
Advances in Babesia Vaccine Development: An Overview

Abstract
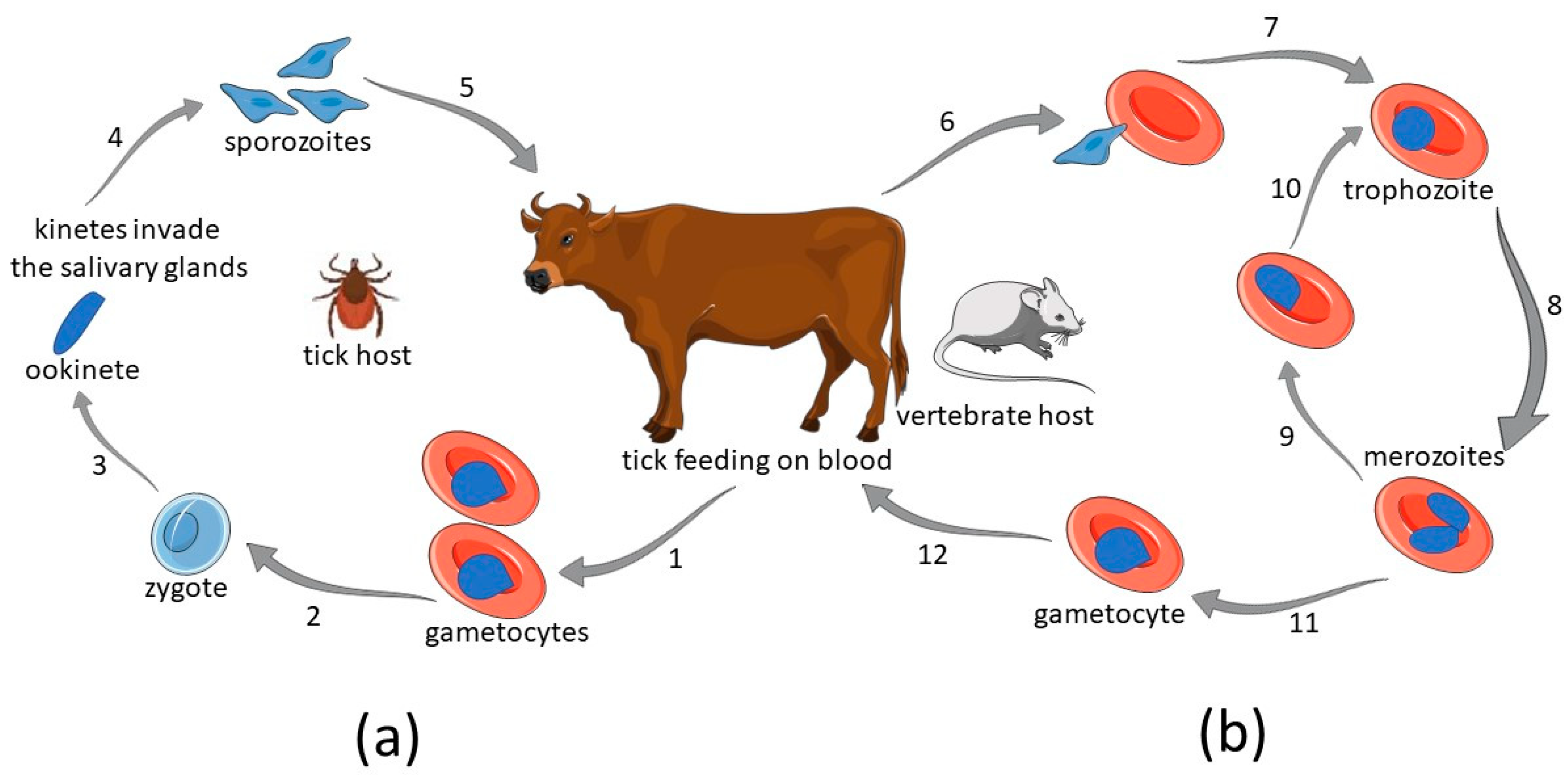
:1. Introduction
2. Advances in B. microti Vaccine Development
2.1. Characterization of Whole-Parasite Vaccines
2.1.1. Inactivated (Killed) Whole-Parasite Vaccines
2.1.2. Attenuated Whole-Parasite Vaccines
2.1.3. Limitations of Whole-Parasite Vaccines
2.1.4. Further Development of Whole-Parasite Vaccines
2.2. Characterization of Subunit Vaccines
2.2.1. Limitations of Protein-Based Subunit Vaccines
2.2.2. Further Development of Protein-Based Subunit Vaccines
3. Bovine Babesiosis Vaccines
3.1. Vaccination Achievements in B. bovis Combat
Ideas for the Future of Vaccine Development against B. bovis
3.2. Vaccine Models for B. bigemina
3.2.1. Protein-Based Vaccines
3.2.2. Live Attenuated Vaccines
3.2.3. Transfection of B. bigemina
4. Remaining Babesia Species Vaccine Development
5. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bock, R.E.; de Vos, A.J.; Kingston, T.G.; Shiels, I.A.; Dalgliesh, R.J. Investigations of breakdowns in protection provided by living Babesia bovis vaccine. Vet. Parasitol. 1992, 43, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Homer, M.J.; Aguilar-Delfin, I.; Telford III, S.R.; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Westblade, L.F.; Simon, M.S.; Mathison, B.A.; Kirkman, L.A. Babesia microti: From Mice to Ticks to an Increasing Number of Highly Susceptible Humans. J. Clin. Microbiol. 2017, 55, 2903–2912. [Google Scholar] [CrossRef]
- Babes, V. Sur l’he’moglobinurie bacte´rienne du boeuf. CR Acad. Sci. 1888, 107, 692–694. [Google Scholar]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Cornillot, E.; Hadj-Kaddour, K.; Dassouli, A.; Noel, B.; Ranwez, V.; Vacherie, B.; Augagneur, Y.; Bres, V.; Duclos, A.; Randazzo, S.; et al. Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res. 2012, 40, 9102–9114. [Google Scholar] [CrossRef]
- Villatoro, T.; Karp, J.K. Transfusion-Transmitted Babesiosis. Arch. Pathol. Lab. Med. 2018, 143, 130–134. [Google Scholar] [CrossRef]
- Fox, L.M.; Wingerter, S.; Ahmed, A.; Arnold, A.; Chou, J.; Rhein, L.; Levy, O. Neonatal babesiosis: Case report and review of the literature. Pediatr. Infect. Dis. J. 2006, 25, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; Linden, J.V.; Bosserman, E.; Young, C.; Olkowska, D.; Wilson, M. Transfusion-Associated Babesiosis in the United States: A Description of Cases. Ann. Intern. Med. 2011, 155, 509–519. [Google Scholar] [CrossRef]
- Brennan, M.B.; Herwaldt, B.L.; Kazmierczak, J.J.; Weiss, J.W.; Klein, C.L.; Leith, C.P.; He, R.; Oberley, M.J.; Tonnetti, L.; Wilkins, P.P.; et al. Transmission of Babesia microti Parasites by Solid Organ Transplantation. Emerg. Infect. Dis. 2016, 22, 1869–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, M.; Wei, Q.; Zamoto, A.; Morita, C.; Arai, S.; Shiota, T.; Fujimagari, M.; Itagaki, A.; Fujita, H.; Ishihara, C. Human Babesiosis in Japan: Epizootiologic Survey of Rodent Reservoir and Isolation of New Type of Babesia microti-Like Parasite. J. Clin. Microbiol. 2001, 39, 4316–4322. [Google Scholar] [CrossRef]
- Zamoto, A.; Tsuji, M.; Wei, Q.; Cho, S.H.; Shin, E.H.; Kim, T.S.; Leonova, G.N.; Hagiwara, K.; Asakawa, M.; Kariwa, H.; et al. Epizootiologic survey for Babesia microti among small wild mammals in northeastern Eurasia and a geographic diversity in the beta-tubulin gene sequences. J. Vet. Med. Sci. 2004, 66, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Hersh, M.H.; Tibbetts, M.; Strauss, M.; Ostfeld, R.S.; Keesing, F. Reservoir Competence of Wildlife Host Species for Babesia microti. Emerg. Infect. Dis. 2012, 18, 1951–1957. [Google Scholar] [CrossRef]
- Friedhoff, K.T.; Smith, R.D. Transmission of Babesia by ticks. In Babesiosis; Kreier, M.R., Kreier, J.P., Eds.; Academic Press: New York, NY, USA, 1981; pp. 267–321. [Google Scholar]
- Sanchez, E.; Vannier, E.; Wormser, G.P.; Hu, L.T. Diagnosis, treatment, and prevention of lyme disease, human Granulocytic anaplasmosis, and babesiosis: A review. JAMA 2016, 315, 1767–1777. [Google Scholar] [CrossRef]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The Complexity of Piroplasms Life Cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a Bovine Blood Parasite of Veterinary and Zoonotic Importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef]
- Falagas, M.E.; Klempner, M.S. Babesiosis in Patients with AIDS: A Chronic Infection Presenting as Fever of Unknown Origin. Clin. Infect. Dis. 1996, 22, 809–812. [Google Scholar] [CrossRef]
- Hatcher, J.C.; Greenberg, P.D.; Antique, J.; Jimenez-Lucho, V.E. Severe Babesiosis in Long Island: Review of 34 Cases and Their Complications. Clin. Infect. Dis. 1996, 32, 1117–1125. [Google Scholar] [CrossRef]
- Froberg, M.K.; Dannen, D.; Bakken, J.S. Babesiosis and HIV. Lancet 2004, 363, 704. [Google Scholar] [CrossRef]
- Krause, P.J.; Daily, J.; Telford, S.R.; Vannier, E.; Lantos, P.; Spielman, A. Shared features in the pathobiology of babesiosis and malaria. Trends Parasitol. 2007, 23, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Dumic, I.; Madrid, C.; Prada, L.R.; Nordstrom, C.W.; Taweesedt, P.T.; Ramanan, P. Splenic Complications of Babesia microti Infection in Humans: A Systematic Review. Can. J. Infect. Dis. Med. Microbiol. 2020, 2020, 6934149. [Google Scholar] [CrossRef]
- Smith, T.; Kilborne, F.L. Investigations into the nature, causation and prevention of Southern cattle fever. USDA Bur. Anim Lnd Bull. 1893, 1, 1–372. [Google Scholar]
- M’Fadyean, J.; Stockman, S. A New Species of Piroplasm Found in the Blood of British Cattle. J. Comp. Pathol. Ther. 1911, 24, 340–354. [Google Scholar] [CrossRef]
- Howell, J.M.; Ueti, M.W.; Palmer, G.H.; Scoles, G.A.; Knowles, D.P. Persistently Infected Calves as Reservoirs for Acquisition and Transovarial Transmission of Babesia bovis by Rhipicephalus (Boophilus) microplus. J. Clin. Microbiol. 2007, 45, 3155–3159. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, D.F.; Ross, D.R. Epizootiological factors in the control of bovine babesiosis. Aust. Vet. J. 1972, 48, 292–298. [Google Scholar] [CrossRef]
- Hunfeld, K.-P.; Hildebrandt, A.; Gray, J.S. Babesiosis: Recent insights into an ancient disease. Int. J. Parasitol. 2008, 38, 1219–1237. [Google Scholar] [CrossRef]
- Suarez, C.E.; Noh, S. Emerging perspectives in the research of bovine babesiosis and anaplasmosis. Vet. Parasitol. 2011, 180, 109–125. [Google Scholar] [CrossRef]
- Rodríguez-Vivas, R.I.; Grisi, L.; Pérez de León, A.A.; Villela, H.S.; Torres-Acosta, J.F.D.J.; Fragoso Sánchez, H.; Romero Salas, D.; Rosario Cruz, R.; Saldierna, F.; García Carrasco, D. Potential economic impact assessment for cattle parasites in Mexico. Rev. Rev. Mex. Cienc. Pec. 2017, 8, 61–74. [Google Scholar] [CrossRef]
- Bock, R.E.; de Vos, A.J. lmmunity following use of Australian tick fever vaccine: A review of the evidence. Aust. Vet. J. 2001, 79, 832–839. [Google Scholar] [CrossRef]
- Good, M.F.; Stanisic, D.I. Whole parasite vaccines for the asexual blood stages of Plasmodium. Immunol. Rev. 2020, 293, 270–282. [Google Scholar] [CrossRef]
- Al-Nazal, H.A.; Cooper, E.; Ho, M.F.; Eskandari, S.; Majam, V.; Giddam, A.K.; Hussein, W.M.; Islam, T.; Skwarczynski, M.; Toth, I.; et al. Pre-clinical evaluation of a whole-parasite vaccine to control human babesiosis. Cell Host Microbe 2021, 29, 894–903. [Google Scholar] [CrossRef]
- Giddam, A.K.; Reiman, J.M.; Zaman, M.; Skwarczynski, M.; Toth, I.; Good, M.F. A semi-synthetic whole parasite vaccine designed to protect against blood stage malaria. Acta Biomater. 2016, 44, 295–303. [Google Scholar] [CrossRef]
- Ssemaganda, A.; Giddam, A.K.; Zaman, M.; Skwarczynski, M.; Toth, I.; Stanisic, D.I.; Good, M.F. Induction of Plasmodium-Specific Immune Responses Using Liposome-Based Vaccines. Front. Immunol. 2019, 10, 135. [Google Scholar] [CrossRef]
- Callow, L.L.; Mellors, L.T. A new vaccine for Babesia argentina infection prepared in splenectomised calves. Aust. Vet. J. 1966, 42, 464–465. [Google Scholar] [CrossRef]
- Stahl, P.; Poinsignon, Y.; Pouedras, P.; Ciubotaru, V.; Berry, L.; Emu, B.; Krause, P.J.; Ben Mamoun, C.; Cornillot, E. Case report of the patient source of the Babesia microti R1 reference strain and implications for travelers. J. Travel Med. 2018, 25, tax073. [Google Scholar] [CrossRef] [PubMed]
- Al-Nazal, H.; Low, L.M.; Kumar, S.; Good, M.F.; Stanisic, D.I. A vaccine for human babesiosis: Prospects and feasibility. Trends Parasitol. 2022, 38, 904–918. [Google Scholar] [CrossRef]
- Ssemaganda, A.; Giddam, A.K.; Low, L.M.; Liu, X.Q.; Ho, M.-F.; Zaman, M.; Hussein, W.M.; Skwarczynski, M.; Toth, I.; Stanisic, D.I.; et al. Mannosylated liposomes formulated with whole parasite P. falciparum blood-stage antigens are highly immunogenic in mice. Vaccine 2020, 38, 1494–1504. [Google Scholar] [CrossRef]
- Rodriguez, M.; Alhassan, A.; Ord, R.L.; Cursino-Santos, J.R.; Singh, M.; Gray, J.; Lobo, C.A. Identification and characterization of the RouenBd1987 Babesia divergens Rhopty-Associated Protein 1. PLoS ONE. 2014, 9, e107727. [Google Scholar] [CrossRef]
- Stanisic, D.; Liu, X.Q.; De, S.L.; Batzloff, M.R.; Forbes, T.; Davis, C.B.; Sekuloski, S.; Chavchich, M.; Chung, W.; Trenholme, K.; et al. Development of cultured Plasmodium falciparum blood-stage malaria cell banks for early phase in vivo clinical trial assessment of anti-malaria drugs and vaccines. Malar. J. 2015, 14, 143. [Google Scholar] [CrossRef]
- Stanisic, D.I.; Fink, J.; Mayer, J.; Coghill, S.; Gore, L.; Liu, X.Q.; El-Deeb, I.; Rodriguez, I.B.; Powell, J.; Willemsen, N.M.; et al. Vaccination with chemically attenuated Plasmodium falciparum asexual blood-stage parasites induces parasite-specific cellular immune responses in malaria-naïve volunteers: A pilot study. BMC Med. 2018, 16, 184. [Google Scholar] [CrossRef] [PubMed]
- Man, S.; Fu, Y.; Guan, Y.; Feng, M.; Qiao, K.; Li, X.; Gao, H.; Cheng, X. Evaluation of a Major Surface Antigen of Babesia microti Merozoites as a Vaccine Candidate against Babesia Infection. Front. Microbiol. 2017, 8, 2545. [Google Scholar] [CrossRef]
- Li, M.; Ao, Y.; Guo, J.; Nie, Z.; Liu, Q.; Yu, L.; Luo, X.; Zhan, X.; Zhao, Y.; Wang, S.; et al. Surface Antigen 1 Is a Crucial Secreted Protein That Mediates Babesia microti Invasion Into Host Cells. Front. Microbiol. 2020, 10, 3046. [Google Scholar] [CrossRef]
- Elton, C.M.; Rodriguez, M.; Ben Mamoun, C.; Lobo, C.A.; Wright, G.J. A library of recombinant Babesia microti cell surface and secreted proteins for diagnostics discovery and reverse vaccinology. Int. J. Parasitol. 2019, 49, 115–125. [Google Scholar] [CrossRef]
- Zhou, X.; Huang, J.-L.; Shen, H.-M.; Xu, B.; Chen, J.-H.; Zhou, X.-N. Immunomics analysis of Babesia microti protein markers by high-throughput screening assay. Ticks Tick-Borne Dis. 2018, 9, 1468–1474. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Huang, J.; Xu, B.; Chen, J.; Dai, J.; Zhou, X. Babesia microti Protein BmSP44 Is a Novel Protective Antigen in a Mouse Model of Babesiosis. Front. Immunol. 2020, 11, 1437. [Google Scholar] [CrossRef]
- Munkhjargal, T.; Yokoyama, N.; Igarashi, I. Recombinant methionine aminopeptidase protein of Babesia microti: Immunobi-ochemical characterization as a vaccine candidate against human babesiosis. Parasitol. Res. 2016, 115, 3669–3676. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Aboge, G.; Jia, H.; Goo, Y.K.; Ooka, H.; Yamagishi, J.; Nishikawa, Y.; Yokoyama, N.; Igarashi, I.; Kawazu, S.I.; et al. Molecular and immunological characterization of Babesia gibsoni and Babesia microti heat shock protein-70. Parasite Immunol. 2009, 31, 328–340. [Google Scholar] [CrossRef]
- Gunning, P.W.; Ghoshdastider, U.; Whitaker, S.; Popp, D.; Robinson, R.C. The evolution of compositionally and functionally distinct actin filaments. J. Cell Sci. 2015, 128, 2009–2019. [Google Scholar] [CrossRef]
- Munkhjargal, T.; Aboge, G.O.; Ueno, A.; Aboulaila, M.; Yokoyama, N.; Igarashi, I. Identification and characterization of profilin antigen among Babesia species as a common vaccine candidate against babesiosis. Exp. Parasitol. 2016, 166, 29–36. [Google Scholar] [CrossRef]
- Tyler, J.S.; Treeck, M.; Boothroyd, J.C. Focus on the ringleader: The role of AMA1 in apicomplexan invasion and replication. Trends Parasitol. 2011, 27, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Moitra, P.; Zheng, H.; Anantharaman, V.; Banerjee, R.; Takeda, K.; Kozakai, Y.; Lepore, T.; Krause, P.J.; Aravind, L.; Kumar, S. Expression, Purification, and Biological Characterization of Babesia microti Apical Membrane Antigen 1. Infect. Immun. 2015, 83, 3890–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Yu, L.; Efstratiou, A.; Moumouni, P.; Liu, M.; Guo, H.; Gao, Y.; Cao, S.; Zhou, M.; Li, J.; et al. Evaluation of the protective effect of a prime-boost strategy with plasmid DNA followed by recombinant adenovirus expressing BmAMA1 as vaccines against Babesia microti infection in hamster. Acta Parasitol. 2018, 63, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Ord, R.L.; Rodriguez, M.; Cursino-Santos, J.R.; Hong, H.; Singh, M.; Gray, J.; Lobo, C.A. Identification and Characterization of the Rhoptry Neck Protein 2 in Babesia divergens and B. microti. Infect. Immun. 2016, 84, 1574–1584. [Google Scholar] [CrossRef]
- Wang, G.; Efstratiou, A.; Moumouni, P.F.A.; Liu, M.; Jirapattharasate, C.; Guo, H.; Gao, Y.; Cao, S.; Zhou, M.; Suzuki, H.; et al. Expression of truncated Babesia microti apical membrane protein 1 and rhoptry neck protein 2 and evaluation of their protective efficacy. Exp. Parasitol. 2017, 172, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.C.; Yang, C.L.; Hu, W.; Song, P.; Xu, B.; Lu, Y.; Ai, L.; Chu, Y.H.; Chen, M.X.; Chen, J.X.; et al. Molecular Characterization and Immunological Evaluation of Truncated Babesia microti Rhoptry Neck Protein 2 as a Vaccine Candidate. Front. Immunol. 2021, 12, 616343. [Google Scholar] [CrossRef] [PubMed]
- Sondgeroth, K.S.; McElwain, T.F.; Allen, A.J.; Chen, A.V.; Lau, A.O. Loss of neurovirulence is associated with reduction of cerebral capillary sequestration during acute Babesia bovis infection. Parasites Vectors 2013, 6, 181. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Cornillot, E.; McCracken, C.; Usmani-Brown, S.; Dwivedi, A.; Ifeonu, O.O.; Crabtree, J.; Gotia, H.T.; Virji, A.Z.; Reynes, C.; et al. Genome-wide diversity and gene expression profiling of Babesia microti isolates identify polymorphic genes that mediate host-pathogen interactions. Sci. Rep. 2016, 6, 35284. [Google Scholar] [CrossRef]
- Lemieux, J.E.; Tran, A.D.; Freimark, L.; Schaffner, S.F.; Goethert, H.; Andersen, K.G.; Bazner, S.; Li, A.; McGrath, G.; Sloan, L.; et al. A global map of genetic diversity in Babesia microti reveals strong population structure and identifies variants associated with clinical relapse. Nat. Microbiol. 2016, 1, 16079. [Google Scholar] [CrossRef]
- Verma, N.; Puri, A.; Essuman, E.; Skelton, R.; Anantharaman, V.; Zheng, H.; White, S.; Gunalan, K.; Takeda, K.; Bajpai, S.; et al. Antigen Discovery, Bioinformatics and Biological Characterization of Novel Immunodominant Babesia microti Antigens. Sci. Rep. 2020, 10, 9598. [Google Scholar] [CrossRef]
- Xu, B.; Liu, X.-F.; Cai, Y.-C.; Huang, J.-L.; Zhang, R.-X.; Chen, J.-H.; Cheng, X.-J.; Zhou, X.; Xu, X.-N.; Zhou, Y.; et al. Screening for biomarkers reflecting the progression of Babesia microti infection. Parasites Vectors 2018, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Magni, R.; Luchini, A.; Liotta, L.; Molestina, R.E. Analysis of the Babesia microti proteome in infected red blood cells by a combination of nanotechnology and mass spectrometry. Int. J. Parasitol. 2019, 49, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Jaijyan, D.K.; Govindasamy, K.; Singh, J.; Bhattacharya, S.; Singh, A.P. Establishment of a stable transfection method in Babesia microti and identification of a novel bidirectional promoter of Babesia microti. Sci. Rep. 2020, 10, 15614. [Google Scholar] [CrossRef]
- Wei, N.; Lu, J.; Gong, H.; Xu, Z.; Zhang, H.; Cui, L.; Zhou, J.; Lin, Z. Inclusion of PD-L1 into a recombinant profilin antigen enhances immunity against Babesia microti in a murine model. Ticks Tick-Borne Dis. 2020, 11, 101446. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.G.; Alzan, H.F.; Rathinasamy, V.A.; Cooke, B.M.; Dellagostin, O.A.; Barletta, R.G.; Suarez, C.E. Harnessing Mycobacterium bovis BCG Trained Immunity to Control Human and Bovine Babesiosis. Vaccines 2022, 10, 123. [Google Scholar] [CrossRef]
- Clark, I.A.; Allison, A.C.; Cox, F.E. Protection of mice against Babesia, and Plasmodium with BCG. Nature 1976, 259, 309–311. [Google Scholar] [CrossRef]
- Clark, I.; Wills, E.J.; Richmond, J.; Allison, A.C. Suppression of babesiosis in BCG-infected mice and its correlation with tumor inhibition. Infect. Immun. 1977, 17, 430–438. [Google Scholar] [CrossRef]
- Underwood, W.J.; Blauwiekel, R.; Delano, M.L.; Gillesby, R.; Mischler, S.A.; Schoell, A. Biology and Diseases of Ruminants (Sheep, Goats, and Cattle). Lab. Anim. Med. 2015, 15, 623–694. [Google Scholar] [CrossRef]
- Spickler, A.R. Bovine Babesiosis; CFSPH: Ames, IA, USA, 2018. [Google Scholar]
- Florin-Christensen, M.; Suarez, C.E.; Rodriguez, A.E.; Flores, D.A.; Schnittger, L. Vaccines against bovine babesiosis: Where we are now and possible roads ahead. Parasitology 2004, 141, 1563–1592. [Google Scholar] [CrossRef]
- Mazuz, M.L.; Laughery, J.M.; Lebovitz, B.; Yasur-Landau, D.; Rot, A.; Bastos, R.G.; Edery, N.; Fleiderovitz, L.; Levi, M.M.; Suarez, C.E. Experimental Infection of Calves with Transfected Attenuated Babesia bovis Expressing the Rhipicephalus microplus Bm86 Antigen and eGFP Marker: Preliminary Studies towards a Dual Anti-Tick/Babesia Vaccine. Pathogens 2021, 10, 135. [Google Scholar] [CrossRef] [PubMed]
- Dalgliesh, R.J.; Jorgensen, W.K.; de Vos, A.J. New Australian vaccines for the control of babesiosis and anaplasmosis in the world cattle trade. Trop. Anim. Health Prod. 1990, 22, 44–52. [Google Scholar] [CrossRef]
- Timms, P.; Stewart, N.P.; De Vos, A.J. Study of virulence and vector transmission of Babesia bovis by use of cloned parasite lines. Infect. Immun. 1990, 58, 2171–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangold, A.J.; Aguirre, D.H.; Cafrune, M.M.; de Echaide, S.; Guglielmone, A.A. Evaluation of the infectivity of a vaccinal and a pathogenic Babesia bovis strain from Argentina to Boophilus microplus. Vet. Parasitol. 1993, 51, 143–148. [Google Scholar] [CrossRef]
- Shkap, V.; Leibovitz, B.; Krigel, Y.; Hammerschlag, J.; Marcovics, A.; Fish, L.; Molad, T.; Savitsky, I.; Mazuz, M. Vaccination of older Bos taurus bulls against bovine babesiosis. Vet. Parasitol. 2005, 129, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Standfast, N.F.; Bock, R.E.; Wiecek, M.M.; Devos, A.J.; Jorgensen, W.K.; Kingston, T.G. Overcoming constraints to meeting increased demand for Babesia bigemina vaccine in Australia. Vet. Parasitol. 2003, 115, 213–222. [Google Scholar] [CrossRef]
- Gimenez, A.M.; Françoso, K.S.; Ersching, J.; Icimoto, M.Y.; Oliveira, V.; Rodriguez, A.E.; Schnittger, L.; Florin-Christensen, M.; Rodrigues, M.M.; Soares, I.S. A recombinant multi-antigen vaccine formulation containing Babesia bovis merozoite surface antigens MSA-2a1, MSA-2b and MSA-2c elicits invasion-inhibitory antibodies and IFN-γ producing cells. Parasites Vectors 2016, 9, 577. [Google Scholar] [CrossRef]
- Shkap, V.; de Vos, A.J.; Zweygarth, E.; Jongejan, F. Attenuated vaccines for tropical theileriosis, babesiosis and heartwater: The continuing necessity. Trends Parasitol. 2007, 23, 420–426. [Google Scholar] [CrossRef]
- Suarez, C.E.; Florin-Christensen, M.; Hines, S.A.; Palmer, G.H.; Brown, W.C.; McElwain, T.F. Characterization of Allelic Variation in the Babesia bovis Merozoite Surface Antigen 1 (MSA-1) Locus and Identification of a Cross-Reactive Inhibition-Sensitive MSA-1 Epitope. Infect. Immun. 2000, 68, 6865–6870. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Ruiz, M.; Mejia-López, S.; Pérez-Serrano, R.M.; Zaldívar-Lelo de Larrea, G.; Ganzinelli, S.; Florin-Christensen, M.; Suarez, C.E.; Hernández-Ortiz, R.; Mercado-Uriostegui, M.A.; Rodríguez-Torres, A.; et al. Babesia bovis AMA-1, MSA-2c and RAP-1 contain conserved B and T-cell epitopes, which generate neutralizing antibodies and a long-lasting Th1 immune response in vaccinated cattle. Vaccine 2022, 40, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Cuy-Chaparro, L.; Ricaurte-Contreras, L.A.; Bohórquez, M.D.; Arévalo-Pinzón, G.; Barreto-Santamaria, A.; Pabón, L.; Reyes, C.; Moreno-Pérez, D.A.; Patarroyo, M.A. Identification of Babesia bovis MSA-1 functionally constraint regions capable of binding to bovine erythrocytes. Vet. Parasitol. 2022, 312, 109834. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.E.; Florin-Christensen, M.; Flores, D.A.; Echaide, I.; Suarez, C.E.; Schnittger, L. The glycosylphosphatidylino-sitol-anchored protein repertoire of Babesia bovis and its significance for erythrocyte invasion. Ticks Tick-Borne Dis. 2014, 5, 343–348. [Google Scholar] [CrossRef]
- Ramos, C.A.; Araújo, F.R.; Souza, I.I.; Oliveira, R.H.; Elisei, C.; Soares, C.O.; Sacco, A.M.; Rosinha, G.M.; Alves, L.C. Molecular and antigenic characterisation of ribosomal phosphoprotein P0 from Babesia bovis. Mem. Inst. Oswaldo. Cruz 2009, 104, 998–1002. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, S.D.; Palmer, G.H.; McElwain, T.F.; McGuire, T.C.; Ruef, B.J.; Chitko-McKown, M.G.; Brown, W.C. CD4+ T-helper lymphocyte responses against Babesia bigemina rhoptry-associated protein I. Infect. Immun. 1996, 64, 2079–2087. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda, J.; McElwain, T.F.; Stiller, D.; Palmer, G.H. Babesia bovis merozoite surface antigen 1 and rhoptry-associated protein 1 are expressed in sporozoites, and specific antibodies inhibit sporozoite attachment to erythrocytes. Infect. Immun. 2002, 70, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Terkawi, M.A.; Ratthanophart, J.; Salama, A.; AbouLaila, M.; Asada, M.; Ueno, A.; Alhasan, H.; Guswanto, A.; Masatani, T.; Yokoyama, N.; et al. Molecular Characterization of a New Babesia bovis Thrombospondin-Related Anonymous Protein (BbTRAP2). PLoS ONE 2013, 8, e83305. [Google Scholar] [CrossRef]
- Fus-Kujawa, A.; Prus, P.; Bajdak-Rusinek, K.; Teper, P.; Gawron, K.; Kowalczuk, A.; Sieron, A.L. An Overview of Methods and Tools for Transfection of Eukaryotic Cells in vitro. Front. Bioeng. Biotechnol. 2021, 9, 498–504. [Google Scholar] [CrossRef]
- Suarez, C.E.; McElwain, T.F. Transient transfection of purified Babesia bovis merozoites. Exp. Parasitol. 2008, 118, 498–504. [Google Scholar] [CrossRef]
- Asada, M.; Tanaka, M.; Goto, Y.; Yokoyama, N.; Inoue, N.; Kawazu, S. Stable expression of green fluorescent protein and targeted disruption of thioredoxin peroxidase-1 gene in Babesia bovis with the WR99210/dhfr selection system. Mol. Biochem. Parasitol. 2012, 181, 162–170. [Google Scholar] [CrossRef]
- Suarez, C.E.; McElwain, T.F. Stable expression of a GFP-BSD fusion protein in Babesia bovis merozoites. Int. J. Parasitol. 2009, 39, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.E.; Laughery, J.M.; Schneider, D.A.; Sondgeroth, K.S.; McElwain, T.F. Acute and persistent infection by a transfected Mo7 strain of Babesia bovis. Mol. Biochem. Parasitol. 2012, 185, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Oldiges, D.P.; Laughery, J.M.; Tagliari, N.J.; Filho, R.V.L.; Davis, W.C.; da Silva Vaz, I., Jr.; Termignoni, C.; Knowles, D.P.; Suarez, C.E. Transfected Babesia bovis Expressing a Tick GST as a Live Vector Vaccine. PLoS Negl. Trop. Dis. 2016, 10, e0005152. [Google Scholar] [CrossRef]
- Kumar, B.; Murugan, K.; Ray, D.D.; Ghosh, S. Efficacy of rBm86 against Rhipicephalus (Boophilus) microplus (IVRI-I line) and Hyalomma anatolicum anatolicum (IVRI-II line) infestations on bovine calves. Parasitol. Res. 2012, 111, 629–635. [Google Scholar] [CrossRef]
- Montenegro, V.N.; Paoletta, M.S.; Jaramillo Ortiz, J.M.; Suarez, C.E.; Wilkowsky, S.E. Identification and characterization of a Babesia bigemina thrombospondin-related superfamily member, TRAP-1: A novel antigen containing neutralizing epitopes involved in merozoite invasion. Parasit Vectors 2020, 13, 602. [Google Scholar] [CrossRef]
- Hernández-Silva, D.J.; Valdez-Espinoza, U.M.; Mercado-Uriostegui, M.A.; Aguilar-Tipacamú, G.; Ramos-Aragón, J.A.; Hernández-Ortiz, R.; Ueti, M.; Mosqueda, J. Immunomolecular Characterization of MIC-1, a Novel Antigen in Babesia bigemina, Which Contains Conserved and Immunodominant B-Cell Epitopes that Induce Neutralizing Antibodies. Vet. Sci. 2018, 5, 32. [Google Scholar] [CrossRef]
- Brown, W.C.; McElwain, T.F.; Palmer, G.H.; Chantler, S.E.; Estes, D.M. Bovine CD4(+) T-lymphocyte clones specific for rhoptry-associated protein 1 of Babesia bigemina stimulate enhanced immunoglobulin G1 (IgG1) and IgG2 synthesis. Infect. Immun. 1999, 67, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.C.; McElwain, T.F.; Hötzel, I.; Suarez, C.E.; Palmer, G.H. Helper T-cell epitopes encoded by the Babesia bigemina rap-1 gene family in the constant and variant domains are conserved among parasite strains. Infect. Immun. 1998, 66, 1561–1569. [Google Scholar] [CrossRef]
- Camacho-Nuez, M.; Hernández-Silva, D.J.; Castañeda-Ortiz, E.J.; Paredes-Martínez, M.E.; Rocha-Martínez, M.K.; Alvarez-Sánchez, M.E.; Mercado-Curiel, R.F.; Aguilar-Tipacamu, G.; Mosqueda, J. Hap2, a novel gene in Babesia bigemina is expressed in tick stages, and specific antibodies block zygote formation. Parasites Vectors 2017, 10, 568. [Google Scholar] [CrossRef]
- Rauf, U.; Suleman, M.; Abid, A.; Jamil, H.; Menghwar, H.; Durrani, A.Z.; Rashid, M.I.; Akbar, H. Humoral and Cell-Mediated Immune Response Validation in Calves after a Live Attenuated Vaccine of Babesia bigemina. Pathogens 2020, 9, 936. [Google Scholar] [CrossRef]
- Silva, M.G.; Knowles, D.P.; Mazuz, M.L.; Cooke, B.M.; Suarez, C.E. Stable transformation of Babesia bigemina and Babesia bovis using a single transfection plasmid. Sci. Rep. 2018, 8, 6096. [Google Scholar] [CrossRef]
- Antunes, S.; Merino, O.; Mosqueda, J.; Moreno-Cid, J.A.; Bell-Sakyi, L.; Fragkoudis, R.; Weisheit, S.; de la Lastra, J.M.P.; Alberdi, P.; Domingos, A.; et al. Tick capillary feeding for the study of proteins involved in tick-pathogen interactions as potential antigens for the control of tick infestation and pathogen infection. Parasites Vectors 2014, 7, 42. [Google Scholar] [CrossRef]
- Nie, Z.; Ao, Y.; Wang, S.; Shu, X.; Li, M.; Zhan, X.; Yu, L.; An, X.; Sun, Y.; Guo, J.; et al. Erythrocyte Adhesion of Merozoite Surface Antigen 2c1 Expressed During Extracellular Stages of Babesia orientalis. Front. Immunol. 2021, 12, 623492. [Google Scholar] [CrossRef]
- Montero, E.; Rodriguez, M.; Oksov, Y.; Lobo, C.A. Babesia divergens Apical Membrane Antigen 1 and Its Interaction with the Human Red Blood Cell. Infect. Immun. 2009, 77, 4783–4793. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, S.A.E.-S.; Rizk, M.A.; Eldoumani, H.; Sorour, S.S.; Terkawi, M.A.; AbouLaila, M.; Igarashi, I.; Sayed-Ahmed, M.Z. Identification and Characterization of P0 Protein as a Vaccine Candidate against Babesia divergens, Blood Parasite of Veterinary and Zoonotic Importance. Front. Vet. Sci. 2022, 8, 795906. [Google Scholar] [CrossRef]


{kind=link}
| Species | Protection for Animals | Protection for Humans |
|---|---|---|
| B. microti | X | BMSA BmSA1 BmSP44 Babesia microti methionine aminopeptidase protein 1 B. microti heat-shock protein-70 BmPROF BboPROF BbigPROF BmAMA-1 BmRON2 N-terminal and C-terminal fragments of BmRON2 Bm2D41 BmSERA1 BmMCFPR1 BmPiβS1 BmBAHCS1 |
| B. bovis | MSA-1/MSA-2 PHOSPHOPROTEIN P0 RAP-1 BpTRAP2 | BboPROF |
| B. bigemina | BbiTRAP-1 MIC-1 RAP-1 HAP2/GCS1 TROSPA | BbigPROF |
| B. divergens | BdAMA1 BdP0 | BdAMA1 BdP0 |
| B. orientalis | BoMSA-2c1 | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jerzak, M.; Gandurski, A.; Tokaj, M.; Stachera, W.; Szuba, M.; Dybicz, M. Advances in Babesia Vaccine Development: An Overview. Pathogens 2023, 12, 300. https://doi.org/10.3390/pathogens12020300
Jerzak M, Gandurski A, Tokaj M, Stachera W, Szuba M, Dybicz M. Advances in Babesia Vaccine Development: An Overview. Pathogens. 2023; 12(2):300. https://doi.org/10.3390/pathogens12020300
Chicago/Turabian StyleJerzak, Michał, Albert Gandurski, Marta Tokaj, Weronika Stachera, Magdalena Szuba, and Monika Dybicz. 2023. "Advances in Babesia Vaccine Development: An Overview" Pathogens 12, no. 2: 300. https://doi.org/10.3390/pathogens12020300