
A Systematic Review of Apicomplexa Looking into Epigenetic Pathways and the Opportunity for Novel Therapies

Abstract
:1. Introduction
2. Material and Methods
2.1. Information Sources and Search Strategy
2.2. Search Analysis—Eligibility Criteria
- Research article: only original research articles were included.
- Epigenetic approaches: papers showed one or more tests to demonstrate the epigenetic event.
- Host–parasite interaction: studies investigating modifications due to the interaction between the parasite and the host.
- Host species: only studies in mammalian hosts were included.
- Parasite species: only Apicomplexa parasites were included.
- Time period: articles were included independently of the year of publication.
- Idiom: articles published in any language were considered. For those published in idioms other than Portuguese or English, Google Translate (https://translate.google.com/, accessed on 25 January 2023) was used for translation into English.
2.3. Study Records, Data Items, Outcome, and Bias Risk Evaluation
2.4. Additional Analysis
3. Results and Discussion
3.1. Document Selection
3.2. Analysis of Affiliated Countries
3.3. Keywords Co-Occurrence Analysis
3.4. Epidrugs: Drug Targets and Host-Directed Therapy
3.4.1. Histone Post-Translational Modifications and Chromatin Remodeling
3.4.2. DNA Methylation
3.4.3. ncRNA
4. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vilcinskas, A. The Role of Epigenetics in Host–Parasite Coevolution: Lessons from the Model Host Insects Galleria Mellonella and Tribolium Castaneum. Zoology 2016, 119, 273–280. [Google Scholar] [CrossRef]
- Waddington, C.H. The Epigenotype. Endeavour 1942, 1, 18–20. [Google Scholar] [CrossRef]
- Nicoglou, A.; Merlin, F. Epigenetics: A Way to Bridge the Gap between Biological Fields. Stud. Hist. Philos. Sci. Part C Stud. Hist. Philos. Biol. Biomed. Sci. 2017, 66, 73–82. [Google Scholar] [CrossRef]
- Kumar, S.; Gonzalez, E.A.; Rameshwar, P.; Etchegaray, J.P. Non-Coding RNAs as Mediators of Epigenetic Changes in Malignancies. Cancers 2020, 12, 3657. [Google Scholar] [CrossRef]
- Brandão, Y.d.O.; Toledo, M.B.; Chequin, A.; Cristo, T.G.; Sousa, R.S.; Ramos, E.A.S.; Klassen, G. DNA Methylation Status of the Estrogen Receptor α Gene in Canine Mammary Tumors. Vet. Pathol. 2018, 55, 510–516. [Google Scholar] [CrossRef]
- Pires, V.S.; de, O. Ganzella, F.A.; Minozzo, G.A.; de Castro, L.L.; Moncada, A.D.B.; Klassen, G.; Ramos, E.A.S.; Molento, M.B. Epigenetic Regulation of SLC11a1 Gene in Horses Infected with Cyathostomins. Gene Rep. 2021, 25, 101410. [Google Scholar] [CrossRef]
- Berdasco, M.; Esteller, M. Clinical Epigenetics: Seizing Opportunities for Translation. Nat. Rev. Genet. 2019, 20, 109–127. [Google Scholar] [CrossRef]
- Ganesan, A.; Arimondo, P.B.; Rots, M.G.; Jeronimo, C.; Berdasco, M. The Timeline of Epigenetic Drug Discovery: From Reality to Dreams. Clin. Epigenetics 2019, 11, 174. [Google Scholar] [CrossRef]
- Egea, P.F. Crossing the Vacuolar Rubicon: Structural Insights into Effector Protein Trafficking in Apicomplexan Parasites. Microorganisms 2020, 8, 865. [Google Scholar] [CrossRef]
- Villares, M.; Berthelet, J.; Weitzman, J.B. The Clever Strategies Used by Intracellular Parasites to Hijack Host Gene Expression. Semin. Immunopathol. 2020, 42, 215–226. [Google Scholar] [CrossRef]
- Chen, X.-M.; Splinter, P.L.; O’Hara, S.P.; LaRusso, N.F. A Cellular Micro-RNA, Let-7i, Regulates Toll-like Receptor 4 Expression and Contributes to Cholangiocyte Immune Responses against Cryptosporidium parvum Infection. J. Biol. Chem. 2007, 282, 28929–28938. [Google Scholar] [CrossRef]
- Zhou, R.; Hu, G.; Liu, J.; Gong, A.-Y.; Drescher, K.M.; Chen, X.-M. NF-KappaB P65-Dependent Transactivation of MiRNA Genes Following Cryptosporidium parvum Infection Stimulates Epithelial Cell Immune Responses. PLoS Pathog. 2009, 5, e1000681. [Google Scholar] [CrossRef]
- Gong, A.-Y.; Zhou, R.; Hu, G.; Liu, J.; Sosnowska, D.; Drescher, K.M.; Dong, H.; Chen, X.-M. Cryptosporidium parvum Induces B7-H1 Expression in Cholangiocytes by Down-Regulating MicroRNA-513. J. Infect. Dis. 2010, 201, 160–169. [Google Scholar] [CrossRef]
- O’Hara, S.P.; Splinter, P.L.; Gajdos, G.B.; Trussoni, C.E.; Fernandez-Zapico, M.E.; Chen, X.M.X.-M.; LaRusso, N.F. NFκB P50-CCAAT/Enhancer-Binding Protein β (C/EBPβ)-Mediated Transcriptional Repression of MicroRNA Let-7i Following Microbial Infection. J. Biol. Chem. 2010, 285, 216–225. [Google Scholar] [CrossRef]
- Hu, G.; Zhou, R.; Liu, J.; Gong, A.-Y.; Eischeid, A.N.; Dittman, J.W.; Chen, X.-M. MicroRNA-98 and Let-7 Confer Cholangiocyte Expression of Cytokine-Inducible Src Homology 2-Containing Protein in Response to Microbial Challenge. J. Immunol. 2009, 183, 1617–1624. [Google Scholar] [CrossRef]
- Hu, G.; Zhou, R.; Liu, J.; Gong, A.-Y.; Chen, X.-M. MicroRNA-98 and Let-7 Regulate Expression of Suppressor of Cytokine Signaling 4 in Biliary Epithelial Cells in Response to Cryptosporidium parvum Infection. J. Infect. Dis. 2010, 202, 125–135. [Google Scholar] [CrossRef]
- Gong, A.-Y.; Hu, G.; Zhou, R.; Liu, J.; Feng, Y.; Soukup, G.A.; Chen, X.-M. MicroRNA-221 Controls Expression of Intercellular Adhesion Molecule-1 in Epithelial Cells in Response to Cryptosporidium parvum Infection. Int. J. Parasitol. 2011, 41, 397–403. [Google Scholar] [CrossRef]
- Zhou, R.; Gong, A.-Y.; Eischeid, A.N.; Chen, X.-M. MiR-27b Targets KSRP to Coordinate TLR4-Mediated Epithelial Defense against Cryptosporidium parvum Infection. PLoS Pathog. 2012, 8, e1002702. [Google Scholar] [CrossRef]
- Hu, G.; Gong, A.-Y.; Roth, A.L.; Huang, B.Q.; Ward, H.D.; Zhu, G.; LaRusso, N.F.; Hanson, N.D.; Chen, X.-M. Release of Luminal Exosomes Contributes to TLR4-Mediated Epithelial Antimicrobial Defense. PLoS Pathog. 2013, 9, e1003261. [Google Scholar] [CrossRef]
- Xie, H.; Lei, N.; Gong, A.-Y.; Chen, X.-M.; Hu, G. Cryptosporidium parvum Induces SIRT1 Expression in Host Epithelial Cells through Downregulating Let-7i. Hum. Immunol. 2014, 75, 760–765. [Google Scholar] [CrossRef]
- Guesdon, W.; Auray, G.; Pezier, T.; Bussière, F.I.; Drouet, F.; Le Vern, Y.; Marquis, M.; Potiron, L.; Rabot, S.; Bruneau, A.; et al. CCL20 Displays Antimicrobial Activity Against Cryptosporidium parvum, but Its Expression Is Reduced During Infection in the Intestine of Neonatal Mice. J. Infect. Dis. 2015, 212, 1332–1340. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, A.-Y.; Ma, S.; Chen, X.-M.X.; Strauss-Soukup, J.K.; Chen, X.-M.X. Delivery of Parasite Cdg7_Flc_0990 RNA Transcript into Intestinal Epithelial Cells during Cryptosporidium parvum Infection Suppresses Host Cell Gene Transcription through Epigenetic Mechanisms. Cell. Microbiol. 2017, 19, e12760. [Google Scholar] [CrossRef]
- Ming, Z.; Gong, A.-Y.; Wang, Y.; Zhang, X.-T.; Li, M.; Mathy, N.W.; Strauss-Soukup, J.K.; Chen, X.-M. Involvement of Cryptosporidium parvum Cdg7_FLc_1000 RNA in the Attenuation of Intestinal Epithelial Cell Migration via Trans-Suppression of Host Cell SMPD3. J. Infect. Dis. 2018, 217, 122–133. [Google Scholar] [CrossRef]
- Ming, Z.; Gong, A.-Y.; Wang, Y.; Zhang, X.; Li, M.; Li, Y.; Pang, J.; Dong, S.; Strauss-Soukup, J.K.; Chen, X.-M. Trans-Suppression of Host CDH3 and LOXL4 Genes during Cryptosporidium parvum Infection Involves Nuclear Delivery of Parasite Cdg7_FLc_1000 RNA. Int. J. Parasitol. 2018, 48, 423–431. [Google Scholar] [CrossRef]
- Jainul, M.A.; Isa, M.L.M.; Yusof, A.M.M. staining and mirna expression of H.-8 and H.-29 cell lines upon cryptosporidium infection Immunofluorescent Staining and Mirna Expression of HCT-8 and HT-29 Cell Lines upon Cryptosporidium Infection. Malays. Appl. Biol. 2018, 47, 101–112. [Google Scholar]
- Li, M.; Gong, A.-Y.; Zhang, X.-T.; Wang, Y.; Mathy, N.W.; Martins, G.A.; Strauss-Soukup, J.K.; Chen, X.-M. Induction of a Long Noncoding RNA Transcript, NR_045064, Promotes Defense Gene Transcription and Facilitates Intestinal Epithelial Cell Responses against Cryptosporidium Infection. J. Immunol. 2018, 201, 3630–3640. [Google Scholar] [CrossRef]
- Liu, T.-L.; Fan, X.-C.; Li, Y.-H.; Yuan, Y.-J.; Yin, Y.-L.; Wang, X.-T.; Zhang, L.-X.; Zhao, G.-H. Expression Profiles of MRNA and LncRNA in HCT-8 Cells Infected With Cryptosporidium parvum IId Subtype. Front. Microbiol. 2018, 9, 1409. [Google Scholar] [CrossRef]
- Zhao, G.-H.; Gong, A.-Y.; Wang, Y.; Zhang, X.-T.; Li, M.; Mathy, N.W.; Chen, X.-M. Nuclear Delivery of Parasite Cdg2_FLc_0220 RNA Transcript to Epithelial Cells during Cryptosporidium parvum Infection Modulates Host Gene Transcription. Vet. Parasitol. 2018, 251, 27–33. [Google Scholar] [CrossRef]
- Wang, C.; Liu, L.; Zhu, H.; Zhang, L.; Wang, R.; Zhang, Z.; Huang, J.; Zhang, S.; Jian, F.; Ning, C.; et al. MicroRNA Expression Profile of HCT-8 Cells in the Early Phase of Cryptosporidium parvum Infection. BMC Genom. 2019, 20, 37. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, Y.; Niu, Z.; Wang, C.; Xie, F.; Li, J.; Zhang, S.; Qi, M.; Jian, F.; Ning, C.; et al. Cryptosporidium parvum Upregulates MiR-942-5p Expression in HCT-8 Cells via TLR2/TLR4-NF-ΚB Signaling. Parasites Vectors 2020, 13, 4–9. [Google Scholar] [CrossRef]
- Gong, A.-Y.; Wang, Y.; Li, M.; Zhang, X.-T.; Deng, S.; Chen, J.M.; Lu, E.; Mathy, N.W.; Martins, G.A.; Strauss-Soukup, J.K.; et al. LncRNA XR_001779380 Primes Epithelial Cells for IFN-γ-Mediated Gene Transcription and Facilitates Age-Dependent Intestinal Antimicrobial Defense. MBio 2021, 12, e0212721. [Google Scholar] [CrossRef]
- Sabir, M.J.; Low, R.; Hall, N.; Kamli, M.R.; Malik, M.Z. A Bioinformatics Approach to Identifying Potential Biomarkers for Cryptosporidium parvum: A Coccidian Parasite Associated with Fetal Diarrhea. Vaccines 2021, 9, 1427. [Google Scholar] [CrossRef]
- Yin, Y.-L.; Liu, T.-L.; Yao, Q.; Wang, Y.-X.; Wu, X.-M.; Wang, X.-T.; Yang, X.; Song, J.-K.; Zhao, G.-H. Circular RNA CiRS-7 Affects the Propagation of Cryptosporidium parvum in HCT-8 Cells by Sponging MiR-1270 to Activate the NF-ΚB Signaling Pathway. Parasites Vectors 2021, 14, 238. [Google Scholar] [CrossRef]
- Mathy, N.W.; Deng, S.; Gong, A.-Y.; Li, M.; Wang, Y.; Burleigh, O.; Kochvar, A.; Whiteford, E.R.; Shibata, A.; Chen, X.-M. The Long Non-Coding RNA Nostrill Regulates Transcription of Irf7 Through Interaction With NF-ΚB P65 to Enhance Intestinal Epithelial Defense Against Cryptosporidium parvum. Front. Immunol. 2022, 13, 863957. [Google Scholar] [CrossRef]
- Sawant, M.; Benamrouz-Vanneste, S.; Meloni, D.; Gantois, N.; Even, G.; Guyot, K.; Creusy, C.; Duval, E.; Wintjens, R.; Weitzman, J.B.; et al. Putative SET-Domain Methyltransferases in Cryptosporidium parvum and Histone Methylation during Infection. Virulence 2022, 13, 1632–1650. [Google Scholar] [CrossRef]
- Ulusan Bagci, Ö.; Caner, A.; Ulusan Bağcı, Ö.; Caner, A.; Ulusan Bagci, Ö.; Caner, A.; Bağci, Ö.U.; Caner, A. miRNA Expression Profile in Ileocecal Adenocarcinoma Cells Infected with Cryptosporidium. Mikrobiyoloji Bul. 2022, 56, 449–465. [Google Scholar] [CrossRef]
- Xie, F.; Zhang, Y.; Li, J.; Sun, L.; Zhang, L.; Qi, M.; Zhang, S.; Jian, F.; Li, X.; Li, J.; et al. MiR-942-5p Targeting the IFI27 Gene Regulates HCT-8 Cell Apoptosis via a TRAIL-Dependent Pathway during the Early Phase of Cryptosporidium parvum Infection. Parasites Vectors 2022, 15, 291. [Google Scholar] [CrossRef]
- El-Assaad, F.; Hempel, C.; Combes, V.; Mitchell, A.J.; Ball, H.J.; Kurtzhals, J.A.L.L.; Hunt, N.H.; Mathys, J.-M.M.; Grau, G.E.R.R. Differential MicroRNA Expression in Experimental Cerebral and Noncerebral Malaria. Infect. Immun. 2011, 79, 2379–2384. [Google Scholar] [CrossRef]
- Hentzschel, F.; Hammerschmidt-Kamper, C.; Börner, K.; Heiss, K.; Knapp, B.; Sattler, J.M.; Kaderali, L.; Castoldi, M.; Bindman, J.G.; Malato, Y.; et al. AAV8-Mediated In Vivo Overexpression of MiR-155 Enhances the Protective Capacity of Genetically Attenuated Malarial Parasites. Mol. Ther. 2014, 22, 2130–2141. [Google Scholar] [CrossRef]
- Martin-Alonso, A.; Cohen, A.; Quispe-Ricalde, M.A.; Foronda, P.; Benito, A.; Berzosa, P.; Valladares, B.; Grau, G.E. Differentially Expressed MicroRNAs in Experimental Cerebral Malaria and Their Involvement in Endocytosis, Adherens Junctions, FoxO and TGF-β Signalling Pathways. Sci. Rep. 2018, 8, 11277. [Google Scholar] [CrossRef]
- Delic, D.; Dkhil, M.; Al-Quraishy, S.; Wunderlich, F.; Delić, D.; Dkhil, M.; Al-Quraishy, S.; Wunderlich, F. Hepatic MiRNA Expression Reprogrammed by Plasmodium Chabaudi Malaria. Parasitol. Res. 2011, 108, 1111–1121. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Dkhil, M.A.; Delic, D.; Abdel-Baki, A.A.; Wunderlich, F. Organ-Specific Testosterone-Insensitive Response of MiRNA Expression of C57BL/6 Mice to Plasmodium Chabaudi Malaria. Parasitol. Res. 2012, 111, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Al-Quraishy, S.; Dkhil, M.A.; Abdel-Baki, A.A.S.; Delic, D.; Santourlidis, S.; Wunderlich, F. Genome-Wide Screening Identifies Plasmodium Chabaudi-Induced Modifications of DNA Methylation Status of Tlr1 and Tlr6 Gene Promoters in Liver, but Not Spleen, of Female C57BL/6 Mice. Parasitol. Res. 2013, 112, 3757–3770. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Dkhil, M.A.; Abdel-Baki, A.-A.S.; Ghanjati, F.; Erichsen, L.; Santourlidis, S.; Wunderlich, F.; Araúzo-Bravo, M.J. Protective Vaccination and Blood-Stage Malaria Modify DNA Methylation of Gene Promoters in the Liver of Balb/c Mice. Parasitol. Res. 2017, 116, 1463–1477. [Google Scholar] [CrossRef]
- Nahrendorf, W.; Ivens, A.; Spence, P.J. Inducible Mechanisms of Disease Tolerance Provide an Alternative Strategy of Acquired Immunity to Malaria. Elife 2021, 10, e63838. [Google Scholar] [CrossRef] [PubMed]
- Rathjen, T.; Nicol, C.; McConkey, G.; Dalmay, T. Analysis of Short RNAs in the Malaria Parasite and Its Red Blood Cell Host. FEBS Lett. 2006, 580, 5185–5188. [Google Scholar] [CrossRef]
- LaMonte, G.; Philip, N.; Reardon, J.; Lacsina, J.R.R.; Majoros, W.; Chapman, L.; Thornburg, C.D.; Telen, M.J.; Ohler, U.; Nicchitta, C.V.; et al. Translocation of Sickle Cell Erythrocyte MicroRNAs into Plasmodium Falciparum Inhibits Parasite Translation and Contributes to Malaria Resistance. Cell Host Microbe 2012, 12, 187–199. [Google Scholar] [CrossRef]
- Mantel, P.-Y.; Hjelmqvist, D.; Walch, M.; Kharoubi-Hess, S.; Nilsson, S.; Ravel, D.; Ribeiro, M.; Grüring, C.; Ma, S.Y.; Padmanabhan, P.; et al. Infected Erythrocyte-Derived Extracellular Vesicles Alter Vascular Function via Regulatory Ago2-MiRNA Complexes in Malaria. Nat. Commun. 2016, 7, 12727. [Google Scholar] [CrossRef]
- Babatunde, K.A.; Mbagwu, S.; Hernández-Castañeda, M.A.; Adapa, S.R.; Walch, M.; Filgueira, L.; Falquet, L.; Jiang, R.H.Y.; Ghiran, I.; Mantel, P.-Y. Malaria Infected Red Blood Cells Release Small Regulatory RNAs through Extracellular Vesicles. Sci. Rep. 2018, 8, 884. [Google Scholar] [CrossRef]
- Burel, J.G.; Apte, S.H.; Groves, P.L.; Boyle, M.J.; Langer, C.; Beeson, J.G.; McCarthy, J.S.; Doolan, D.L. Dichotomous MiR Expression and Immune Responses Following Primary Blood-Stage Malaria. JCI Insight 2017, 2, e93434. [Google Scholar] [CrossRef]
- Gupta, H.; Chaudhari, S.; Rai, A.; Bhat, S.; Sahu, P.K.; Hande, M.H.; D’Souza, S.C.; Shashikiran, U.; Satyamoorthy, K. Genetic and Epigenetic Changes in Host ABCB1 Influences Malaria Susceptibility to Plasmodium Falciparum. PLoS ONE 2017, 12, e0175702. [Google Scholar] [CrossRef]
- Wang, Z.; Xi, J.; Hao, X.; Deng, W.; Liu, J.; Wei, C.; Gao, Y.; Zhang, L.; Wang, H. Red Blood Cells Release Microparticles Containing Human Argonaute 2 and MiRNAs to Target Genes of Plasmodium Falciparum. Emerg. Microbes Infect. 2017, 6, 1–11. [Google Scholar] [CrossRef]
- Li, J.; Huang, M.; Li, Z.; Li, W.; Wang, F.; Wang, L.; Li, X.; Zheng, X.; Zou, Y. Identification of Potential Whole Blood MicroRNA Biomarkers for the Blood Stage of Adult Imported Falciparum Malaria through Integrated MRNA and MiRNA Expression Profiling. Biochem. Biophys. Res. Commun. 2018, 506, 471–477. [Google Scholar] [CrossRef]
- Schrum, J.E.; Juliet, N.C.; Dobbs, K.R.; Michael, C.K.; George, W.R.; Gazzinelli, R.T.; Netea, M.G.; Kazura, J.W.; Dent, A.E.; Katherine, A.F. Plasmodium Falciparum Induces Trained Innate Immunity. J. Immunol. 2018, 200, 1243–1248. [Google Scholar] [CrossRef]
- Dandewad, V.; Vindu, A.; Joseph, J.; Seshadri, V. Import of Human MiRNA-RISC Complex into Plasmodium Falciparum and Regulation of the Parasite Gene Expression. J. Biosci. 2019, 44, 50. [Google Scholar] [CrossRef]
- Walk, J.; Keramati, F.; de Bree, L.C.J.; Arts, R.J.W.; Blok, B.; Netea, M.G.; Stunnenberg, H.G.; Sauerwein, R.W. Controlled Human Malaria Infection Induces Long-Term Functional Changes in Monocytes. Front. Mol. Biosci. 2020, 7, 604553. [Google Scholar] [CrossRef]
- Chakrabarti, M.; Garg, S.; Rajagopal, A.; Pati, S.; Singh, S. Targeted Repression of Plasmodium Apicortin by Host MicroRNA Impairs Malaria Parasite Growth and Invasion. Dis. Model. Mech. 2020, 13, dmm042820. [Google Scholar] [CrossRef]
- Ketprasit, N.; Cheng, I.S.; Deutsch, F.; Tran, N.; Imwong, M.; Combes, V.; Palasuwan, D. The Characterization of Extracellular Vesicles-Derived MicroRNAs in Thai Malaria Patients. Malar. J. 2020, 19, 285. [Google Scholar] [CrossRef]
- Bertrams, W.; Griss, K.; Han, M.; Seidel, K.; Hippenstiel, S.; Suttorp, N.; Finkernagel, F.; Wilhelm, J.; Vogelmeier, C.F.; Schmeck, B. Transcriptional Analysis Identifies Potential Biomarkers and Molecular Regulators in Acute Malaria Infection. Life Sci. 2021, 270, 119158. [Google Scholar] [CrossRef] [PubMed]
- Gupta, H.; Rubio, M.; Sitoe, A.; Varo, R.; Cisteró, P.; Madrid, L.; Cuamba, I.; Jimenez, A.; Martiáñez-Vendrell, X.; Barrios, D.; et al. Plasma MicroRNA Profiling of Plasmodium Falciparum Biomass and Association with Severity of Malaria Disease. Emerg. Infect. Dis. 2021, 27, 430–442. [Google Scholar] [CrossRef]
- Hadighi, R.; Heidari, A.; Fallah, P.; Keshavarz, H.; Tavakoli, Z.; Mansouri, S.; Sadrkhanloo, M. Key Plasma MicroRNAs Variations in Patients with Plasmodium Vivax Malaria in Iran. Heliyon 2022, 8, e09018. [Google Scholar] [CrossRef] [PubMed]
- Oxendine Harp, K.; Bashi, A.; Botchway, F.; Dei-adomakoh, Y.; Iqbal, S.A.; Wilson, M.D.; Adjei, A.A.; Stiles, J.K.; Driss, A.; Harp, K.O.; et al. MicroRNAs MiR-451a and Let-7i-5p Profiles in Circulating Exosomes Vary among Individuals with Different Sickle Hemoglobin Genotypes and Malaria. J. Clin. Med. 2022, 11, 500. [Google Scholar] [CrossRef] [PubMed]
- Chamnanchanunt, S.; Kuroki, C.; Desakorn, V.; Enomoto, M.; Thanachartwet, V.; Sahassananda, D.; Sattabongkot, J.; Jenwithisuk, R.; Fucharoen, S.; Svasti, S.; et al. Downregulation of Plasma MiR-451 and MiR-16 in Plasmodium Vivax Infection. Exp. Parasitol. 2015, 155, 19–25. [Google Scholar] [CrossRef]
- Kaur, H.; Sehgal, R.; Kumar, A.; Sehgal, A.; Bansal, D.; Sultan, A.A. Screening and Identification of Potential Novel Biomarker for Diagnosis of Complicated Plasmodium Vivax Malaria. J. Transl. Med. 2018, 16, 272. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.L.S.S.; Coimbra, R.S.; Sousa, T.N.; Guimarães, L.F.F.F.; Gomes, M.S.; Amaral, L.R.; Pereira, D.B.; Fontes, C.J.F.F.; Hawwari, I.; Franklin, B.S.; et al. The Interface Between Inflammatory Mediators and MicroRNAs in Plasmodium Vivax Severe Thrombocytopenia. Front. Cell. Infect. Microbiol. 2021, 11, 631333. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Zinger, A.; Tiberti, N.; Grau, G.E.R.; Combes, V. Differential Plasma Microvesicle and Brain Profiles of MicroRNA in Experimental Cerebral Malaria. Malar. J. 2018, 17, 192. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, S.-C.; Fan, X.-Y.; Jin, Y.-L.; Li, X.; Du, Y.-T. Plasmodium Manipulates the Expression of Host Long Non-Coding RNA during Red Blood Cell Intracellular Infection. Parasites Vectors 2022, 15, 182. [Google Scholar] [CrossRef]
- Cock-Rada, A.M.; Medjkane, S.; Janski, N.; Yousfi, N.; Perichon, M.; Chaussepied, M.; Chluba, J.; Langsley, G.; Weitzman, J.B. SMYD3 Promotes Cancer Invasion by Epigenetic Upregulation of the Metalloproteinase MMP-9. Cancer Res. 2012, 72, 810–820. [Google Scholar] [CrossRef]
- Marsolier, J.; Pineau, S.; Medjkane, S.; Perichon, M.; Yin, Q.Y.; Flemington, E.; Weitzman, M.D.; Weitzman, J.B. OncomiR Addiction Is Generated by a MiR-155 Feedback Loop in Theileria-Transformed Leukocytes. PLoS Pathog. 2013, 9, e1003222. [Google Scholar] [CrossRef]
- Gillan, V.; Simpson, D.M.; Kinnaird, J.; Maitland, K.; Shiels, B.; Devaney, E. Characterisation of Infection Associated MicroRNA and Protein Cargo in Extracellular Vesicles of Theileria Annulata Infected Leukocytes. Cell. Microbiol. 2019, 21, e12969. [Google Scholar] [CrossRef]
- Haidar, M.; Rchiad, Z.; Ansari, H.R.; Ben-Rached, F.; Tajeri, S.; Latre De Late, P.; Langsley, G.; Pain, A. MiR-126-5p by Direct Targeting of JNK-Interacting Protein-2 (JIP-2) Plays a Key Role in Theileria-Infected Macrophage Virulence. PLoS Pathog. 2018, 14, e1006942. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.; Denkers, E.Y. Toxoplasma gondii Inhibits Covalent Modification of Histone H3 at the IL-10 Promoter in Infected Macrophages. PLoS ONE 2009, 4, e7589. [Google Scholar] [CrossRef]
- Leng, J.; Butcher, B.A.; Egan, C.E.; Abi Abdallah, D.S.; Denkers, E.Y. Toxoplasma gondii Prevents Chromatin Remodeling Initiated by TLR-Triggered Macrophage Activation. J. Immunol. 2009, 182, 489–497. [Google Scholar] [CrossRef]
- Zeiner, G.M.; Norman, K.L.; Thomson, J.M.; Hammond, S.M.; Boothroyd, J.C. Toxoplasma gondii Infection Specifically Increases the Levels of Key Host MicroRNAs. PLoS ONE 2010, 5, e8742. [Google Scholar] [CrossRef]
- Lang, C.; Hildebrandt, A.; Brand, F.; Opitz, L.; Dihazi, H.; Luder, C.G.K. Impaired Chromatin Remodelling at STAT1-Regulated Promoters Leads to Global Unresponsiveness of Toxoplasma gondii-Infected Macrophages to IFN-Gamma. PLoS Pathog. 2012, 8, e1002483. [Google Scholar] [CrossRef]
- Cai, Y.; Chen, H.; Jin, L.; You, Y.; Shen, J. STAT3-Dependent Transactivation of MiRNA Genes Following Toxoplasma gondii Infection in Macrophage. Parasites Vectors 2013, 6, 356. [Google Scholar] [CrossRef]
- Xu, M.-J.; Zhou, D.-H.; Nisbet, A.J.; Huang, S.-Y.; Fan, Y.-F.; Zhu, X.-Q. Characterization of Mouse Brain MicroRNAs after Infection with Cyst-Forming Toxoplasma gondii. Parasites Vectors 2013, 6, 154. [Google Scholar] [CrossRef]
- Cai, Y.; Chen, H.; Mo, X.; Tang, Y.; Xu, X.; Zhang, A.; Lun, Z.; Lu, F.; Wang, Y.; Shen, J. Toxoplasma gondii Inhibits Apoptosis via a Novel STAT3-MiR-17–92-Bim Pathway in Macrophages. Cell. Signal. 2014, 26, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Cannella, D.; Brenier-Pinchart, M.-P.; Braun, L.; van Rooyen, J.M.; Bougdour, A.; Bastien, O.; Behnke, M.S.; Curt, R.-L.; Curt, A.; Saeij, J.P.J.; et al. MiR-146a and MiR-155 Delineate a MicroRNA Fingerprint Associated with Toxoplasma Persistence in the Host Brain. Cell Rep. 2014, 6, 928–937. [Google Scholar] [CrossRef]
- Jia, B.; Chang, Z.; Wei, X.; Lu, H.; Yin, J.; Jiang, N.; Chen, Q. Plasma MicroRNAs Are Promising Novel Biomarkers for the Early Detection of Toxoplasma gondii Infection. Parasites Vectors 2014, 7, 433. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kannan, G.; Pletnikov, M.V.; Yolken, R.H.; Xiao, J. Chronic Infection of Toxoplasma gondii Downregulates MiR-132 Expression in Multiple Brain Regions in a Sex-Dependent Manner. Parasitology 2014, 142, 623–632. [Google Scholar] [CrossRef]
- Xiao, J.; Li, Y.; Prandovszky, E.; Karuppagounder, S.S.; Talbot, C.C.; Dawson, V.L.; Dawson, T.M.; Yolken, R.H. MicroRNA-132 Dysregulation in Toxoplasma gondii Infection Has Implications for Dopamine Signaling Pathway. Neuroscience 2014, 268, 128–138. [Google Scholar] [CrossRef]
- Dvorakova-Hortova, K.; Sidlova, A.; Ded, L.; Hladovcova, D.; Vieweg, M.; Weidner, W.; Steger, K.; Stopka, P.; Paradowska-Dogan, A. Toxoplasma gondii Decreases the Reproductive Fitness in Mice. PLoS ONE 2014, 9, e96770. [Google Scholar] [CrossRef]
- Hari Dass, S.A.; Vyas, A.; Dass, S.A.H.; Vyas, A.; Hari Dass, S.A.; Vyas, A.; Dass, S.A.H.; Vyas, A.; Hari Dass, S.A.; Vyas, A.; et al. Toxoplasma gondii Infection Reduces Predator Aversion in Rats through Epigenetic Modulation in the Host Medial Amygdala. Mol. Ecol. 2014, 23, 6114–6122. [Google Scholar] [CrossRef]
- Rosowski, E.E.; Nguyen, Q.P.; Camejo, A.; Spooner, E.; Saeij, J.P.J. Toxoplasma gondii Inhibits Gamma Interferon (IFN-Gamma)- and IFN-Beta-Induced Host Cell STAT1 Transcriptional Activity by Increasing the Association of STAT1 with DNA. Infect. Immun. 2014, 82, 706–719. [Google Scholar] [CrossRef]
- He, J.-J.; Ma, J.; Wang, J.-L.; Xu, M.-J.; Zhu, X.-Q. Analysis of MiRNA Expression Profiling in Mouse Spleen Affected by Acute Toxoplasma gondii Infection. Infect. Genet. Evol. 2016, 37, 137–142. [Google Scholar] [CrossRef]
- Kim, M.-K.J.; Jung, B.-K.; Cho, J.; Song, H.; Pyo, K.-H.; Lee, J.M.; Kim, M.-K.J.; Chai, J.-Y. Exosomes Secreted by Toxoplasma gondii-Infected L6 Cells: Their Effects on Host Cell Proliferation and Cell Cycle Changes. Korean J. Parasitol. 2016, 54, 147–154. [Google Scholar] [CrossRef]
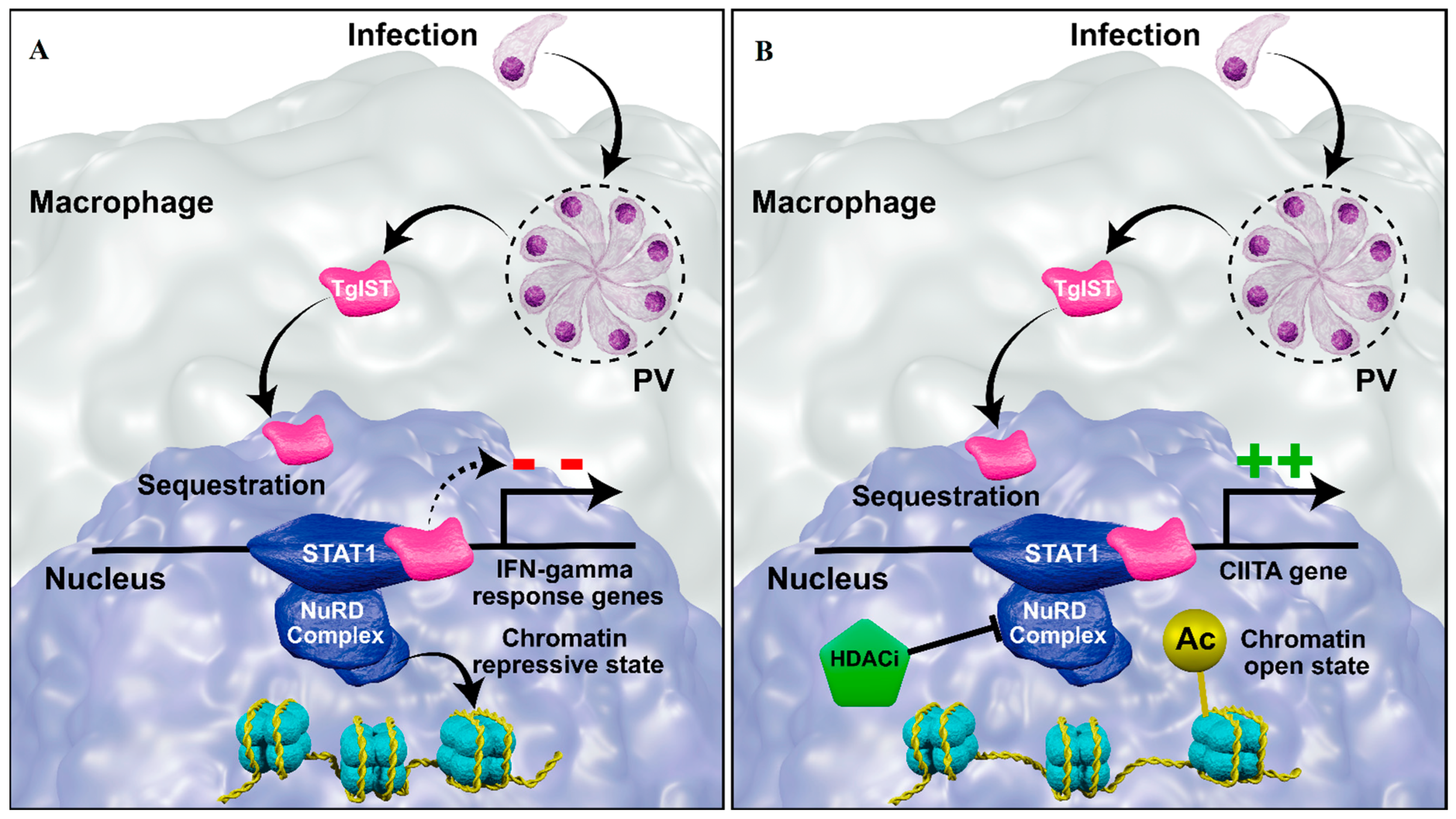
- Gay, G.; Braun, L.; Brenier-Pinchart, M.-P.P.; Vollaire, J.; Josserand, V.; Bertini, R.-L.L.; Varesano, A.; Touquet, B.; De Bock, P.-J.J.; Coute, Y.; et al. Toxoplasma gondii TgIST Co-Opts Host Chromatin Repressors Dampening STAT1-Dependent Gene Regulation and IFN-γ-Mediated Host Defenses. J. Exp. Med. 2016, 213, 1779–1798. [Google Scholar] [CrossRef]
- Olias, P.; Etheridge, R.D.; Zhang, Y.; Holtzman, M.J.; Sibley, L.D. Toxoplasma Effector Recruits the Mi-2/NuRD Complex to Repress STAT1 Transcription and Block IFN-γ-Dependent Gene Expression. Cell Host Microbe 2016, 20, 72–82. [Google Scholar] [CrossRef]
- Cong, W.; Zhang, X.-X.; He, J.-J.; Li, F.-C.; Elsheikha, H.M.; Zhu, X.-Q. Global MiRNA Expression Profiling of Domestic Cat Livers Following Acute Toxoplasma gondii Infection. Oncotarget 2017, 8, 25599–25611. [Google Scholar] [CrossRef]
- Ngo, H.M.; Zhou, Y.; Lorenzi, H.; Wang, K.; Kim, T.-K.; Zhou, Y.; El Bissati, K.; Mui, E.; Fraczek, L.; Rajagopala, S.V.; et al. Toxoplasma Modulates Signature Pathways of Human Epilepsy, Neurodegeneration & Cancer. Sci. Rep. 2017, 7, 11496. [Google Scholar] [CrossRef]
- Syn, G.; Anderson, D.; Blackwell, J.M.; Jamieson, S.E. Epigenetic Dysregulation of Host Gene Expression in Toxoplasma Infection with Specific Reference to Dopamine and Amyloid Pathways. Infect. Genet. Evol. 2018, 65, 159–162. [Google Scholar] [CrossRef]
- Hu, R.S.; He, J.J.; Elsheikha, H.M.; Zhang, F.K.; Zou, Y.; Zhao, G.H.; Cong, W.; Zhu, X.Q. Differential Brain MicroRNA Expression Profiles After Acute and Chronic Infection of Mice With Toxoplasma gondii Oocysts. Front. Microbiol. 2018, 9, 2316. [Google Scholar] [CrossRef]
- Liu, W.; Huang, L.; Wei, Q.; Zhang, Y.; Zhang, S.; Zhang, W.; Cai, L.; Liang, S. Microarray Analysis of Long Non-Coding RNA Expression Profiles Uncovers a Toxoplasma-Induced Negative Regulation of Host Immune Signaling. Parasites Vectors 2018, 11, 174. [Google Scholar] [CrossRef]
- Menard, K.L.; Haskins, B.E.; Colombo, A.P.; Denkers, E.Y. Toxoplasma gondii Manipulates Expression of Host Long Noncoding RNA during Intracellular Infection. Sci. Rep. 2018, 8, 15017. [Google Scholar] [CrossRef]
- Rezaei, F.; Daryani, A.; Sharifi, M.; Sarvi, S.; Jafari, N.; Pagheh, A.S.; Hashemi, N.; Hejazi, S.H. MiR-20a Inhibition Using Locked Nucleic Acid (LNA) Technology and Its Effects on Apoptosis of Human Macrophages Infected by Toxoplasma gondii RH Strain. Microb. Pathog. 2018, 121, 269–276. [Google Scholar] [CrossRef]
- Braun, L.; Brenier-Pinchart, M.-P.; Hammoudi, P.-M.; Cannella, D.; Kieffer-Jaquinod, S.; Vollaire, J.; Josserand, V.; Touquet, B.; Couté, Y.; Tardieux, I.; et al. The Toxoplasma Effector TEEGR Promotes Parasite Persistence by Modulating NF-ΚB Signalling via EZH2. Nat. Microbiol. 2019, 4, 1208–1220. [Google Scholar] [CrossRef]
- Detavernier, A.; Azouz, A.; Shehade, H.; Splittgerber, M.; Van Maele, L.; Nguyen, M.; Thomas, S.; Achouri, Y.; Svec, D.; Calonne, E.; et al. Monocytes Undergo Multi-Step Differentiation in Mice during Oral Infection by Toxoplasma gondii. Commun. Biol. 2019, 2, 472. [Google Scholar] [CrossRef]
- Hou, Z.; Liu, D.; Su, S.; Wang, L.; Zhao, Z.; Ma, Y.; Li, Q.; Jia, C.; Xu, J.; Zhou, Y.; et al. Comparison of Splenocyte MicroRNA Expression Profiles of Pigs during Acute and Chronic Toxoplasmosis. BMC Genom. 2019, 20, 97. [Google Scholar] [CrossRef]
- Li, D.-L.; Zou, W.-H.; Deng, S.-Q.; Peng, H.-J. Analysis of the Differential Exosomal MiRNAs of DC2.4 Dendritic Cells Induced by Toxoplasma gondii Infection. Int. J. Mol. Sci. 2019, 20, 5506. [Google Scholar] [CrossRef]
- Li, S.; Yang, J.; Wang, L.; Du, F.; Zhao, J.; Fang, R. Expression Profile of MicroRNAs in Porcine Alveolar Macrophages after Toxoplasma gondii Infection. Parasites Vectors 2019, 12, 65. [Google Scholar] [CrossRef]
- Lie, S.; Rochet, E.; Segerdell, E.; Ma, Y.; Ashander, L.M.; Shadforth, A.M.A.; Blenkinsop, T.A.; Michael, M.Z.; Appukuttan, B.; Wilmot, B.; et al. Immunological Molecular Responses of Human Retinal Pigment Epithelial Cells to Infection with Toxoplasma gondii. Front. Immunol. 2019, 10, 708. [Google Scholar] [CrossRef]
- Rochet, E.; Appukuttan, B.; Ma, Y.; Ashander, L.M.; Smith, J.R. Expression of Long Non-Coding RNAs by Human Retinal Muller Glial Cells Infected with Clonal and Exotic Virulent Toxoplasma gondii. Non-Coding RNA 2019, 5, 48. [Google Scholar] [CrossRef]
- Jiang, H.; Zhai, T.; Yu, Y.; Li, X.; Gong, P.; Zhang, X.; Li, G.; Li, J. Delayed IL-12 Production by Macrophages during Toxoplasma gondii Infection Is Regulated by MiR-187. Parasitol. Res. 2020, 119, 1023–1033. [Google Scholar] [CrossRef]
- Medina, L.; Castillo, C.; Liempi, A.; Guerrero-Muñoz, J.; Rojas-Pirela, M.; Maya, J.D.; Prieto, H.; Kemmerling, U.; Guerrero-Munoz, J.; Rojas-Pirela, M.; et al. Trypanosoma Cruzi and Toxoplasma gondii Induce a Differential MicroRNA Profile in Human Placental Explants. Front. Immunol. 2020, 11, 595250. [Google Scholar] [CrossRef]
- Meira-Strejevitch, C.S.; de Siqueira Pereira, I.; Hippólito, D.D.C.; Maia, M.M.; Cruz, A.B.; Gava, R.; Brandão de Mattos, C.C.; Frederico, F.B.; Siqueira, R.C.; Mattos, L.C.; et al. Ocular Toxoplasmosis Associated with Up-Regulation of MiR-155-5p/MiR-29c-3p and down-Regulation of MiR-21-5p/MiR-125b-5p. Cytokine 2020, 127, 154990. [Google Scholar] [CrossRef]
- de Siqueira Pereira, I.; Maia, M.M.; da Cruz, A.B.; Telles, J.P.M.; Vidal, J.E.; Gava, R.; Meira-Strejevitch, C.S.; Pereira-Chioccola, V.L.; De Siqueira, I.; Marta, P.; et al. Plasma Extracellular MicroRNAs Are Related to AIDS/Cerebral Toxoplasmosis Co-Infection. Parasite Immunol. 2020, 42, e12696. [Google Scholar] [CrossRef]
- Tyebji, S.; Hannan, A.J.; Tonkin, C.J. Pathogenic Infection in Male Mice Changes Sperm Small RNA Profiles and Transgenerationally Alters Offspring Behavior. Cell Rep. 2020, 31, 107573. [Google Scholar] [CrossRef]
- Zhou, C.-X.; Ai, K.; Huang, C.-Q.; Guo, J.-J.; Cong, H.; He, S.-Y.; Zhu, X.-Q. MiRNA and CircRNA Expression Patterns in Mouse Brain during Toxoplasmosis Development. BMC Genom. 2020, 21, 46. [Google Scholar] [CrossRef]
- Nast, R.; Choepak, T.; Lüder, C.G.K.; Lueder, C.G.K.; Lüder, C.G.K. Epigenetic Control of IFN-γ Host Responses During Infection with Toxoplasma gondii. Front. Immunol. 2020, 11, 581241. [Google Scholar] [CrossRef]
- Sabou, M.; Doderer-Lang, C.C.; Leyer, C.; Konjic, A.; Kubina, S.; Lennon, S.; Rohr, O.; Viville, S.S.; Cianférani, S.; Candolfi, E.; et al. Toxoplasma gondii ROP16 Kinase Silences the Cyclin B1 Gene Promoter by Hijacking Host Cell UHRF1-Dependent Epigenetic Pathways. Cell. Mol. Life Sci. 2020, 77, 2141–2156. [Google Scholar] [CrossRef]
- Sciume, G.; Mikami, Y.; Jankovic, D.; Nagashima, H.; Villarino, A.V.; Morrison, T.; Yao, C.; Signorella, S.; Sun, H.-W.W.; Brooks, S.R.; et al. Rapid Enhancer Remodeling and Transcription Factor Repurposing Enable High Magnitude Gene Induction upon Acute Activation of NK Cells. Immunity 2020, 53, 745–758.e4. [Google Scholar] [CrossRef]
- Singh, D.K.; Hari Dass, S.A.; Abdulai-Saiku, S.; Vyas, A. Testosterone Acts Within the Medial Amygdala of Rats to Reduce Innate Fear to Predator Odor Akin to the Effects of Toxoplasma gondii Infection. Front. Psychiatry 2020, 11, 630. [Google Scholar] [CrossRef]
- El-Sayad, M.; Abdel Rahman, M.; Hussein, N.; Abdel Aziz, R.; El-Taweel, H.A.; Abd El-Latif, N. MicroRNA-155 Expression and Butyrylcholinesterase Activity in the Liver Tissue of Mice Infected with Toxoplasma gondii (Avirulent and Virulent Strains). Acta Parasitol. 2021, 66, 1167–1176. [Google Scholar] [CrossRef]
- Maia, M.M.; Cruz, A.B.; de Siqueira Pereira, I.; Taniwaki, N.N.; Namiyama, G.M.; Pereira-Chioccola, V.L. Characterization of Murine Extracellular Vesicles and Toxoplasma gondii Infection. Parasite Immunol. 2021, 43, e12869. [Google Scholar] [CrossRef]
- Menard, K.L.; Bu, L.; Denkers, E.Y. Transcriptomics Analysis of Toxoplasma gondii-Infected Mouse Macrophages Reveals Coding and Noncoding Signatures in the Presence and Absence of MyD88. BMC Genom. 2021, 22, 130. [Google Scholar] [CrossRef]
- Quiarim, T.M.; Maia, M.M.; da Cruz, A.B.; Taniwaki, N.N.; Namiyama, G.M.; Pereira-Chioccola, V.L. Characterization of Extracellular Vesicles Isolated from Types I, II and III Strains of Toxoplasma gondii. Acta Trop. 2021, 219, 105915. [Google Scholar] [CrossRef]
- Yang, J.; He, Z.; Chen, C.; Li, S.; Qian, J.; Zhao, J.; Fang, R. Toxoplasma gondii Infection Inhibits Histone Crotonylation to Regulate Immune Response of Porcine Alveolar Macrophages. Front. Immunol. 2021, 12, 696061. [Google Scholar] [CrossRef]
- Hou, Z.; Wang, L.; Su, D.; Cai, W.; Zhu, Y.; Liu, D.; Huang, S.; Xu, J.; Pan, Z.; Tao, J. Global MicroRNAs Expression Profile Analysis Reveals Possible Regulatory Mechanisms of Brain Injury Induced by Toxoplasma gondii Infection. Front. Neurosci. 2022, 16, 827570. [Google Scholar] [CrossRef]
- Hou, Z.; Zhang, H.; Xu, K.; Zhu, S.; Wang, L.; Su, D.; Liu, J.; Su, S.; Liu, D.; Huang, S.; et al. Cluster Analysis of Splenocyte MicroRNAs in the Pig Reveals Key Signal Regulators of Immunomodulation in the Host during Acute and Chronic Toxoplasma gondii Infection. Parasites Vectors 2022, 15, 58. [Google Scholar] [CrossRef]
- Jiang, D.; Wu, S.; Xu, L.; Xie, G.; Li, D.; Peng, H. Anti-Infection Roles of MiR-155-5p Packaged in Exosomes Secreted by Dendritic Cells Infected with Toxoplasma gondii. Parasites Vectors 2022, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.-K.; Song, H.; Shin, H.; Chai, J.-Y. Exosomal MiRNA-21 from Toxoplasma gondii-Infected Microglial Cells Induces the Growth of U87 Glioma Cells by Inhibiting Tumor Suppressor Genes. Sci. Rep. 2022, 12, 16450. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-S.; Zhou, C.-X.; Elsheikha, H.M.; He, J.-J.; Zou, F.-C.; Zheng, W.-B.; Zhu, X.-Q.; Zhao, G.-H. Temporal Transcriptomic Changes in Long Non-Coding RNAs and Messenger RNAs Involved in the Host Immune and Metabolic Response during Toxoplasma gondii Lytic Cycle. Parasites Vectors 2022, 15, 22. [Google Scholar] [CrossRef]
- Guo, X.-D.; Zhou, C.-X.; Li, L.-Y.; Ai, K.; Wang, Y.-L.; Zhou, D.-H. Comprehensive Analysis of MRNA-LncRNA Co-Expression Profiles in Mouse Brain during Infection with Toxoplasma gondii. Acta Trop. 2023, 237, 106722. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Delic, D.; Sies, H.; Wunderlich, F.; Abdel-Baki, A.A.S.; Dkhil, M.A.M. Differential MiRNA Expression in the Mouse Jejunum during Garlic Treatment of Eimeria Papillata Infections. Parasitol. Res. 2011, 109, 387–394. [Google Scholar] [CrossRef]
- Dkhil, M.; Abdel-Baki, A.A.; Delić, D.; Wunderlich, F.; Sies, H.; Al-Quraishy, S. Eimeria papillata: Upregulation of Specific miRNA-Species in the Mouse Jejunum. Exp. Parasitol. 2011, 127, 581–586. [Google Scholar] [CrossRef]
- Bigna, J.J.; Tochie, J.N.; Tounouga, D.N.; Bekolo, A.O.; Ymele, N.S.; Youda, E.L.; Sime, P.S.; Nansseu, J.R. Global, Regional, and Country Seroprevalence of Toxoplasma gondii in Pregnant Women: A Systematic Review, Modelling and Meta-Analysis. Sci. Rep. 2020, 10, 12102. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2019; WHO: Geneva, Switzerland, 2019; ISBN 9789241565721. [Google Scholar]
- King, P.; Tyler, K.M.; Hunter, P.R. Anthroponotic Transmission of Cryptosporidium parvum Predominates in Countries with Poorer Sanitation: A Systematic Review and Meta-Analysis. Parasites Vectors 2019, 12, 16. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Longcore, T.; Barbieri, M.; Dabritz, H.; Hill, D.; Klein, P.N.; Lepczyk, C.; Lilly, E.L.; McLeod, R.; Milcarsky, J.; et al. The One Health Approach to Toxoplasmosis: Epidemiology, Control, and Prevention Strategies. Ecohealth 2019, 16, 378–390. [Google Scholar] [CrossRef]
- Lodoen, M.B.; Smith, N.C.; Soldati-Favre, D.; Ferguson, D.J.P.; van Dooren, G.G. Nanos Gigantium Humeris Insidentes: Old Papers Informing New Research into Toxoplasma gondii. Int. J. Parasitol. 2021, 51, 1193–1212. [Google Scholar] [CrossRef]
- Christensen, P.; Racklyeft, A.; Ward, K.E.; Matheson, J.; Suwanarusk, R.; Chua, A.C.Y.; Kaneko, O.; Aung, H.L.; Rénia, L.; Amanzougaghene, N.; et al. Improving in Vitro Continuous Cultivation of Plasmodium cynomolgi, a Model for P. Vivax. Parasitol. Int. 2022, 89, 102589. [Google Scholar] [CrossRef]
- Certad, G.; Viscogliosi, E.; Chabé, M.; Cacciò, S.M. Pathogenic Mechanisms of Cryptosporidium and Giardia. Trends Parasitol. 2017, 33, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, M.S.; Morris, K.V. Transcriptional Gene Silencing in Humans. Nucleic Acids Res. 2016, 44, 6505–6517. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Liu, C.; Li, H.; Zhang, L.; Luo, G.; Liang, S.; Lü, M. Research Progress on the Interactions between Long Non-Coding RNAs and MicroRNAs in Human Cancer (Review). Oncol. Lett. 2020, 19, 595–605. [Google Scholar] [CrossRef]
- Singh, B.; Varikuti, S.; Halsey, G.; Volpedo, G.; Hamza, O.M.; Satoskar, A.R. Host-Directed Therapies for Parasitic Diseases. Future Med. Chem. 2019, 11, 1999–2018. [Google Scholar] [CrossRef]
- Coetzee, N.; von Grüning, H.; Opperman, D.; van der Watt, M.; Reader, J.; Birkholtz, L.-M. Epigenetic Inhibitors Target Multiple Stages of Plasmodium Falciparum Parasites. Sci. Rep. 2020, 10, 2355. [Google Scholar] [CrossRef]
- Pennini, M.E.; Pai, R.K.; Schultz, D.C.; Boom, W.H.; Harding, C.V. Mycobacterium tuberculosis 19-KDa Lipoprotein Inhibits IFN-γ-Induced Chromatin Remodeling of MHC2TA by TLR2 and MAPK Signaling. J. Immunol. 2006, 176, 4323–4330. [Google Scholar] [CrossRef]
- Parmar, N.; Chandrakar, P.; Kar, S. Leishmania Donovani Subverts Host Immune Response by Epigenetic Reprogramming of Macrophage M(Lipopolysaccharides + IFN-γ)/M(IL-10) Polarization. J. Immunol. 2020, 204, 2762–2778. [Google Scholar] [CrossRef]
- Lima, T.S.; Lodoen, M.B. Mechanisms of Human Innate Immune Evasion by Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2019, 9, 103. [Google Scholar] [CrossRef]
- Ramirez Ramirez, A.D.; de Jesus, M.C.S.; Rossit, J.; Reis, N.F.; Santos-Filho, M.C.; Sudré, A.P.; de Oliveira-Ferreira, J.; de Souza Baptista, A.R.; Storti-Melo, L.M.; Machado, R.L.D. Association of Toll-like Receptors in Malaria Susceptibility and Immunopathogenesis: A Meta-Analysis. Heliyon 2022, 8, e09318. [Google Scholar] [CrossRef]
- Jan, I.; Rather, R.A.; Mushtaq, I.; Malik, A.A.; Besina, S.; Baba, A.B.; Farooq, M.; Yousuf, T.; Rah, B.; Afroze, D. Helicobacter Pylori Subdues Cytokine Signaling to Alter Mucosal Inflammation via Hypermethylation of Suppressor of Cytokine Signaling 1 Gene During Gastric Carcinogenesis. Front. Oncol. 2021, 10, 604747. [Google Scholar] [CrossRef] [PubMed]
- Marr, A.K.; MacIsaac, J.L.; Jiang, R.; Airo, A.M.; Kobor, M.S.; McMaster, W.R. Leishmania donovani Infection Causes Distinct Epigenetic DNA Methylation Changes in Host Macrophages. PLoS Pathog. 2014, 10, e1004419. [Google Scholar] [CrossRef]
- Ullah, H.; Di Minno, A.; Esposito, C.; El-Seedi, H.R.; Khalifa, S.A.M.; Baldi, A.; Greco, A.; Santonastaso, S.; Cioffi, V.; Sperandeo, R.; et al. Efficacy of a Food Supplement Based on S-Adenosyl Methionine and Probiotic Strains in Subjects with Subthreshold Depression and Mild-to-Moderate Depression: A Monocentric, Randomized, Cross-over, Double-Blind, Placebo-Controlled Clinical Trial. Biomed. Pharmacother. 2022, 156, 113930. [Google Scholar] [CrossRef]
- Navik, U.; Sheth, V.G.; Kabeer, S.W.; Tikoo, K. Dietary Supplementation of Methyl Donor L-Methionine Alters Epigenetic Modification in Type 2 Diabetes. Mol. Nutr. Food Res. 2019, 63, 1801401. [Google Scholar] [CrossRef]
- McConkey, G.A.; Martin, H.L.; Bristow, G.C.; Webster, J.P. Toxoplasma gondii Infection and Behaviour—Location, Location, Location? J. Exp. Biol. 2013, 216, 113–119. [Google Scholar] [CrossRef]
- Ratti, M.; Lampis, A.; Ghidini, M.; Salati, M.; Mirchev, M.B.; Valeri, N.; Hahne, J.C. MicroRNAs (MiRNAs) and Long Non-Coding RNAs (LncRNAs) as New Tools for Cancer Therapy: First Steps from Bench to Bedside. Target. Oncol. 2020, 15, 261–278. [Google Scholar] [CrossRef]
- Jung, T.W.; Park, J.; Sun, J.L.; Ahn, S.H.; Abd El-Aty, A.M.; Hacimuftuoglu, A.; Kim, H.-C.; Shim, J.-H.; Shin, S.; Jeong, J.H. Administration of Kynurenic Acid Reduces Hyperlipidemia-Induced Inflammation and Insulin Resistance in Skeletal Muscle and Adipocytes. Mol. Cell. Endocrinol. 2020, 518, 110928. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite | Regulatory Mechanism | Target Process | Research Model | Country | Ref. | |
|---|---|---|---|---|---|---|
| In Vitro/In Silico | In Vivo | |||||
| C. parvum | miRNA | Host defense | H69 cells | - | USA | [11,12,13,14] |
| C. parvum | miRNA | Host defense | H69 and HIBEpiC cells | - | USA | [15,16] |
| C. parvum | miRNA | Cell proliferation | H69 and HIBEpiC cell lines | - | USA, China | [17] |
| C. parvum | miRNA | Host defense | H69, HIBEpiC, 603B, IEC4.1, SW480 cells | C57BL/6J mice | USA | [18] |
| C. parvum | miRNA | Host defense | H69, 603B, IEC4.1 | C57BL/6J mice | USA | [19] |
| C. parvum | miRNA | Host defense | H69 cells | - | USA, China | [20] |
| C. parvum | miRNA | Host defense | mICcl2 cells | C57BL/6 mice | France | [21] |
| C. parvum | PTHM | Host defense | FHs 74 INT, HCT-8, IEC4.1 cells | Mice in vivo infection | USA | [22] |
| C. parvum | PTHM | Cell migration | FHs 74 INT, HCT-8 cells | - | USA, China | [23,24] |
| C. parvum | miRNA | Apoptosis; cancer | HT-29 and HCT-8 cells | - | Malaysia | [25] |
| C. parvum | lncRNA; PTHM | Host defense | IEC4.1, muINTEPI cells | C57BL/6J mice | USA | [26] |
| C. parvum | lncRNA | Cell adhesion | HCT-8 cells | - | China | [27] |
| C. parvum | PTHM | Host defense | FHs 74 INT, HCT-8 cells | - | USA, China | [28] |
| C. parvum | miRNA | Host defense | HCT-8 cells | - | China | [29] |
| C. parvum | miRNA | Host defense; apoptosis | HCT-8 cells | - | China | [30] |
| C. parvum | lncRNA | Host defense | IEC4.1, RAW264.7 cells | C57BL/6J mice | USA | [31] |
| C. parvum | miRNA | Cell cycle regulation; cancer | Bioinformatics (GEO)/in silico | - | UK, Saudi Arabia, India | [32] |
| C. parvum | circRNA | Apoptosis | HCT-8 cells | - | China | [33] |
| C. parvum | lncRNA; PTHM | Host defense | HCT-8, IEC4.1 cells | C57BL/6J mice | USA | [34] |
| C. parvum | PTHM | Host defense; cancer | HCT-8 cells | CB17-SCID mice | France, Belgium | [35] |
| C. parvum | miRNA | Host defense; cell proliferation; apoptosis | HCT-8 cells | - | Turkey | [36] |
| C. parvum | miRNA | Apoptosis | HCT-8 cells | - | China | [37] |
| P. berghei | miRNA | Host defense | - | C57BL/6 and CBA mice | Australia, Denmark | [38] |
| P. berghei | miRNA | Host defense; cell proliferation; apoptosis | HEK293T cells | C57BL/6J and NMRI mice | Germany, USA | [39] |
| P. berghei and P. yoelli | miRNA | Host defense; cell adhesion; endocytosis; apoptosis | - | CBA mice | Spain, Australia, Peru | [40] |
| P. chabaudi | miRNA | Host defense; apoptosis | - | C57BL/6 mice | Germany, Saudi Arabia | [41] |
| P. chabaudi | miRNA | Host defense; cell cycle regulation; apoptosis; cancer | - | C57BL/6 mice | Saudi Arabia, Germany, Egypt | [42] |
| P. chabaudi | DNAm | Host defense | - | C57BL/6 mice | Saudi Arabia, Germany, Egypt | [43] |
| P. chabaudi | DNAm | Host defense; erythropoiesis | - | Balb/c mice | Saudi Arabia, Germany, Egypt, Spain | [44] |
| P. chabaudi | PTHM | Host defense | - | C57BL/6J mice | UK | [45] |
| P. falciparum | miRNA | Host defense | Human RBC, J2E, F4N, BW5147, J558L and 416B cells | - | UK | [46] |
| P. falciparum | miRNA | Host defense | Human RBC | - | USA, UK | [47] |
| P. falciparum | miRNA | Host defense | Human RBC | - | USA, Switzerland, UK | [48] |
| P. falciparum | miRNA | Host defense; cell communication | Human RBC | - | USA, Switzerland | [49] |
| P. falciparum | miRNA | Host defense | - | Artificial human infection | Australia | [50] |
| P. falciparum | DNAm | Drug resistance | - | Malaria patients | India | [51] |
| P. falciparum | miRNA | Host defense | Human RBC | - | China | [52] |
| P. falciparum and P. vivax | miRNA | Host defense; cell adhesion | - | Malaria patients | China | [53] |
| P. falciparum | PTHM | Host defense | - | Malaria patients | USA, Brazil, Netherlands | [54] |
| P. falciparum | miRNA | Host defense | Human RBC | - | India | [55] |
| P. falciparum | PTHM | Host defense | Artificial human infection | Netherlands, Denmark, Germany | [56] | |
| P. falciparum | miRNA | Host defense | Human RBC and HEK 293T cells | - | India, Belgium | [57] |
| P. falciparum and P. vivax | miRNA | Host defense; cell adhesion | - | Malaria patients | Thailand, Australia | [58] |
| P. falciparum | miRNA | Host defense | - | Malaria patients | Germany | [59] |
| P. falciparum | miRNA | Host defense; cell proliferation; neural pathways | HBE cells | Malaria patients | Spain, Mozambique, USA | [60] |
| P. falciparum | miRNA | Host defense; cell adhesion; erythropoiesis | - | Malaria patients | Iran | [61] |
| P. falciparum | miRNA | Host defense | - | Malaria patients | USA, Ghana | [62] |
| P. vivax | miRNA | Host defense | - | Malaria patients | Thailand, Japan | [63] |
| P. vivax | miRNA | Host defense; erythropoiesis; apoptosis | - | Malaria patients | India, Qatar | [64] |
| P. vivax | miRNA | Host defense | - | Malaria patients | Brazil, Germany | [65] |
| P. yoelli | miRNA | Host defense; neural pathways | - | CBA mice | Australia, USA | [66] |
| P. yoelli | lncRNA | Host defense | - | BALB/c mice | China | [67] |
| T. annulata | PTHM | Cancer | BL3 cells | - | France | [68] |
| T. annulata | miRNA | Cancer | BL3 cells | - | France, USA | [69] |
| T. annulata | miRNA | Host defense; cancer | BL20 cells | - | UK | [70] |
| T. annulata | miRNA | Cancer | BL3,BL20 cells | - | Saudi Arabia, France, Japan | [71] |
| T. gondii | PTHM | Host defense | - | C57BL/6 mice | USA | [72] |
| T. gondii | PTHM | Host defense | - | C57BL/6 mice | USA | [73] |
| T. gondii | miRNA | Apoptosis | HFF, VERO cells | - | USA | [74] |
| T. gondii | PTHM | Host defense | RAW264.7, BMDM (BALB/c mice) cells | - | Germany | [75] |
| T. gondii | miRNA | Apoptosis | Human macrophage, THP-1 cells | - | China, Australia | [76] |
| T. gondii | miRNA | Host defense | - | Kunming mice | China, UK | [77] |
| T. gondii | miRNA | Apoptosis | Human macrophage, THP-1 cells | - | China | [78] |
| T. gondii | miRNA | Host defense | HFF cells | Swiss OF1, BALB/cJRj, C57BL/6JRj mice | France, USA | [79] |
| T. gondii | miRNA | Host defense; cell cycle regulation | - | BALB/c mice | China | [80] |
| T. gondii | miRNA | Neural pathways | - | BALB/c mice | USA, China | [81] |
| T. gondii | miRNA | Neural pathways | SK-N-MC cells | CD-1 mice | USA, China | [82] |
| T. gondii | DNAm | Reproductive pathways | - | C57Bl/6 mice | Czech Republic, Germany | [83] |
| T. gondii | DNAm | Neural pathways | - | Wistar rats | Singapore | [84] |
| T. gondii | PTHM | Host defense | HFF, 293FT, HEK293 cells | - | USA | [85] |
| T. gondii | miRNA | Host defense; apoptosis; cancer | - | Kunming mice | China | [86] |
| T. gondii | miRNA | Cell cycle regulation; cell proliferation | L6 cells | - | Korea | [87] |
| T. gondii | PTHM | Host defense | HFF, RAW264.7, 2fTGH, U3A, HEK-Blue IFN-γ cells | BALB/cJRj mice | France | [88] |
| T. gondii | PTHM | Host defense | RAW 264.7, U3A, U3A-STAT1, HeLa cells | CD-1, C57BL/6 mice | USA | [89] |
| T. gondii | miRNA | Host defense; apoptosis | - | Chinese Li Hua domestic cats (Felis catus) | China | [90] |
| T. gondii | miRNA | Neural pathways | L-NSC, S-NSC, human monocytic cells | Toxoplasmosis patients | USA, UK, Canada, Australia | [91] |
| T. gondii | DNAm | Neural pathways | WERI-Rb-1 cells | - | Australia | [92] |
| T. gondii | miRNA | Host defense; cancer | - | BALB/c mice | China, UK | [93] |
| T. gondii | lncRNA | Host defense | HFF, THP-1 cells | - | China | [94] |
| T. gondii | lncRNA | Host defense | BMDM (C57BL/6 mice) | - | USA | [95] |
| T. gondii | miRNA | Apoptosis | Human macrophage | - | Iran | [96] |
| T. gondii | PTHM | Host defense | BMDM (C57BL/6 mice), HFF, 293-T-Rex cells | BALB/cJRj mice | France | [97] |
| T. gondii | PTHM | Host defense | HFF cells | C57BL/6 mice | Belgium, Czech Republic | [98] |
| T. gondii | miRNA | Host defense; apoptosis | - | Pigs (Sus scrofa) | China | [99] |
| T. gondii | miRNA | Host defense; apoptosis | DC2.4, HFF cells | - | China | [100] |
| T. gondii | miRNA | Host defense; cell proliferation; apoptosis | HFF, porcine alveolar macrophages cells | - | China | [101] |
| T. gondii | miRNA | Host defense | Primary human retinal pigment epithelial cells, ARPE-19 cells | - | Australia, USA | [102] |
| T. gondii | lncRNA | Host defense; cell proliferation; cancer | Primary retinal Müller cells and MIO-M1 cells | - | Australia | [103] |
| T. gondii | miRNA | Host defense | Mouse macrophage (C57BL/6J mice) | - | China | [104] |
| T. gondii | miRNA | Host defense; apoptosis; cell cycle regulation | Human placental explants | - | Chile | [105] |
| T. gondii | miRNA | Host defense | - | Ocular toxoplasmosis patients | Brazil | [106] |
| T. gondii | miRNA | Host defense | - | Toxoplasmosis and AIDS patients | Brazil | [107] |
| T. gondii | miRNA | Neural pathways | - | C57BL/6J mice | Australia | [108] |
| T. gondii | circRNA; miRNA | Host defense; cell proliferation; erythropoiesis; cancer | - | BALB/c mice | China | [109] |
| T. gondii | PTHM; DNAm | Host defense | RAW264.7 cells | - | Germany | [110] |
| T. gondii | PTHM; DNAm | Cell cycle regulation | BeWo, U-118MG cells | - | France | [111] |
| T. gondii | PTHM | Host defense | - | C57BL/6J mice | USA, Italy | [112] |
| T. gondii | DNAm | Neural pathways | - | Wistar rats | Singapore | [113] |
| T. gondii | miRNA | Host defense | - | Swiss Albin mice | Egypt | [114] |
| T. gondii | miRNA | Host defense | - | A/Sn mice | Brazil | [115] |
| T. gondii | lncRNA | Apoptosis | BMDM (C57BL/6 mice) | - | USA | [116] |
| T. gondii | miRNA | Host defense | VERO cells | - | Brazil | [117] |
| T. gondii | PTHM | Host defense; cell proliferation | 3D4/21, PK-15 cells | - | China | [118] |
| T. gondii | miRNA | Host defense; cell proliferation; apoptosis; cancer | - | Pigs (Sus scrofa) | China | [119] |
| T. gondii | miRNA | Host defense | - | Pigs (Sus scrofa) | China | [120] |
| T. gondii | miRNA | Host defense | DC2.4, HFF, RAW264.7 cells | - | China | [121] |
| T. gondii | miRNA | Cell proliferation | BV2, U87, U118 cells | C57BL/6 mice | Korea | [122] |
| T. gondii | lncRNA | Host defense | HFF cells | - | China, UK | [123] |
| T. gondii | lncRNA | Host defense | - | BALB/c, Kunming mice | China | [124] |
| E. papillata | miRNA | Cancer; host defense; apoptosis | - | Balb/c mice | Saudi Arabia, Germany, Egypt | [125,126] |
| Parasites | Regulatory Mechanism | Targeted Process | Research Model | ||||
|---|---|---|---|---|---|---|---|
| Specie or Genus | Occurrence | Mechanism | Occurrence | Pathway | Occurrence | Model | Occurrence |
| T. gondii | 53/116; 45.7% | ncRNA | 88/116; 75.9% | Host defense | 87/116; 75% | in vitro | 72/116; 62.1% |
| Plasmodium spp. | 30/116; 25.8% | miRNA | 76/88; 86.4% | Apoptosis | 23/116; 19.8% | in vivo | 63/116; 54.3% |
| C. parvum | 27/116;23.3% | lncRNA | 11/88; 12.5% | Cancer | 14/116; 12% | ||
| T. annulata | 4/116; 3.5% | circRNA | 2/88; 2.3% | Cell proliferation | 11/116; 9.5% | ||
| E. papillata | 2/116; 1.8% | PTHM | 23/116; 19.8% | Neural pathways | 9/116;7.8% | ||
| DNAm | 9/116; 7.8% | Cell cycle regulation | 6/116; 5.2% | ||||
| Cell adhesion | 5/116; 4.3% | ||||||
| Erythropoiesis | 4/116; 3.4% | ||||||
| Cell migration | 2/116; 1.7% | ||||||
| Cell comunication | 1/116; 0.9% | ||||||
| Reproductive pathway | 1/116; 0.9% | ||||||
| Parasite | Pathway | Compound | Compound Analogous in Clinical Trial | Ref. | ||||
|---|---|---|---|---|---|---|---|---|
| Compound | Phase | Disease | Number | Status | ||||
| T. gondii | miRNA | anti-miR21 | RG012 | 1 | AS | NCT03373786 | Completed | [122] |
| T. gondii | PTHM | MS-275 | Entinostat (MS-275) | 3 | BC | NCT02115282 | Active | [75] |
| T. gondii | DNAm | L-methionine | SAMe | 4 | RMD | NCT04832178 | Active | [84] |
| T. gondii | PTHM | HDAC2- SiRNA | Vorinostat; romidepsin | FDA approved | CTCL | - | - | [118] |
| T. annulata | miRNA | anti-miR- 155 | Cobomarsen (MRG-106) | 2 | CTCL | NCT02580552 | Complete | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandão, Y.d.O.; Molento, M.B. A Systematic Review of Apicomplexa Looking into Epigenetic Pathways and the Opportunity for Novel Therapies. Pathogens 2023, 12, 299. https://doi.org/10.3390/pathogens12020299
Brandão YdO, Molento MB. A Systematic Review of Apicomplexa Looking into Epigenetic Pathways and the Opportunity for Novel Therapies. Pathogens. 2023; 12(2):299. https://doi.org/10.3390/pathogens12020299
Chicago/Turabian StyleBrandão, Yara de Oliveira, and Marcelo Beltrão Molento. 2023. "A Systematic Review of Apicomplexa Looking into Epigenetic Pathways and the Opportunity for Novel Therapies" Pathogens 12, no. 2: 299. https://doi.org/10.3390/pathogens12020299