
A Review on the Immunological Response against Trypanosoma cruzi
 , , , , ,
, , , , , {kind=link}
{kind=link}
Abstract
:1. Introduction
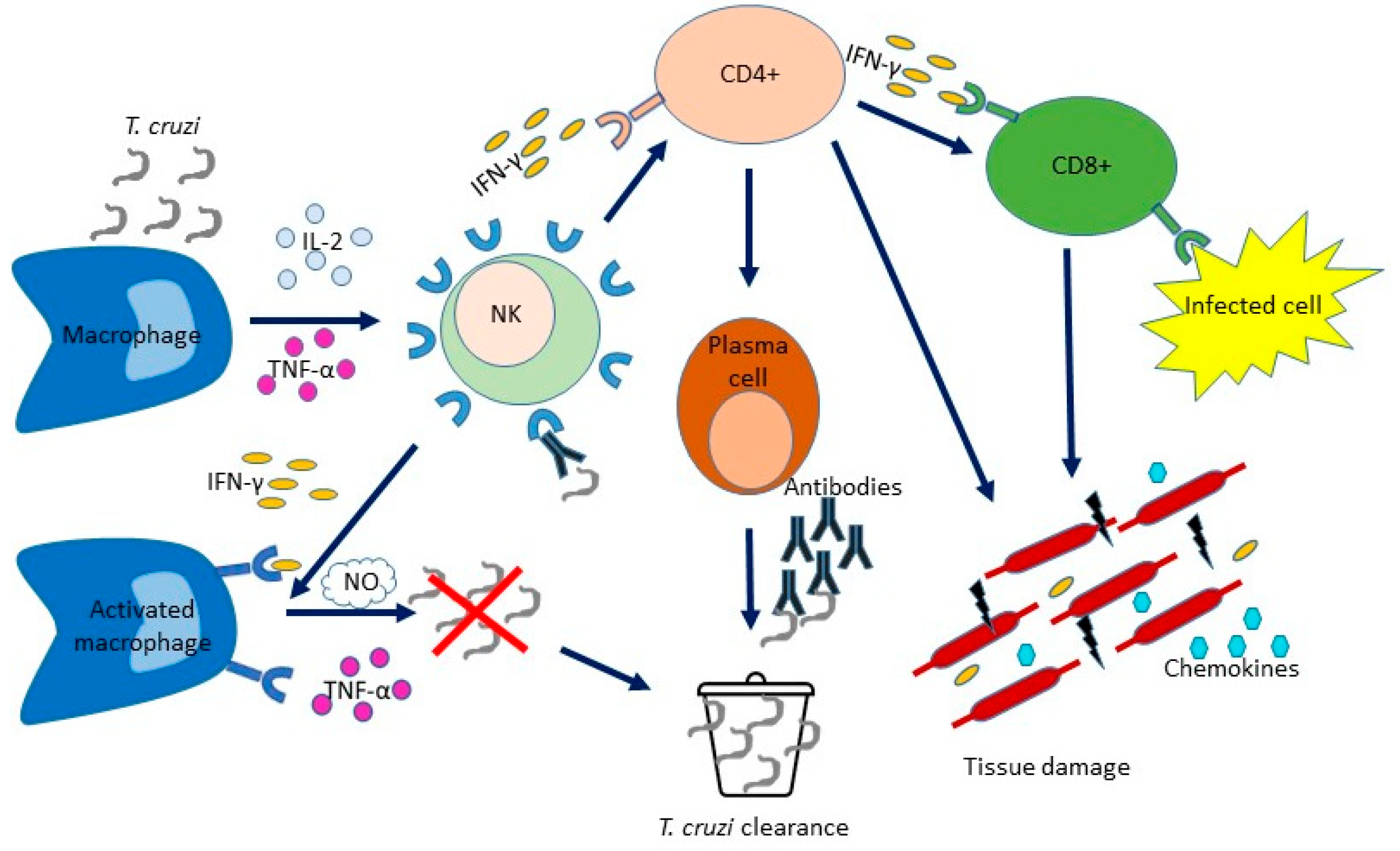
2. The Innate Immune Response to Trypanosoma cruzi
3. The Adaptative Immune Response to Trypanosoma cruzi
4. Toll-like Receptors
5. Virulence Factors
6. Authophagy
7. Therapeutic Approaches
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deane, L.M. Animal reservoirs of Trypanosoma cruzi in Brazil. Rev. Bras. Malariol. Doenças Trop. 1964, 16, 27–48. [Google Scholar]
- Lent, H.; Wygodzinsky, P. Revision of the Triatominae (Hemiptera Reduviidae), and their significance as vector of Chagas disease. Bull. Am. Mus. Nat. History 1979, 163, 123–520. [Google Scholar]
- Galvão, C.; Carcavallo, R.; Rocha, D.S.; Jurberg, J. A checklist of the current valid species of the subfamily Triatominae Jeannel; 1919 (Hemiptera; Reduviidae) and their geographical distribution; with nomenclatural and taxonomic notes. Zootaxa 2003, 202, 1–36. [Google Scholar] [CrossRef]
- Schmunis, G.A. Prevention of transfusional Trypanosoma cruzi infection in Latin America. Memórias Inst. Oswaldo Cruz 1999, 94 (Suppl. 1), 93–101. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Montgomery, S.P.; Katz, L.; Caglioti, S.; Stramer, S.L. Chagas disease and the US blood supply. Curr. Opin. Infect. Dis. 2008, 21, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Pereira, K.S.; Schmidt, F.L.; Guaraldo, A.M.; Franco, R.M.; Dias, V.L.; Passos, L.A. Chagas disease as a foodborne illness. J. Food Prot. 2009, 72, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Tyler, K.M.; Engman, D.M. The life-cycle of Trypanosoma cruzi. In American Trypanosomiasis; Tyler, K.M., Miles, M.A., Eds.; World class parasites; Kluwer Academic Publishers: Boston, MA, USA, 2003; Volume 7, pp. 1–11. [Google Scholar]
- Macedo, A.M.; Machado, C.R.; Oliveira, R.P.; Pena, S.D. Trypanosoma cruzi: Genetic structure of populations and relevance of genetic variability to the pathogenesis of Chagas disease. Memórias Inst. Oswaldo Cruz 2004, 99, 1–12. [Google Scholar] [CrossRef]
- Manoel-Caetano, F.S.; Silva, A.E. Implications of genetic variability of Trypanosoma cruzi for the pathogenesis of Chagas disease. Cad. Saúde Pública 2007, 23, 2263–2274. [Google Scholar] [CrossRef]
- Nobrega, A.A.; Garcia, M.H.; Tatto, E.; Obara, M.T.; Costa, E.; Sobel, J.; Araujo, W.N. Oral transmission of Chagas disease by consumption of acai palm fruit; Brazil. Emerg. Infect. Dis. 2009, 15, 653–655. [Google Scholar] [CrossRef]
- Patel, S.; Sethi, A. Imported tropical diseases. Dermatol. Ther. 2009, 22, 538–549. [Google Scholar] [CrossRef]
- Lupi, O.; Bartlett, B.L.; Haugen, R.N.; Dy, L.C.; Sethi, A.; Klaus, S.N.; Machado Pinto, J.; Bravo, F.; Tyring, S.K. Tropical dermatology: Tropical diseases caused by protozoa. J. Am. Acad. Dermatol. 2009, 60, 897–925. [Google Scholar] [CrossRef]
- WHO. Control of Chagas Disease; Second report of the WHO Expert Committee; Technical report series no 905; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Bittencourt, A.L. Congenital Chagas disease. Am. J. Dis. Child. 1976, 130, 97–103. [Google Scholar] [CrossRef]
- Freilij, H.; Altcheh, J. Congenital Chagas’ disease: Diagnostic and clinical aspects. Clin. Infect. Dis. 1995, 21, 551–555. [Google Scholar] [CrossRef]
- Teixeira, A.R.; Nascimento, R.J.; Sturm, N.R. Evolution and pathology in Chagas disease—A review. Memórias Inst. Oswaldo Cruz 2006, 101, 463–491. [Google Scholar] [CrossRef]
- Martinez, F.; Perna, E.; Perrone, S.V.; Liprandi, A.S. Chagas Disease and Heart Failure: An Expanding Issue Worldwide. Eur. Cardiol. 2019, 14, 82–88. [Google Scholar] [CrossRef]
- Bonney, K.M.; Engman, D.M. Chagas heart disease pathogenesis: One mechanism or many? Curr. Mol. Med. 2008, 8, 510–518. [Google Scholar] [CrossRef]
- Nagajyothi, F.; Machado, F.S.; Burleigh, B.A.; Jelicks, L.A.; Scherer, P.E.; Mukherjee, S.; Lisanti, M.P.; Weiss, L.M.; Garg, N.J.; Tanowitz, H.B. Mechanisms of Trypanosoma cruzi persistence in Chagas disease. Cell. Microbiol. 2012, 14, 634–643. [Google Scholar] [CrossRef]
- Cardillo, F.; Postol, E.; Nihei, J.; Aroeira, L.S.; Nomizo, A.; Mengel, J. B cells modulate T cells so as to favour T helper type 1 and CD8+ T-cell responses in the acute phase of Trypanosoma cruzi infection. Immunology 2007, 122, 584–595. [Google Scholar] [CrossRef]
- Rezende-Oliveira, K.; Sarmento, R.R.; Rodrigues, V., Jr. Production of cytokine and chemokines by human mononuclear cells and whole blood cells after infection with Trypanosoma cruzi. Rev. Soc. Bras. Med. Trop. 2012, 45, 45–50. [Google Scholar] [CrossRef]
- Pinho, R.T.; da Silva, W.S.; de Castro Cortes, L.M.; da Silva Vasconcelos Sousa, P.; de Araujo Soares, R.O.; Alves, C.R. Production of MMP-9 and inflammatory cytokines by Trypanosoma cruzi infected macrophages. Exp. Parasitol. 2014, 147, 72–80. [Google Scholar] [CrossRef]
- Acevedo, G.R.; Girard, M.C.; Gómez, K.A. The unsolved jigsaw puzzle of the immune response in Chagas disease. Front. Immunol. 2018, 9, 1929. [Google Scholar] [CrossRef] [PubMed]
- Gurung, P.; Kanneganti, T.D. Immune responses against protozoan parasites: A focus on the emerging role of Nod-like receptors. Cell. Mol. Life Sci. 2016, 73, 3035–3051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noel, W.; Raes, G.; Hassanzadeh Ghass, G.; De Baetselier, P.; Beschin, A. Alternatively activated macrophages during parasite infections. Trends. Parasitol. 2004, 20, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.S.; Machado, F.S.; Martins, G.A. The role of nitric oxide in the pathogenesis of Chagas disease. Front. Biosci. 2003, 8, s314–s325. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends. Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Camargo, M.; Almeida, I.; Pereira, M.; Ferguson, M.; Travassos, L.; Gazzinelli, R. Glycosylphosphatidylinositol-anchored mucin-like glycoproteins isolated from Trypanosoma cruzi trypomastigotes initiate the synthesis of proinflammatory cytokines by macrophages. J. Immunol. 1997, 158, 5890–5901. [Google Scholar] [CrossRef]
- Campos, M.A.; Almeida, I.C.; Takeuchi, O.; Akira, S.; Valente, E.P.; Procópio, D.O.; Travassos, L.R.; Smith, J.A.; Golenbock, D.T.; Gazzinelli, R.T. Activation of toll-like receptor-2 by glycosylphosphatidylinositol anchors from a protozoan parasite. J. Immunol. 2001, 167, 416–423. [Google Scholar] [CrossRef]
- Tarleton, R.L. CD8+ T cells in Trypanosoma cruzi infection. Semin. Immunopathol. 2015, 37, 233–238. [Google Scholar] [CrossRef]
- da Costa, T.A.; Silva, M.V.; Mendes, M.T.; Carvalho-Costa, T.M.; Batista, L.R.; Lages-Silva, E.; Rodrigues, V.; Oliveira, C.J.; Ramirez, L.E. Immunomodulation by Trypanosoma cruzi: Toward understanding the association of dendritic cells with infecting TcI and TcII populations. J. Immunol. Res. 2014, 2014, 962047. [Google Scholar] [CrossRef]
- Van Overtvelt, L.; Vanderheyde, N.; Verhasselt, V.; Ismaili, J.; De Vos, L.; Goldman, M.; Willems, F.; Vray, B. Trypanosoma cruzi infects human dendritic cells and prevents their maturation: Inhibition of cytokines, HLA-DR, and costimulatory molecules. Infect. Immun. 1999, 67, 4033–4040. [Google Scholar] [CrossRef]
- Sathler-Avelar, R.; Lemos, E.M.; Reis, D.D.; Medrano-Mercado, N.; Araújo-Jorge, T.C.; Antas, P.R.; Corrêa-Oliveira, R.; Teixeira-Carvalho, A.; Elói-Santos, S.M.; Favato, D.; et al. Phenotypic features of peripheral blood leucocytes during early stages of human infection with Trypanosoma cruzi. Scand. J. Immunol. 2003, 58, 655–663. [Google Scholar] [CrossRef]
- Lidani, K.C.F.; Bavia, L.; Ambrosio, A.R.; de Messias-Reason, I.J. The complement system: A prey of Trypanosoma cruzi. Front. Microbiol. 2017, 8, 607. [Google Scholar] [CrossRef]
- Zamboni, D.S.; Lima-Junior, D.S. Inflammasomes in host response to protozoan parasites. Immunol. Rev. 2015, 265, 156–171. [Google Scholar] [CrossRef]
- Kumar, S.; Tarleton, R.L. The relative contribution of antibody production and CD8+ T cell function to immune control of Trypanosoma cruzi. Parasite Immunol. 1998, 20, 207–216. [Google Scholar] [CrossRef]
- Sullivan, N.L.; Eickhoff, C.S.; Sagartz, J.; Hoft, D.F. Deficiency of antigenspecific B cells results in decreased Trypanosoma cruzi systemic but not mucosal immunity due to CD8 T cell exhaustion. J. Immunol. 2015, 194, 1806–1818. [Google Scholar] [CrossRef]
- Andrade, D.V.; Gollob, K.J.; Dutra, W.O. Acute Chagas disease: New global challenges for an old neglected disease. PLoS Negl. Trop. Dis. 2014, 8, e3010. [Google Scholar] [CrossRef]
- Dutra, W.O.; Menezes, C.A.; Villani, F.N.; da Costa, G.C.; da Silveira, A.B.; Reis, D.d.; Gollob, K.J. Cellular and genetic mechanisms involved in the generation of protective and pathogenic immune responses in human Chagas disease. Memórias Inst. Oswaldo Cruz 2009, 104, 208–218. [Google Scholar] [CrossRef]
- Souza, P.E.; Rocha, M.O.; Menezes, C.A.; Coelho, J.S.; Chaves, A.C.; Gollob, K.J.; Dutra, W.O. Trypanosoma cruzi infection induces differential modulation of costimulatory molecules and cytokines by monocytes and T cells from patients with indeterminate and cardiac Chagas’ disease. Infect. Immun. 2007, 75, 1886–1894. [Google Scholar] [CrossRef]
- de Araújo, F.F.; Corrêa-Oliveira, R.; Rocha, M.O.; Chaves, A.T.; Fiuza, J.A.; Fares, R.C.; Ferreira, K.S.; Nunes, M.C.; Keesen, T.S.; Damasio, M.P.; et al. Foxp3+CD25(high) CD4+ regulatory T cells from indeterminate patients with Chagas disease can suppress the effector cells and cytokines and reveal altered correlations with disease severity. Immunobiology 2012, 217, 768–777. [Google Scholar] [CrossRef]
- Ramstead, A.G.; Robison, A.; Blackwell, A.; Jerome, M.; Freedman, B.; Lubick, K.J.; Hedges, J.F.; Jutila, M.A. Roles of Toll-Like Receptor 2 (TLR2), TLR4, and MyD88 During Pulmonary Coxiella burnetii Infection. Infect. Immun. 2016, 84, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Torina, A.; Blanda, V.; Villari, S.; Piazza, A.; La Russa, F.; Grippi, F.; La Manna, M.P.; Di Liberto, D.; de la Fuente, J.; Sireci, G. Immune Response to Tick-Borne Hemoparasites: Host Adaptive Immune Response Mechanisms as Potential Targets for Therapies and Vaccines. Int. J. Mol. Sci. 2020, 21, 8813. [Google Scholar] [CrossRef] [PubMed]
- Sireci, G.; Badami, G.D.; Di Liberto, D.; Blanda, V.; Grippi, F.; Di Paola, L.; Guercio, A.; de la Fuente, J.; Torina, A. Recent Advances on the Innate Immune Response to Coxiella burnetii. Front. Cell. Infect. Microbiol. 2021, 11, 754455. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, A.; Guiñazu, N.; Giordanengo, L.; Cano, R.C.; Gea, S. The role of Toll-like receptors and adaptive immunity in the development of protective or pathological immune response triggered by the Trypanosoma cruzi protozoan. Future Microbiol. 2011, 6, 1521–1533. [Google Scholar] [CrossRef]
- Campos, M.A.; Gazzinelli, R.T. Trypanosoma cruzi and its components as exogenous mediators of inflammation recognized through Toll-like receptors. Mediat. Inflamm. 2004, 13, 139–143. [Google Scholar] [CrossRef]
- Tarleton, R.L. Immune system recognition of Trypanosoma cruzi. Curr. Opin. Immunol. 2007, 19, 430–434. [Google Scholar] [CrossRef]
- Kayama, H.; Takeda, K. The innate immune response to Trypanosoma cruzi infection. Microbes Infect. 2010, 12, 511–517. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Carrera-Silva, E.A.; Guinazu, N.; Pellegrini, A.; Cano, R.C.; Arocena, A.; Aoki, M.P.; Gea, S. Importance of TLR2 on hepatic immune and non-immune cells to attenuate the strong inflammatory liver response during Trypanosoma cruzi acute infection. PLoS Negl. Trop. Dis. 2010, 4, e863. [Google Scholar] [CrossRef]
- Carrera-Silva, E.A.; Carolina, C.R.; Natalia, G.; Pilar, A.M.; Andrea, P.; Gea, S. TLR2, TLR4 and TLR9 are differentially modulated in liver lethally injured from BALB/c and C57BL/6 mice during Trypanosoma cruzi acute infection. Mol. Immunol. 2008, 45, 3580–3588. [Google Scholar] [CrossRef]
- Cerbán, F.M.; Stempin, C.C.; Volpini, X.; Carrera Silva, E.A.; Gea, S.; Motran, C.C. Signaling pathways that regulate Trypanosoma cruzi infection and immune response. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165707. [Google Scholar] [CrossRef]
- Maganto-Garcia, E.; Punzon, C.; Terhorst, C.; Fresno, M. Rab5 activation by Toll-like receptor 2 is required for Trypanosoma cruzi internalization and replication in macrophages. Traffic 2008, 9, 1299–1315. [Google Scholar] [CrossRef] [Green Version]
- Ropert, C.; Ferreira, L.R.; Campos, M.A.; Procópio, D.O.; Travassos, L.R.; Ferguson, M.A.; Reis, L.F.; Teixeira, M.M.; Almeida, I.C.; Gazzinelli, R.T. Macrophage signaling by glycosylphosphatidylinositol-anchored mucin-like glycoproteins derived from Trypanosoma cruzi trypomastigotes. Microbes Infect. 2002, 4, 1015–1025. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Peixoto, J.R.; de Arruda, L.B.; Campos, M.A.; Gazzinelli, R.T.; Golenbock, D.T.; Akira, S.; Previato, J.O.; Mendonça-Previato, L.; Nobrega, A.; et al. Expression of functional TLR4 confers proinflammatory responsiveness to Trypanosoma cruzi glycoinositolphospholipids and higher resistance to infection with T. cruzi. J. Immunol. 2004, 173, 5688–5696. [Google Scholar] [CrossRef]
- Coelho, P.S.; Klein, A.; Talvani, A.; Coutinho, S.F.; Takeuchi, O.; Akira, S.; Silva, J.S.; Canizzaro, H.; Gazzinelli, R.T.; Teixeira, M.M. Glycosylphosphatidylinositol-anchored mucin-like glycoproteins isolated from Trypanosoma cruzi trypomastigotes induce in vivo leukocyte recruitment dependent on MCP-1 production by IFN-gamma-primed-macrophages. J. Leukoc. Biol. 2002, 71, 837–844. [Google Scholar] [CrossRef]
- Stahl, P.; Schwarz, R.T.; Debierre-Grockiego, F.; Meyer, T. Trypanosoma cruzi parasites fight for control of the JAK-STAT pathway by disarming their host. JAKSTAT 2015, 3, e1012964. [Google Scholar]
- Bafica, A.; Santiago, H.C.; Goldszmid, R.; Ropert, C.; Gazzinelli, R.T.; Sher, A. Cutting edge: TLR9 and TLR2 signaling together account for MyD88-dependent control of parasitemia in Trypanosoma cruzi infection. J. Immunol. 2006, 177, 3515–3519. [Google Scholar] [CrossRef]
- Shoda, L.K.; Kegerreis, K.A.; Suarez, C.E.; Roditi, I.; Corral, R.S.; Bertot, G.M.; Norimine, J.; Brown, W.C. DNA from protozoan parasites Babesia bovis, Trypanosoma cruzi, and T. brucei is mitogenic for B lymphocytes and stimulates macrophage expression of interleukin-12, tumor necrosis factor alpha, and nitric oxide. Infect. Immun. 2001, 69, 2162–2171. [Google Scholar] [CrossRef]
- Caetano, B.C.; Carmo, B.B.; Melo, M.B.; Cerny, A.; dos Santos, S.L.; Bartholomeu, D.C.; Golenbock, D.T.; Gazzinelli, R.T. Requirement of UNC93B1 reveals a critical role for TLR7 in host resistance to primary infection with Trypanosoma cruzi. J. Immunol. 2011, 187, 1903–1911. [Google Scholar] [CrossRef]
- Sousa-Rocha, D.; Thomaz-Tobias, M.; Diniz, L.F.A.; Souza, P.S.S.; Pinge-Filho, P.; Toledo, K.A. Trypanosoma cruzi and Its Soluble Antigens Induce NET Release by Stimulating Toll-Like Receptors. PLoS ONE 2015, 10, e0139569. [Google Scholar] [CrossRef]
- Cronemberger-Andrade, A.; Xander, P.; Soares, R.P.; Pessoa, N.L.; Campos, M.A.; Ellis, C.C.; Grajeda, B.; Ofir-Birin, Y.; Almeida, I.C.; Regev-Rudzki, N.; et al. Trypanosoma cruzi-Infected Human Macrophages Shed Proinflammatory Extracellular Vesicles That Enhance Host-Cell Invasion via Toll-Like Receptor 2. Front. Cell. Infect. Microbiol. 2020, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, G.; Salazar-Alcalá, E.; Hernández, F.; Deglesne, P.A.; Bello, Z.D.; de Noya, B.A.; Noya, O.; Fernández-Mestre, M. Polymorphisms of the TLR4 gene: Risk factor for chronicity and severity in oral vectorial Chagas disease. Exp. Parasitol. 2022, 238, 108243. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Lizama, L.E.; Cruz-Chan, J.V.; Versteeg, L.; Teh-Poot, C.F.; Hoffman, K.; Kendricks, A.; Keegan, B.; Pollet, J.; Gusovsky, F.; Hotez, P.J.; et al. TLR4 agonist protects against Trypanosoma cruzi acute lethal infection by decreasing cardiac parasite burdens. Parasite Immunol. 2020, 42, e12769. [Google Scholar] [CrossRef] [PubMed]
- Queiroga, T.B.D.; Pereira, N.S.; da Silva, D.D.; Andrade, C.M.; de Araújo Júnior, R.F.; Brito, C.R.D.N.; Galvão, L.M.D.C.; da Câmara, A.C.J.; Nascimento, M.S.L.; Guedes, P.M.M. Virulence of Trypanosoma cruzi Strains Is Related to the Differential Expression of Innate Immune Receptors in the Heart. Front. Cell. Infect. Microbiol. 2021, 11, 696719. [Google Scholar] [CrossRef] [PubMed]
- Pereira, N.S.; Queiroga, T.B.D.; Nunes, D.F.; Andrade, C.M.; Nascimento, M.S.L.; Do-Valle-Matta, M.A.; da Câmara, A.C.J.; Galvão, L.M.D.C.; Guedes, P.M.M.; Chiari, E. Innate immune receptors over expression correlate with chronic chagasic cardiomyopathy and digestive damage in patients. PLoS Negl. Trop. Dis. 2018, 12, e0006589. [Google Scholar] [CrossRef]
- Castillo, C.; Muñoz, L.; Carrillo, I.; Liempi, A.; Medina, L.; Galanti, N.; Maya, J.D.; Kemmerling, U. Toll-like receptor-2 mediates local innate immune response against Trypanosoma cruzi in ex vivo infected human placental chorionic villi explants. Placenta 2017, 60, 40–46. [Google Scholar] [CrossRef]
- Blanda, V.; Bracale, U.M.; Di Taranto, M.D.; Fortunato, G. Galectin-3 in Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 9232. [Google Scholar] [CrossRef]
- Pineda, M.A.; Cuervo, H.; Fresno, M.; Soto, M.; Bonay, P. Lack of Galectin-3 Prevents Cardiac Fibrosis and Effective Immune Responses in a Murine Model of Trypanosoma cruzi Infection. J. Infect. Dis. 2015, 212, 1160–1171. [Google Scholar] [CrossRef]
- Brown, S.P.; Cornforth, D.M.; Mideo, N. Evolution of virulence in opportunistic pathogens: Generalism, plasticity, and control. Trends. Microbiol. 2012, 20, 336–342. [Google Scholar] [CrossRef]
- Epting, C.L.; Coates, B.M.; Engman, D.M. Molecular mechanisms of host cell invasion by Trypanosoma cruzi. Exp. Parasitol. 2010, 126, 283–291. [Google Scholar] [CrossRef]
- Koo, S.-J.; Szczesny, B.; Wan, X.; Putluri, N.; Garg, N.J. Pentose Phosphate Shunt Modulates Reactive Oxygen Species and Nitric Oxide Production Controlling Trypanosoma cruzi in Macrophages. Front. Immunol. 2018, 9, 202. [Google Scholar] [CrossRef]
- Mesıas, A.C.; Garg, N.J.; Zago, M.P. Redox Balance Keepers and Possible Cell Functions Managed by Redox Homeostasis in Trypanosoma cruzi. Front. Cell. Infect. Microbiol. 2019, 9, 435. [Google Scholar] [CrossRef]
- Piacenza, L.; Peluffo, G.; Alvarez, M.N.; Martınez, A.; Radi, R. Trypanosoma cruzi antioxidant enzymes as virulence factors in chagas disease. Antioxid. Redox Signal. 2013, 19, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Kipnis, T.L.; David, J.R.; Alper, C.A.; Sher, A.; da Silva, W.D. Enzymatic treatment transforms trypomastigotes of Trypanosoma cruzi into activators of alternative complement pathway and potentiates their uptake by macrophages. Proc. Natl. Acad. Sci. USA 1981, 78, 602–605. [Google Scholar] [CrossRef]
- Norris, K.A.; Bradt, B.; Cooper, N.R.; So, M. Characterization of a Trypanosoma cruzi C3 binding protein with functional and genetic similarities to the human complement regulatory protein, decay-accelerating factor. J. Immunol. 1991, 147, 2240–2247. [Google Scholar] [CrossRef]
- Tambourgi, D.V.; Kipnis, T.L.; da Silva, W.D.; Joiner, K.A.; Sher, A.; Heath, S.; Hall, B.F.; Ogden, G.B. A partial cDNA clone of trypomastigote decay-accelerating factor (T-DAF), a developmentally regulated complement inhibitor of Trypanosoma cruzi, has genetic and functional similarities to the human complement inhibitor DAF. Infect. Immun. 1993, 61, 3656–3663. [Google Scholar] [CrossRef]
- Schenkman, S.; Eichinger, D.; Pereira, M.E.A.; Nussenzweig, V. Structural and functional properties of Trypanosoma trans-sialidase. Annu. Rev. Microbiol. 1994, 48, 499–523. [Google Scholar] [CrossRef]
- Valck, C.; Ramirez, G.; Lopez, N.; Ribeiro, C.H.; Maldonado, I.; Sanchez, G.; Ferreira, V.P.; Schwaeble, W.; Ferreira, A. Molecular mechanisms involved in the inactivation of the first component of human complement by Trypanosoma cruzi calreticulin. Mol. Immunol. 2010, 47, 1516–1521. [Google Scholar] [CrossRef]
- Chamond, N.; Gregoire, C.; Coatnoan, N.; Rougeot, C.; Freitas Junior, L.H.; da Silveira, J.F.; Degrave, W.M.; Minoprio, P. Biochemical characterization of proline racemases from the human protozoan parasite Trypanosoma cruzi and definition of putative protein signatures. J. Biol. Chem. 2003, 278, 15484–15494. [Google Scholar] [CrossRef]
- Reina-San-Martin, B.; Degrave, W.; Rougeot, C.; Cosson, A.; Chamond, N.; Cordeiro-Da-Silva, A.; Arala-Chaves, M.; Coutinho, A.; Minoprio, P. A B-cell mitogen from a pathogenic trypanosome is a eukaryotic proline racemase. Nat. Med. 2000, 6, 890–897. [Google Scholar] [CrossRef]
- Chamond, N.; Goytia, M.; Coatnoan, N.; Barale, J.C.; Cosson, A.; Degrave, W.M.; Minoprio, P. Trypanosoma cruzi proline racemases are involved in parasite differentiation and infectivity. Mol. Microbiol. 2005, 58, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Ouaissi, M.A.; Dubremetz, J.F.; Schoneck, R.; Fernandez-Gomez, R.; Gomez-Corvera, R.; Billaut-Mulot, O.; Taibi, A.; Loyens, M.; Tartar, A.; Sergheraert, C.; et al. Trypanosoma cruzi: A 52-kDa protein sharing sequence homology with glutathione S-transferase is localized in parasite organelles morphologically resembling reservosomes. Exp. Parasitol. 1995, 81, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Ouaissi, M.A.; Guilvard, E.; Delneste, Y.; Caron, G.; Magistrelli, G.; Herbault, N.; Thieblemont, N.; Jeannin, P. The Trypanosoma cruzi Tc52- released protein induces human dendritic cell maturation, signals via toll-like receptor 2, and confers protection against lethal infection. J. Immunol. 2002, 168, 6366–6374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfim-Melo, A.; Ferreira, E.R.; Florentino, P.T.V.; Mortara, R.A. Amastigote Synapse: The Tricks of Trypanosoma cruzi Extracellular Amastigotes. Front. Microbiol. 2018, 9, 1341. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.A.; Clemente, T.M.; Dos Santos, M.A.; Machado, F.C.; Gomes, R.G.; Moreira, H.H.; Cruz, M.C.; Brígido, P.C.; Dos Santos, P.C.; Martins, F.A.; et al. A Recombinant Protein Based on Trypanosoma cruzi P21 Enhances Phagocytosis. PLoS ONE 2012, 7, e51384. [Google Scholar] [CrossRef]
- Ferreira, É.R.; Horjales, E.; Bonfim-Melo, A.; Cortez, C.; da Silva, C.V.; De Groote, M.; Sobreira, T.J.P.; Cruz, M.C.; Lima, F.M.; Cordero, E.M.; et al. Unique behavior of Trypanosoma cruzi mevalonate kinase: A conserved glycosomal enzyme involved in host cell invasion and signaling. Sci. Rep. 2016, 6, 24610. [Google Scholar] [CrossRef]
- de Castro Neto, A.L.; da Silveira, J.F.; Mortara, R.A. Comparative Analysis of Virulence Mechanisms of Trypanosomatids Pathogenic to Humans. Front. Cell. Infect. Microbiol. 2021, 16, 669079. [Google Scholar] [CrossRef]
- Freire-De-Lima, L.; Fonseca, L.M.; Oeltmann, T.; Mendoncą-Previato, L.; Previato, J.O. The trans-sialidase, the major Trypanosoma cruzi virulence factor: Three decades of studies. Glycobiology 2015, 25, 1142–1149. [Google Scholar] [CrossRef]
- Campetella, O.; Buscaglia, C.A.; Mucci, J.; Leguizamón, M.S. Parasite-host glycan interactions during Trypanosoma cruzi infection: Trans-Sialidase rides the show. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2020, 1866, 165692. [Google Scholar] [CrossRef]
- Magdesian, M.H.; Giordano, R.; Ulrich, H.; Juliano, M.A.; Juliano, L.; Schumacher, R.I.; Colli, W.; Alves, M.J.M. Infection by Trypanosoma cruzi. J. Biol. Chem. 2001, 276, 19382–19389. [Google Scholar] [CrossRef]
- Acosta-Serrano, A.; Almeida, I.C.; Freitas-Junior, L.H.; Yoshida, N.; Schenkman, S. The mucin-like glycoprotein super-family of Trypanosoma cruzi: Structure and biological roles. Mol. Biochem. Parasitol. 2001, 114, 143–150. [Google Scholar] [CrossRef]
- Herreros-Cabello, A.; Callejas-Hernández, F.; Gironès, N.; Fresno, M. Trypanosoma cruzi Genome: Organization, Multi-Gene Families, Transcription, and Biological Implications. Genes 2020, 11, 1196. [Google Scholar] [CrossRef]
- dos Santos, S.L.; Freitas, L.M.; Lobo, F.P.; Rodrigues-Luiz, G.F.; Mendes, T.A.; Oliveira, A.C.; Andrade, L.O.; Chiari, E.; Gazzinelli, R.T.; Teixeira, S.M.; et al. The MASP Family of Trypanosoma cruzi: Changes in Gene Expression and Antigenic Profile during the Acute Phase of Experimental Infection. PloS Negl. Trop. Dis. 2012, 6, e1779. [Google Scholar] [CrossRef]
- Bartholomeu, D.C.; Cerqueira, G.C.; Leão, A.C.A.; daRocha, W.D.; Pais, F.S.; Macedo, C.; Djikeng, A.; Teixeira, S.M.; El-Sayed, N.M. Genomic organization and expression profile of the mucin-associated surface protein (masp) family of the human pathogen Trypanosoma cruzi. Nucleic Acids Res. 2009, 37, 3407–3417. [Google Scholar] [CrossRef]
- Maeda, F.Y.; Cortez, C.; Yoshida, N. Cell signaling during Trypanosoma cruzi invasion. Front. Immunol. 2012, 3, 361. [Google Scholar] [CrossRef]
- Osorio, L.; Rıos, I.; Gutierrez, B.; González, J. Virulence factors of Trypanosoma cruzi: Who is who? Microbes Infect. 2012, 14, 1390–1402. [Google Scholar] [CrossRef]
- Alvarez, V.E.; Niemirowicz, G.T.; Cazzulo, J.J. The peptidases of Trypanosoma cruzi: Digestive enzymes, virulence factors, and mediators of autophagy and programmed cell death. Biochim. Biophys. Acta-Proteins Proteom. 2012, 1824, 195–206. [Google Scholar] [CrossRef]
- Garcia, E.S.; Ratcliffe, N.A.; Whitten, M.M.; Gonzalez, M.S.; Azambuja, P. Exploring the role of insect host factors in the dynamics of Trypanosoma cruzi-Rhodnius prolixus interactions. J. Insect. Physiol. 2007, 53, 11–21. [Google Scholar] [CrossRef]
- Tardieux, I.; Nathanson, M.H.; Andrews, N.W. Role in host cell invasion of Trypanosoma cruzi-induced cytosolic-free Ca2+ transients. J. Exp. Med. 1994, 179, 1017–1022. [Google Scholar] [CrossRef]
- Rodríguez, A.; Samoff, E.; Rioult, M.G.; Chung, A.; Andrews, N.W. Host cell invasion by trypanosomes requires lysosomes and microtubule/kinesin-mediated transport. J. Cell Biol. 1996, 134, 349–362. [Google Scholar] [CrossRef]
- Hissa, B.; de Oliveira Andrade, L. Trypasonoma cruzi uses a specific subset of host cell lysosomes for cell invasion. Parasitol. Int. 2015, 64, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Cueto, J.A.; Vanrell, M.C.; Salassa, B.N.; Nola, S.; Galli, T.; Colombo, M.I.; Romano, P.S. Soluble N-ethylmaleimide-sensitive factor attachment protein receptors required during Trypanosoma cruzi parasitophorous vacuole development. Cell. Microbiol. 2017, 19, e12713. [Google Scholar] [CrossRef] [PubMed]
- Tardieux, I.; Webster, P.; Ravesloot, J.; Boron, W.; Lunn, J.A.; Heuser, J.E.; Andrews, N.W. Lysosome recruitment and fusion are early events required for trypanosome invasion of mammalian cells. Cell 1992, 71, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Woolsey, A.M.; Sunwoo, L.; Petersen, C.A.; Brachmann, S.M.; Cantley, L.C.; Burleigh, B.A. Novel PI 3-kinase-dependent mechanisms of trypanosome invasion and vacuole maturation. J. Cell Sci. 2003, 116, 3611–3622. [Google Scholar] [CrossRef]
- Andrade, L.O.; Andrews, N.W. Lysosomal fusion is essential for the retention of Trypanosoma cruzi inside host cells. J. Exp. Med. 2004, 200, 1135–1143. [Google Scholar] [CrossRef]
- Andrade, L.O.; Andrews, N.W. The Trypanosoma cruzi—Host-cell interplay: Location, invasion, retention. Nat. Rev. Microbiol. 2005, 3, 819–823. [Google Scholar] [CrossRef]
- Romano, P.S.; Arboit, M.A.; Vázquez, C.L.; Colombo, M.I. The autophagic pathway is a key component in the lysosomal dependent entry of Trypanosoma cruzi into the host cell. Autophagy 2009, 5, 6–18. [Google Scholar] [CrossRef]
- Veiga-Santos, P.; Desoti, V.C.; Miranda, N.; Ueda-Nakamura, T.; Dias-Filho, B.P.; Silva, S.O.; Cortez, D.A.; de Mello, J.C.; Nakamura, C.V. The natural compounds piperovatine and piperlonguminine induce autophagic cell death on Trypanosoma cruzi. Acta Trop. 2013, 125, 349–356. [Google Scholar] [CrossRef]
- Lazarin-Bidóia, D.; Desoti, V.C.; Martins, S.C.; Ribeiro, F.M.; Ud Din, Z.M.; Rodrigues-Filho, E.; Ueda-Nakamura, T.; Nakamura, C.V.; de Oliveira Silva, S. Dibenzylideneacetones are potent trypanocidal compounds that affect the Trypanosoma cruzi redox system. Antimicrob. Agents Chemother. 2016, 60, 890–903. [Google Scholar] [CrossRef]
- Duque, T.L.A.; Siqueira, M.S.; Travassos, L.H.; Moreira, O.C.; Bozza, P.T.; Melo, R.C.N.; Henriques-Pons, A.; Menna-Barreto, R.F.S. The induction of host cell autophagy triggers defense mechanisms against Trypanosoma cruzi infection in vitro. Eur. J. Cell Biol. 2020, 99, 51060. [Google Scholar] [CrossRef]
- Losinno, A.D.; Martínez, S.J.; Labriola, C.A.; Carrillo, C.; Romano, P.S. Induction of autophagy increases the proteolytic activity of reservosomes during Trypanosoma cruzi metacyclogenesis. Autophagy 2021, 17, 439–456. [Google Scholar] [CrossRef]
- Castro, J.A.; de Mecca, M.M.; Bartel, L.C. Toxic Side Effects of Drugs Used to Treat Chagas’ Disease (American Trypanosomiasis). Hum. Exp. Toxicol. 2006, 25, 471–479. [Google Scholar] [CrossRef]
- Bruno, F.; Castelli, G.; Vitale, F.; Catanzaro, S.; Badaco, V.V.; Roberti, M.; Colomba, C.; Cascio, A.; Tolomeo, M. Antiparasitic Effect of Stilbene and Terphenyl Compounds against Trypanosoma cruzi Parasites. Pharmaceuticals 2021, 14, 1199. [Google Scholar] [CrossRef]
- Kratz, J.M. Drug discovery for chagas disease: A viewpoint. Acta Trop. 2019, 198, 105107. [Google Scholar] [CrossRef]
- Kratz, J.M.; Garcia Bournissen, F.; Forsyth, C.J.; Sosa-Estani, S. Clinical and pharmacological profile of benznidazole for treatment of Chagas disease. Expert Rev. Clin. Pharmacol. 2018, 11, 943–957. [Google Scholar] [CrossRef]
- Molina, I.; Salvador, F.; Sánchez-Montalvá, A.; Treviño, B.; Serre, N.; Avilés, A.S.; Almirante, B. Toxic profile of benznidazole in patients with chronic chagas disease: Risk factors and comparison of the product from two different manufacturers. Antimicrob. Agents Chemother. 2015, 59, 6125–6131. [Google Scholar] [CrossRef]
- Morillo, C.A.; Marin-Neto, J.A.; Avezum, A.; Sosa-Estani, S.; Rassi, A.; Rosas, F.; Villena, E.; Quiroz, R.; Bonilla, R.; Britto, C.; et al. BENEFIT Investigators. Randomized trial of benznidazole for chronic chagas’ cardiomyopathy. N. Engl. J. Med. 2015, 373, 1295–1306. [Google Scholar] [CrossRef]
- Ribeiro, I.; Sevcsik, A.M.; Alves, F.; Diap, G.; Don, R.; Harhay, M.O.; Chang, S.; Pecoul, B. New, improved treatments for Chagas disease: From the R&D pipeline to the patients. PLoS Negl. Trop. Dis. 2009, 3, e484. [Google Scholar]
- Chatelain, E. Chagas disease research and development: Is there light at the end of the tunnel? Comput. Struct. Biotechnol. J. 2017, 15, 98–103. [Google Scholar] [CrossRef]
- Rao, S.P.S.; Barrett, M.P.; Dranoff, G.; Faraday, C.J.; Gimpelewicz, C.R.; Hailu, A.; Jones, C.L.; Kelly, J.M.; Lazdins-Helds, J.K.; Mäser, P.; et al. Drug discovery for kinetoplastid diseases: Future directions. ACS Infect. 2019, 5, 152–157. [Google Scholar] [CrossRef]
- Rios, L.E.; Vázquez-Chagoyán, J.C.; Pacheco, A.O.; Zago, M.P.; Garg, N.J. Immunity and vaccine development efforts against Trypanosoma cruzi. Acta Trop. 2019, 200, 105168. [Google Scholar] [CrossRef] [PubMed]
- Camargo, E.P.; Gazzinelli, R.T.; Morel, C.M.; Precioso, A.R. Why do we still have not a vaccine against Chagas disease? Memórias Inst. Oswaldo Cruz 2022, 117, e200314. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Chagoyán, J.C.; Gupta, S.; Garg, N.J. Vaccine development against Trypanosoma cruzi and Chagas disease. Adv Parasitol. 2011, 75, 121–146. [Google Scholar] [PubMed]
- Schmunis, G.A.; Yadon, Z.E. Chagas disease: A Latin American health problem becoming a world health problem. Acta Trop. 2010, 115, 14–21. [Google Scholar] [CrossRef]
- Brisseau, J.M.; Cebron, J.P.; Petit, T.; Marjolet, M.; Cuilliere, P.; Godin, J.; Grolleau, J.Y. Chagas & myocarditis imported into France. Lancet 1988, 7, 1046. [Google Scholar]
- Crovato, F.; Rebora, A. Chagas & disease: A potential plague for Europe. Dermatology 1997, 195, 184–185. [Google Scholar]
- Angheben, A.; Anselmi, M.; Gobbi, F.; Marocco, S.; Monteiro, G.; Buonfrate, D.; Tais, S.; Talamo, M.; Zavarise, G.; Strohmeyer, M.; et al. Chagas disease in Italy: Breaking an epidemiological silence. Eurosurveillance 2011, 16, 19969. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macaluso, G.; Grippi, F.; Di Bella, S.; Blanda, V.; Gucciardi, F.; Torina, A.; Guercio, A.; Cannella, V. A Review on the Immunological Response against Trypanosoma cruzi. Pathogens 2023, 12, 282. https://doi.org/10.3390/pathogens12020282
Macaluso G, Grippi F, Di Bella S, Blanda V, Gucciardi F, Torina A, Guercio A, Cannella V. A Review on the Immunological Response against Trypanosoma cruzi. Pathogens. 2023; 12(2):282. https://doi.org/10.3390/pathogens12020282
Chicago/Turabian StyleMacaluso, Giusi, Francesca Grippi, Santina Di Bella, Valeria Blanda, Francesca Gucciardi, Alessandra Torina, Annalisa Guercio, and Vincenza Cannella. 2023. "A Review on the Immunological Response against Trypanosoma cruzi" Pathogens 12, no. 2: 282. https://doi.org/10.3390/pathogens12020282