danzeisen, R.C.; Freidinger, R.M.; Garsky, V.M.; Hurni, W.M.; Joyce, J.G.; Liang, X.; Miller, M.; Shiver, J.; et al. HIV-1 vaccine development: Constrained peptide immunogens show improved binding to the anti-HIV-1 gp41 MAb. Biochemistry 2003, 42, 3214–3223. [Google Scholar] [CrossRef]
- Chuang, G.Y.; Liou, D.; Kwong, P.D.; Georgiev, I.S. NEP: Web Server for Epitope Prediction Based on Antibody Neutralization of Viral Strains with Diverse Sequences. Nucleic Acids Res. 2014, 42, W64–W71. [Google Scholar] [CrossRef] [Green Version]
- Freund, N.T.; Wang, H.; Scharf, L.; Nogueira, L.; Horwitz, J.A.; Bar-On, Y.; Golijanin, J.; Sievers, S.A.; Sok, D.; Nussenzweig, M.C.; et al. Coexistence of potent HIV-1 broadly neutralizing antibodies and antibody-sensitive viruses in a viremic controller. Sci. Transl. Med. 2017, 9, eaal2144. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pseudoviruses ID | Mutation Positions in MPER | IC50 (μg/mL) | ||
|---|---|---|---|---|
| 4E10 | 2F5 | 10E8 | ||
| 06–01 | None | 0.60 | 0.88 | 0.08 |
| 06–02 | 1.12 | 0.88 | 0.11 | |
| 06–07 | 2.79 | 3.85 | 0.36 | |
| 06–13 | 0.35 | 0.47 | 0.05 | |
| 09–04 | 0.44 | 0.37 | 0.05 | |
| 09–07 | 0.39 | 0.50 | 0.05 | |
| 09–08 | 0.37 | 0.50 | 0.07 | |
| Mean ± SD | 0.87 ± 0.89 | 1.06 ± 1.25 | 0.11 ± 0.11 | |
| 06–12 | 677R | 3.89 | 1.62 | 0.17 |
| 06–14 | 0.25 | 1.32 | 0.16 | |
| 06–20 | 2.70 | 1.35 | 0.13 | |
| 06–24 | 1.72 | 2.53 | 0.26 | |
| Mean ± SD | 2.14 ± 1.54 | 1.70 ± 0.57 | 0.18 ± 0.06 | |
| 09–05 | 659D | 7.87 | 4.76 | 0.14 |
| 09–21 | 2.11 | 0.97 | 0.14 | |
| 09–22 | 671S, 677N | 0.74 | 0.84 | 0.03 |
| Pseudoviruses ID | Mutation Positions in MPER | ID50 of Autologous Plasmas against Pseudoviruses | |||||
|---|---|---|---|---|---|---|---|
| 2005.7 | 2006.4 | 2007.4 | 2008.11 | 2009.6 | 2010.11 | ||
| 06–01 | None | 115 | 205 | 847 | 886 | 707 | 611 |
| 06–02 | 125 | 158 | 1480 | 1399 | 424 | 498 | |
| 06–07 | 521 | 559 | 696 | 1455 | 1335 | 1116 | |
| 06–13 | 137 | 179 | 818 | 871 | 380 | 674 | |
| 06–12 | 677R | <20 | <20 | 84 | 889 | 1131 | 597 |
| 06–14 | 60 | 304 | 598 | 727 | 1294 | 644 | |
| 06–20 | <20 | <20 | 80 | 880 | 1082 | 572 | |
| 06–24 | <20 | <20 | 150 | 411 | 400 | 401 | |
| GEOMEAN (n = 8) | 53 | 75 | 379 | 882 | 746 | 614 | |
| 09–04 | None | 28 | 49 | 39 | 22 | 40 | 1338 |
| 09–07 | 28 | 49 | 54 | 23 | 37 | 835 | |
| 09–08 | 51 | 67 | 79 | 42 | 76 | 898 | |
| 09–05 | 659D | <20 | <20 | <20 | 24 | <20 | 312 |
| 09–21 | 25 | 27 | 43 | 68 | 127 | 265 | |
| 09–22 | 671S, 677N | <20 | <20 | <20 | 34 | <20 | 223 |
| GEOMEAN (n = 6) | 22 | 28 | 30 | 33 | 34 | 514 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, W.; Yuan, Z.; Wang, Z.; Ren, L.; Li, D.; Wang, S.; Hao, Y.; Li, J.; Shen, X.; Ruan, Y.; et al. Neutralization Sensitivity and Evolution of Virus in a Chronic HIV-1 Clade B Infected Patient with Neutralizing Activity against Membrane-Proximal External Region. Pathogens 2023, 12, 497. https://doi.org/10.3390/pathogens12030497
Tang W, Yuan Z, Wang Z, Ren L, Li D, Wang S, Hao Y, Li J, Shen X, Ruan Y, et al. Neutralization Sensitivity and Evolution of Virus in a Chronic HIV-1 Clade B Infected Patient with Neutralizing Activity against Membrane-Proximal External Region. Pathogens. 2023; 12(3):497. https://doi.org/10.3390/pathogens12030497
Chicago/Turabian StyleTang, Wenqi, Zhenzhen Yuan, Zheng Wang, Li Ren, Dan Li, Shuhui Wang, Yanling Hao, Jing Li, Xiuli Shen, Yuhua Ruan, and et al. 2023. "Neutralization Sensitivity and Evolution of Virus in a Chronic HIV-1 Clade B Infected Patient with Neutralizing Activity against Membrane-Proximal External Region" Pathogens 12, no. 3: 497. https://doi.org/10.3390/pathogens12030497