
Exploring Possible Ways to Enhance the Potential and Use of Natural Products through Nanotechnology in the Battle against Biofilms of Foodborne Bacterial Pathogens

 , , and
, , and
Abstract
:1. Introduction
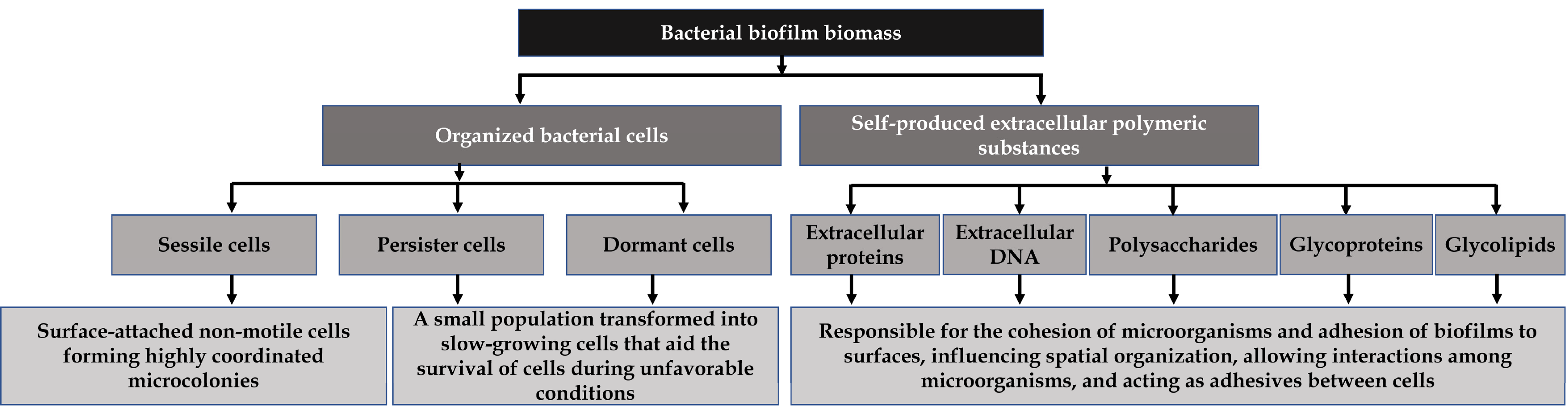
2. Significance of Pathogenic Bacteria and Their Biofilms in the Food Industry
3. Natural Products as a Promising Alternate
3.1. Natural Bioactives as Preservatives

{kind=link}
| Natural Bioactive and Source | Target Foodborne Pathogen(s) | Mechanism of Action | Reference(s) |
|---|---|---|---|
| Plant source | |||
| Essential oil of Citrus reticulata | E. coli and S. aureus | C. reticulata essential oil inhibited biofilm formation and eradicated the mature biofilm | [64] |
| Essential oils components from Allium sativum and Cuminum cyminum | S. Typhimurium | Essential oils eliminated the planktonic and biofilm forms of S. Typhimurium by downregulating QS (sdiA and luxS) and cellulose synthesis (csgD and adrA) genes | [65] |
| Essential oil of Thymus daenensis and Satureja hortensis | E. coli O157:H7 | Essential oils reduced biofilm formation by downregulating the expression of luxS and pfs genes | [66] |
| Essential oil of Artemisia dracunculus | S. Typhimurium and S. aureus | Essential oil inhibited biofilm formation as well as disrupted existing biofilms Downregulation of luxS and pfs in S. Typhimurium and hld in S. aureus are expected to be the mechanism of action | [67] |
| Lemongrass essential oil | L. monocytogenes and S. aureus | In L. monocytogenes, an active compound penetrated the biofilm and inhibited its formation In S. aureus, biofilm disruption was observed | [68] |
| Menthol and menthone from peppermint essential oil (PEO) of Mentha × piperita | S. aureus | Active against biofilm formation in S. aureus Moreover, the essential oil effectively killed and eradicated the residents in preformed biofilms | [69] |
| Thymol from the essential oil of Thymus zygis | L. monocytogenes | Thymol reduced eDNA release and the expression of enzymes related to biofilm formation In addition, thymol downregulated the expression of biofilm and polysaccharide adhesion-related genes | [70] |
| Limonene, pinene, terpinene, and myrcene from the essential oil of Citrus medica | L. monocytogenes | Essential oil components targeted biofilm formation and eradicated the preformed biofilms of L. monocytogenes | [71] |
| Tea catechin extract | MRSA | Tea catechin extract inhibited biofilm formation by downregulating biofilm-related genes such as fnbA and icaBC | [72] |
| Exocarp extract of Gingko biloba | MRSA and S. aureus | The extract inhibited biofilm formation and preformed biofilms of S. aureus Gene expression of icaA and sarA were lowered after 6 h and sigB after 12 h, while upregulation of icaR was observed after 12 h | [73] |
| Peel extract of Citrus sinensis | P. aeruginosa and S. aureus | The peel extract of C. sinensis prevented MRSA and ESBL development in the biofilm matrix | [74] |
| Flesh extract of Moringa oleifera | P. aeruginosa and S. aureus | M. oleifera prevented the development of MRSA biofilm matrix M. oleifera inhibited the development of single as well as mixed biofilm cultures better than C. sinensis | [74] |
| Root extract of Veteriveria zizanioides | S. aureus | The methanolic extract prevented the biofilm formation of S. aureus Targeted the genes involved in the initial attachment like clfA and fnbAB | [75] |
| Barb extract of Butia odorata | S. aureus | The barb extract reduced the number of biofilm cells | [76] |
| Ohelo berry extract | L. monocytogenes | The extract targeted biofilm formation via downregulation of the global regulator gene sigB | [77] |
| Spice extract from Syzygium aromaticum and Cinnamomum verum | L. monocytogenes | The extract targeted biofilm formation by modulating the release of eDNA In addition, it downregulated the genes responsible for biofilm formation such as lapB, actA, flaA, prfA, and inlA | [78] |
| Methanol extract of Lonicera caerulea var. emphyllocalyx | Enteropathogenic E. coli | The methanolic extract targeted biofilm formation and reduced the spread of biofilm Biofilm-related genes, such as fliC, csgA, and fimA, were downregulated upon treatment with the extract | [79] |
| Emodin, chrysophanol, and physcion from Rumex japonicus extract | S. aureus | Inhibition of biofilm formation and removal of biofilm | [80] |
| Baicalein from Scutellaria baicalensis extract | S. aureus | Inhibition and disruption of biofilm formation Downregulated staphylococcal enterotoxin A (sea) and α-hemolysin (hla) levels Downregulated QS-related genes agrA, RNAIII, and sarA as well as icaA | [52] |
| 2-hydroxy-4-methoxybenzaldehyde from Hemidesmus indicus | MRSA | Inhibition and disruption of biofilms Downregulated virulence factors such as hemolysin, nuclease, lipase and staphyloxanthin Downregulated the global regulator, SigB, required for biofilm formation | [43,81] |
| Kaempferol from Glycine max | S. aureus | Inhibited biofilm formation by reducing the expression of sortase (srtA) and adhesion-related genes | [82] |
| Quinic acid | S. aureus | Reduction in biofilm biomass Downregulation of sarA and upregulation of agrA Reduction in the number of sedentary cells on stainless steel | [83] |
| Naringenin | S. aureus | Inhibited biofilm formation by downregulating the expression of sigB, icaA, agrA, and sarA. | [84] |
| Papain from Carica papaya | S. aureus and C. jejuni | Inhibited biofilm formation Degraded the mature biofilm formed on stainless steel | [85] |
| Phloretin | L. monocytogenes | Phloretin targeted biofilm formation by repressing quorum-sensing gene expression | [86] |
| Pristimerin | S. aureus | Inhibition, disruption, and dispersal of biofilm | [87] |
| Quercetin | S. Typhimurium and S. enteritidis | Inhibited biofilm formation by downregulating the expression of genes encoding virulence factor (avrA, and hilA), stress response (rpoS), and quorum-sensing (luxS) | [88] |
| Rutin | E. coli and S. aureus | Inhibition and reduction of biofilm formation Reduction of mono- and multi-species biofilms in a concentration-dependent fashion | [89] |
| (−)-tetrahydroberberrubine∙acetate (THBA) from Nandina domestica | B. cereus | Inhibition of biofilm formation and disruption of mature biofilms | [90] |
| Animal source | |||
| BCp12 peptide derived from Milk | S. aureus | Inhibited biofilm formation Downregulated the expression of genes related to the QS system, including agrA, agrB, agrC, and psmB. | [91] |
| β-GBP peptide sequence from the Penaeus vannamei | B. subtilis | Reduced preformed biofilms | [92] |
| Hc-CATH peptide from sea snake Hydrophis cyanocinctus and As-CATH4 and As-CATH5 from Alligator sinensis | A. junii and P. mirabilis | Inhibition, as well as the removal of biofilms | [93] |
| Manuka Honey | E. coli O157:H7 | Inhibition, as well as the removal of biofilms | [94] |
| Apitoxin from Apis mellifera | S. enterica | Reduced biofilm formation Expression of biofilm and virulence-related genes was different in different strains | [36] |
| Avarol from Dysidea avara | P. aeruginosa PAO1 | Reduced biofilm formation | [95] |
| Bacterial source | |||
| Peptide AL705 | L. monocytogenes FBUNT | Reduced biofilm formation by disturbing QS signaling | [96] |
| Sonorensin from Bacillus sonorensis MT93 | S. aureus | Inhibited biofilm formation by increasing membrane permeability | [97] |
| Cell-free supernatant of Lactobacillus curvatus ET31 | L. monocytogenes | Inhibition of biofilm formation by targeting luxS gene expression However, the cell-free supernatant was not able to remove the preformed biofilm | [38] |
| Bacteriophage | |||
| JK004 | Cronobacter sakazakii | Removed the biofilm | [98] |
| vB_STM-2 | S. Typhimurium (ST-4) | Removed the biofilm Penetrated into EPS and depolymerized EPS | [39] |
3.2. Challenges in the Use of Natural Preservatives for Preservation
4. Nanotechnology Approach: An Emerging Antibiofilm Platform
4.1. Techniques for Targeting Biofilms through Natural Product Delivery
4.1.1. Biofabrication of Nanoparticles with Antibiofilm Activity Using Surface Functionalization
| Type of Nanoformulation | Active Agents | Properties of Nanoparticles | Biological Activity against Foodborne Pathogens | Reference |
|---|---|---|---|---|
| Silver nanoparticles | Terminalia catappa | ~26–100 nm in size | At sub-MIC concentrations, silver nanoparticles dislodged the preformed biofilms of L. monocytogenes and entrenched the biofilm cells. The prepared silver nanoparticles enhanced the life span of infected C. elegans by 90%. | [119] |
| Copper oxide nanoparticles | Leaf extract of Eucalyptus globulus | ~16.78 nm in size | The tested concentrations inhibited biofilm formation in the range of 44.41 ± 7 to 70.75 ± 8% and 34.41 ± 7 to 62.29 ± 8% in E. coli and MRSA, respectively. | [123] |
| Gold nanoparticles | Caffeine | 77.0 ± 5.0 nm in size | At 256 µg/mL, caffeine-loaded gold nanoparticles reduced biofilm formation by 69.55, 63.74, and 64.89% in S. aureus, L. monocytogenes, and E. coli, respectively. At MIC (512 µg/mL), the nanoparticles eradicated preformed biofilms by 63.71 and 77.4% in E. coli and S. aureus, respectively. It was 79.66% effective against L. monocytogenes with an MIC value of 1024 µg/mL. Further, the caffeine-loaded nanoparticles were potent enough to eradicate the biofilm-embedded persister cells of the tested pathogens. | [124] |
| Gold nanoparticles | Seed Extract of Trachyspermum ammi | ~24.4 nm in size Shapes appeared to be spherical, spheroid, and a few anisotropies | At 0.5 MIC, the gold nanoparticles reduced biofilm formation by 81 and 73% in Serratia marcescens and L. monocytogenes, respectively. Gold nanoparticles targeted biofilm formation of the test pathogens by reducing the biofilm-related virulence factors such as cell surface hydrophobicity, motility, and exopolysaccharides. Increased ROS production was expected to play a role in eradicating the preformed biofilms of the tested pathogens. | [125] |
| Copper oxide nanoparticles | Leaf extract of Mentha spicata | Spherical nanoparticles with 36 nm in size | The synthesized copper oxide nanoparticles were superior in inhibiting the biofilm formation of S. aureus (98%) compared to E. coli (86%) at 100 µg/mL. | [126] |
| Copper nanoparticles | Cassinopin and isoquercetin from the leaves of Crotalaria candicans | Spherical-shaped cassinopin- and isoquercetin-capped nanoparticles were 66 and 69 nm in size, respectively | The synthesized nanoparticles were proficient enough to thwart the biofilm formation of MRSA, S. Typhi, and E. coli, even at a concentration of 2 µg/mL. | [122] |
| Silver nanoparticles | Kaempferitrin from Crotalaria juncea | Spherical-shaped nanoparticles with an average size of 33 nm and zeta potential of −33.8 mV | Kaempferitrin-loaded silver nanoparticles were able to retard biofilm formation in MRSA. Enhancing the cell membrane permeability in the cells treated with bioactive-loaded silver nanoparticles was suggested to be the antibiofilm mechanism. Further, the silver nanoparticles reduced the bacterial burden by ~1.8-fold in the infected zebrafish with no toxicity. | [127] |
| Copper nanoparticles | Kaempferitrin from Crotalaria juncea | Spherical-shaped nanoparticles with an average size of 56 nm and a zeta potential of −38 mV | Kaempferitrin-loaded copper nanoparticles were able to retard the biofilm formation in MRSA. Enhancing the cell membrane permeability in the cells treated with bioactive-loaded copper nanoparticles is expected to be the possible antibiofilm mechanism. Further, the silver nanoparticles reduced the bacterial burden by ~2-folds in the infected Zebrafish, leaving no toxicity | [127] |
| Gold nanoparticles | Aerial part extract of Origanum majoranum | Spherical-shaped nanoparticles with sizes ranging between 5 and 30 nm | The gold nanoparticles were prepared using L-glutathione. The prepared gold nanoparticles were then conjugated with O. majoranum extract. The gold nanoparticles showed antibacterial, antibiofilm, and antioxidant activities. Compared to the extract, the extract-conjugated gold nanoparticles showed antibiofilm activities against S. aureus and E. coli with inhibition rates of 19.2 and 30%, respectively. | [128] |
| Zinc oxide nanoparticles | Fruit extract of Aegle marmelos | Hexagonal wurtzite-shaped nanoparticle with an average size of 22.5 nm | The zinc oxide nanoparticles coated with A. marmelos efficiently inhibited the biofilm formation of S. aureus and were validated with confocal microscopic analysis. Further, the use of nanoparticles reduced the pathogen’s hydrophobicity index and enhanced the pathogen’s susceptibility to hydrogen peroxide. | [129] |
| Silver nanoparticles | Oscillatoria sp. | Spherical-shaped nanoparticles with an average size of 10 nm and polydispersity index of 0.58 | The synthesized nanoparticles showed potent biofilm inhibition in S. aureus, S. Typhi, and E. coli. The synthesized silver nanoparticles showed reduced toxicity towards the shrimp model Artemia salina with an LC50 of 2060 µg/mL. | [130] |
| Zinc oxide nanoparticles | Leaf extract of Acacia arabica | Rod-shaped nanoparticles with an average size of 11.3 nm | The zinc oxide nanoparticles at 0.5 MIC reduced the biofilm formation of E. coli, S. aureus, and S. enterica. Further, the EPS estimation assay revealed that these nanoparticles targeted EPS production to hinder biofilm production in the tested pathogens. The nanoparticles showed no toxicity towards HeLa cells at the tested concentrations. | [131] |
| Silver nanoparticles | The seed extract of Nigella sativa | Spherical-shaped nanoparticles with an average size of 50 nm | The extract-conjugated nanoparticles showed antibiofilm activity against S. aureus and E. coli with inhibitory rates of 84.92 and 82.84%, respectively. | [132] |
| Iron oxide nanoparticles | Eucalyptus globulus essential oil | Quasi-spherical shaped nanoparticle with an average size of 7.5 ± 2.5 nm | Interfered with the development of biofilm formation in E. coli and S. aureus. The iron oxide nanoparticles were highly biocompatible for the growth and proliferation of amniotic fluid-derived mesenchymal stem cells. | [133] |
4.1.2. Nanoencapsulation of Natural Compounds with Antibiofilm Activity
Polymeric Nanoparticles
Lipid Nanoparticles
Silica Nanoparticles
| Polymers Used for Nanoformulation | Natural Substance | Properties of Nanoparticles | Biological Activity against Foodborne Pathogens | References |
|---|---|---|---|---|
| Nanoparticles—biguanide-based polymetformin from Poly(ethylenimine) | Pluronic F-127 surfactant and tannic acid | Spherical-shaped nanoparticles with an average size of 96 nm and average zeta potential of -43 mV. | The biofilm-eradicating activity of nanoparticles plateaued from 8 µg/mL, which showed a 2.4 log10-fold biofilm reduction in MRSA. Along the same lines, the activity of biguanide-based polymetformin alone plateaued from 32 µg/mL with a 1.5 log10 reduction, and 256 µg/mL for vancomycin with a 1.1 log10 reduction. The nanoparticles showed good compatibility with human red blood cells, mouse embryonic fibroblast 3T3 cells, and human dermal fibroblast cells. | [180] |
| Nanoemulsion—Poly(oxanorbornenimide) modified with guanidinium, maleimide, and tetraethylene glycol monomethyl ether moieties | Eugenol, methyl eugenol, carvacrol, linalool, (+)-limonene, p-cymene, and α-pinene | The average size of the nanoemulsion containing different natural products ranged from ~180 to ~530 nm. | After 3 h of treatment, nanoemulsions with eugenol and carvacrol eradicated 90% of S. aureus biofilm. Compared to a carvacrol-loaded nanoemulsion, eugenol showed low toxicity to 3T3 fibroblast cells. | [181] |
| Polymeric Eudragit® nanocapsules | Carvacrol | Nanocapsules with an average size of 156 nm, PDI value of 0.22, and zeta potential +44.8. | The nanocapsules were active in eradicating the biofilm formation of foodborne pathogens such as Salmonella spp. and E. coli. | [182] |
| Nanoliposomes (L80-T) and nanoarchaeosomes (A80-T) | Thymus vulgaris essential oil | Spherical vesicles. L80-T with Z potential −4.1 ± 0.6 mV, size ∼115 nm, and A80-T with Z potential −6.6 ± 1.5 mV and size ∼130 nm. | Compared to L80-T, A80-T was active against the preformed biofilms of S. aureus and its clinical isolates. Similar to the positive control vancomycin which has no effect even at a concentration 4 to 8-fold higher than the MIC90. However, A80-T at a concentration of MIC90 or MBC dislodged the preformed biofilms. | [183] |
| Liposomes | Berberine and curcumin | Spherical-shaped liposomes with an average size of 253 and a surface charge of −57 mV. | Compared to the free drug, co-encapsulation of BBR and CCR in liposomes decreased their MICs by 87% and 96%, respectively. At 10 µg/mL, dual drug-loaded liposomes disrupted the preformed biofilms of S. aureus. In addition, the dual drug-loaded liposomes reduced the bacterial burden and thereby the infection in L929 fibroblast cells. | [184] |
| Liposomes | Antilisterial peptide (Lys-Val-Asp-His-Phe-Pro-Leu) originated from rice bran protein | Spherical-shaped liposome with an average size of 137.9 ± 3.1 nm and a surface charge of 28.9 ± 0.8mV. | The MIC of liposomes with antilisterial peptide (84.26 µg/mL) against the sessile L. monocytogenes cells was higher than the free peptide (8 µg/mL). However, the liposome encapsulating peptide showed a rapid release and killing of L. monocytogenes cells after 10 min of contact time, which was 120 minutes for free peptide. The liposome-encapsulated drug showed flavor stability for four weeks when stored at 4 °C. | [185] |
| Sol-gel–derived from tetramethylorthosilicate | Garlic extract | -- | Garlic extract-loaded nanoparticles were efficient in penetrating and disrupting the well-established MRSA biofilms. At 5mg/mL, the extract-loaded nanoparticles reduced the biofilm-residing bacterial viability to 80% and reduced the bacterial biofilm thickness to 2.3 µm from 9.6 µm (control). | [186] |
| Solid lipid nanoparticles–Chinese white wax | Curcumin | Spherical-shaped nanoparticles with an average particle size of 402 nm. | The curcumin-loaded nanoparticles showed a sustained drug release profile and showed a higher drug release rate under an acidic (pH 4.5) rather than a neutral (pH 7.4) environment. The curcumin-loaded nanoparticles inhibited the biofilm formation of S. aureus at a concentration of 125 µg/mL. | [187] |
| Solid lipid nanoparticles | Cinnamon oil | Spherical-shaped nanoparticles with an average size of 337.6 nm and zeta potential of −26.6 mV. | Encapsulated solid lipid nanoparticles reduced the antimicrobial activity against E. coli by 2-fold compared to the cinnamon oil alone. Further, the sub-MIC of solid lipid nanoparticles reduced biofilm formation by 55.25%. | [188] |
| Starch nanoparticles | Triphala formulation containing the blended extract of Terminalia chebula, Terminalia bellirica, and Emblica officinalis | The average size of the nanoparticle was 283 nm with a zeta potential of −12 mV. | The starch nanoparticle-encapsulated drug showed a dose-dependent antibiofilm activity against MRSA, with 10 µg/mL being the most effective biofilm inhibitory concentration. | [189] |
| Solid lipid nanoparticles - Glyceryl monostearate and Poloxamer 188 | Anacardic acid | The average size of the nanoparticle was 212 nm with zeta potential and PDI value of−13 mV and 0.285, respectively. | Treatment with anacardic acid-loaded nanoparticles reduced the biofilm formation of S. aureus at 0.097 µg/mL. The nanoparticle treatment reduced the biofilm thickness (50.3 µm, in control) and biofilm biomass (27.77 μm3/μm2, in control) to 18.61 µm and 10.54 μm3/μm2, respectively. | [190] |
| Chitosan nanoparticles | Chrysin | Spherical-shaped nanoparticles with an average size of 355 nm and with PDI value of 0.487. | Chrysin-loaded nanoparticles inhibited the biofilm formation of S. aureus to 54% at 768 µg/mL. The nanoparticles also reduced biofilm-related phenotypes, such as EPS and cell surface hydrophobicity. | [145] |
| Chitosan nanoemulsion | Thymol/thymol essential oil | The mean particle size of thymol and thymol essential oil were 123 and 139 nm, respectively. | Compared to the thymol essential oil, thymol in a nanoemulsion showed better biofilm inhibition against S. aureus (83.78%) and E. coli (83.64%). The thymol nanoemulsion enhanced the shelf life by reducing the total viable count in the pork sample. | [147] |
| Chitosan nanoparticles | Citrus reticulata essential oil | The average particle size ranges between 131 and 162 nm. | The activity of encapsulated nanoparticles was based on the concentration of C. reticulata essential oil. Chitosan nanoparticles loaded with equal volumes of C. reticulata essential oil showed good biofilm inhibition and eradication activity against S. aureus and E. coli. | [148] |
| Chitosan nanoparticles | Cinnamaldehyde | The average size was 298.1 nm with a zeta potential of +38.73 mV. | Cinnamaldehyde-loaded chitosan nanoparticles at 1.25 mg/mL showed antibiofilm activity. Further, these nanoparticles eradicate the preformed biofilm of S. aureus to 48.1%. | [146] |
| Chitosan | Prangos acaulis | Semi-spherical nanoparticles with an average size of 89.8 nm and a zeta potential of 10.78 mV. | The chitosan nanoparticles were effective against the Gram-positive bacteria S. aureus and B. cereus with a biofilm inhibitory rate of nearly 50% compared to the Gram-negative bacteria E. coli with a biofilm inhibitory rate of 32% at 2000 µg/mL. | [191] |
| PLGA | Xylitol | Nanoparticles with an average size of 106–140 nm and a zeta potential of 12.29–34.05 mV. | PLGA nanoparticles loaded with 5% xylitol showed a profound preformed biofilm-eradicating and killing activity against S. aureus. The biofilm penetrating potential of PLGA nanoparticles enhanced the biofilm-eradicating ability of xylitol. | [163] |
| Nanoemulsions | Cinnamon and clove essential oils | The average size of the nanoemulsion was less than 15 nm. The PDI and zeta potential values were below 0.37 and −10.5 mV, respectively. | Compared to Brij-35, the nanoemulsion formed using Tween-20 showed an enhanced rate of biofilm inhibition of 76% in S. aureus. | [192] |
| Nanoemulsions | Eugenol and methyl salicylate | The mean diameter of eugenol and methyl salicylate was 9.389 ± 0.2 nm and 10.81 ± 0.4 nm, with PDI values of 0.345 and 0.371, respectively. | The nanoemulsion carrying the drugs showed higher antibiofilm and antivirulence activity against E. coli O157:H7. The expression of genes related to the biofilm formation in E. coli was significantly downregulated upon treatment with the drug-loaded nanoemulsion. | [174] |
| Nanostructured lipid carriers | Olibanum oil | The resultant nanoparticles had a particle size of ~200 nm, a polydispersity index of ~0.15, and a zeta potential of ~−35mV. | The nanostructured lipid carriers encapsulating olibanum oil effectively killed the biofilm resident C. jejuni cells and had an MIC value of 780 µg/mL, which was very significant compared to the free oil with an MIC of 1–3 mg/mL. | [168] |
5. Strategies to Enhance Biofilm Clearance
5.1. Size and Surface Modifications
5.2. Stimuli-Responsive Release
5.3. Combined Strategies
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef]
- Arunachalam, K.; Davoodbasha, M. Imaging Bacteria and Biofilm by Field Emission Scanning Electron Microscopy. In Analytical Methodologies for Biofilm Research; Springer Protocols Handbooks; Springer: New York, NY, USA, 2021; pp. 205–222. [Google Scholar] [CrossRef]
- van Wolferen, M.; Orell, A.; Albers, S.-V. Archaeal biofilm formation. Nat. Rev. Microbiol. 2018, 16, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Cámara, M.; Green, W.; MacPhee, C.E.; Rakowska, P.D.; Raval, R.; Richardson, M.C.; Slater-Jefferies, J.; Steventon, K.; Webb, J.S. Microbiomes. Economic significance of biofilms: A multidisciplinary and cross-sectoral challenge. Npj Biofilms Microbiomes 2022, 8, 42. [Google Scholar] [CrossRef]
- Alvarez-Ordóñez, A.; Coughlan, L.M.; Briandet, R.; Cotter, P.D. Biofilms in food processing environments: Challenges and opportunities. Annu. Rev. Food Sci. Technol. 2019, 10, 173–195. [Google Scholar] [CrossRef] [PubMed]
- Galie, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Campos, D.; Rodríguez-Melcón, C.; Alonso-Calleja, C.; Capita, R. Persistent Listeria monocytogenes isolates from a poultry-processing facility form more biofilm but do not have a greater resistance to disinfectants than sporadic strains. Pathogens 2019, 8, 250. [Google Scholar] [CrossRef]
- Cadena, M.; Kelman, T.; Marco, M.L.; Pitesky, M.J.F. Understanding antimicrobial resistance (AMR) profiles of Salmonella biofilm and planktonic bacteria challenged with disinfectants commonly used during poultry processing. Foods 2019, 8, 275. [Google Scholar] [CrossRef]
- Pang, X.; Song, X.; Chen, M.; Tian, S.; Lu, Z.; Sun, J.; Li, X.; Lu, Y.; Yuk, H.-G. Combating biofilms of foodborne pathogens with bacteriocins by lactic acid bacteria in the food industry. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1657–1676. [Google Scholar] [CrossRef]
- Uddin Mahamud, A.G.M.S.; Nahar, S.; Ashrafudoulla, M.; Park, S.H.; Ha, S.-D. Insights into antibiofilm mechanisms of phytochemicals: Prospects in the food industry. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, Y.; Li, J.; Wang, X.; He, S.; Yan, X.; Shi, Y.; Zhang, W.; Ding, L. Marine natural products and their synthetic analogs as promising antibiofilm agents for antibiotics discovery and development. Eur. J. Med. Chem. 2022, 239, 114513. [Google Scholar] [CrossRef]
- Martinengo, P.; Arunachalam, K.; Shi, C.J.F. Polyphenolic Antibacterials for Food Preservation: Review, Challenges, and Current Applications. Foods 2021, 10, 2469. [Google Scholar] [CrossRef]
- Chitlapilly Dass, S.; Wang, R. Biofilm through the Looking Glass: A Microbial Food Safety Perspective. Pathogens 2022, 11, 346. [Google Scholar] [CrossRef]
- Mevo, S.I.U.; Ashrafudoulla, M.; Furkanur Rahaman Mizan, M.; Park, S.H.; Ha, S.D.; Safety, F. Promising strategies to control persistent enemies: Some new technologies to combat biofilm in the food industry—A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5938–5964. [Google Scholar] [CrossRef]
- Chlebicz, A.; Śliżewska, K. Campylobacteriosis, salmonellosis, yersiniosis, and listeriosis as zoonotic foodborne diseases: A review. Int. J. Environ. Res. Public Health 2018, 15, 863. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.N.; Perry, K.J.; Regeimbal, J.M.; Regan, P.M.; Higgins, D.E. Identification of Listeria monocytogenes determinants required for biofilm formation. PLoS ONE 2014, 9, e113696. [Google Scholar] [CrossRef] [PubMed]
- Morgado, M.E.; Jiang, C.; Zambrana, J.; Upperman, C.R.; Mitchell, C.; Boyle, M.; Sapkota, A.R.; Sapkota, A.J.E.H. Climate change, extreme events, and increased risk of salmonellosis: Foodborne diseases active surveillance network (FoodNet), 2004–2014. Environ. Health 2021, 20, 105. [Google Scholar] [CrossRef] [PubMed]
- Manfreda, G.; De Cesare, A. Novel food trends and climate changes: Impact on emerging food-borne bacterial pathogens. Curr. Opin. Food Sci. 2016, 8, 99–103. [Google Scholar] [CrossRef]
- Ekici, G.; Dümen, E. Escherichia coli and food safety. In The Universe of Escherichia coli; IntechOpen: London, UK, 2019. [Google Scholar]
- Ageorges, V.; Monteiro, R.; Leroy, S.; Burgess, C.M.; Pizza, M.; Chaucheyras-durand, F.; Desvaux, M. Molecular determinants of surface colonisation in diarrhoeagenic Escherichia coli (DEC): From bacterial adhesion to biofilm formation. FEMS Microbiol. Rev. 2020, 44, 314–350. [Google Scholar] [CrossRef] [PubMed]
- Algharib, S.A.; Dawood, A.; Xie, S. Nanoparticles for treatment of bovine Staphylococcus aureus mastitis. Drug Deliv. 2020, 27, 292–308. [Google Scholar] [CrossRef]
- Arunachalam, K.; Pandurangan, P.; Shi, C.; Lagoa, R. Regulation of Staphylococcus aureus virulence and application of nanotherapeutics to eradicate s. aureus infection. Pharmaceutics 2023, 15, 310. [Google Scholar] [CrossRef]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Hansson, I.; Sandberg, M.; Habib, I.; Lowman, R.; Engvall, E.O. Knowledge gaps in control of Campylobacter for prevention of campylobacteriosis. Transbound. Emerg. Dis. 2018, 65, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Bai, X.; Nakatsu, C.H.; Bhunia, A.K. Bacterial biofilms and their implications in pathogenesis and food safety. Foods 2021, 10, 2117. [Google Scholar] [CrossRef] [PubMed]
- Sarno, E.; Pezzutto, D.; Rossi, M.; Liebana, E.; Rizzi, V. A review of significant European foodborne outbreaks in the last decade. J. Food Prot. 2021, 84, 2059–2070. [Google Scholar] [CrossRef]
- Lin, Y.; Briandet, R.; Kovács, Á.T. Bacillus cereus sensu lato biofilm formation and its ecological importance. Biofilm 2022, 4, 100070. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Hu, Q.; Xu, F.; Ding, S.-Y.; Zhu, K. Characterization of Bacillus cereus in dairy products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef]
- Carter, M.Q.; Feng, D.; Li, H.H. Curli fimbriae confer shiga toxin-producing Escherichia coli a competitive trait in mixed biofilms. Food Microbiol. 2019, 82, 482–488. [Google Scholar] [CrossRef]
- Yang, S.-C.; Lin, C.-H.; Aljuffali, I.A.; Fang, J.-Y. Current pathogenic Escherichia coli foodborne outbreak cases and therapy development. Arch. Microbiol. 2017, 199, 811–825. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of antibiotic resistance in Listeria monocytogenes isolated from food products: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef]
- Smith, A.M.; Tau, N.P.; Smouse, S.L.; Allam, M.; Ismail, A.; Ramalwa, N.R.; Disenyeng, B.; Ngomane, M.; Thomas, J. Outbreak of Listeria monocytogenes in South Africa, 2017–2018: Laboratory activities and experiences associated with whole-genome sequencing analysis of isolates. Foodborne Pathog. Dis. 2019, 16, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, J.; Chai, M.; Li, X.; Deng, Y.; Jin, Q.; Ji, J. Size and charge adaptive clustered nanoparticles targeting the biofilm microenvironment for chronic lung infection management. ACS Nano 2020, 14, 5686–5699. [Google Scholar] [CrossRef]
- Arteaga, V.; Lamas, A.; Regal, P.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Antimicrobial activity of apitoxin from Apis mellifera in Salmonella enterica strains isolated from poultry and its effects on motility, biofilm formation and gene expression. Microb. Pathog. 2019, 137, 103771. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Kannappan, A.; Shi, C.; Lin, X. Marine bacterial secondary metabolites: A treasure house for structurally unique and effective antimicrobial compounds. Mar. Drugs 2021, 19, 530. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.C.; de Paula, O.A.; Todorov, S.D.; Nero, L.A. In vitro evaluation of bacteriocins activity against Listeria monocytogenes biofilm formation. Appl. Biochem. Biotechnol. 2016, 178, 1239–1251. [Google Scholar] [CrossRef]
- Abdelhadi, I.M.A.; Sofy, A.R.; Hmed, A.A.; Refaey, E.E.; Soweha, H.E.; Abbas, M.A. Discovery of Polyvalent Myovirus (vB_STM-2) Phage as a natural antimicrobial system to lysis and biofilm removal of Salmonella Typhimurium Isolates from various food sources. Sustainability 2021, 13, 11602. [Google Scholar] [CrossRef]
- Martínez-Graciá, C.; González-Bermúdez, C.A.; Cabellero-Valcárcel, A.M.; Santaella-Pascual, M.; Frontela-Saseta, C. Use of herbs and spices for food preservation: Advantages and limitations. Curr. Opin. Food Sci. 2015, 6, 38–43. [Google Scholar] [CrossRef]
- Prakash, B.; Kedia, A.; Mishra, P.K.; Dubey, N.K. Plant essential oils as food preservatives to control moulds, mycotoxin contamination and oxidative deterioration of agri-food commodities–Potentials and challenges. Food Control 2015, 47, 381–391. [Google Scholar] [CrossRef]
- Sánchez, G.; Aznar, R.J.F.; Virology, E. Evaluation of natural compounds of plant origin for inactivation of enteric viruses. Food Environ. Virol. 2015, 7, 183–187. [Google Scholar] [CrossRef]
- Kannappan, A.; Santhakumari, S.; Srinivasan, R.; Pandian, S.K.; Ravi, A.V. Hemidesmus indicus, a traditional medicinal plant, targets the adherence of multidrug-resistant pathogens to form biofilms. Biocatal. Agric. Biotechnol. 2019, 21, 101338. [Google Scholar] [CrossRef]
- Srinivasan, R.; Devi, K.R.; Kannappan, A.; Pandian, S.K.; Ravi, A.V. Piper betle and its bioactive metabolite phytol mitigates quorum sensing mediated virulence factors and biofilm of nosocomial pathogen Serratia marcescens in vitro. J. Ethnopharmacol. 2016, 193, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Packiavathy, I.A.S.; Maruthamuthu, S.; Gnanaselvan, G.; Manoharan, S.; Paul, J.B.J.; Annapoorani, A.; Kannappan, A.; Ravi, A.V. The control of microbially induced corrosion by methyl eugenol–A dietary phytochemical with quorum sensing inhibitory potential. Bioelectrochemistry 2019, 128, 186–192. [Google Scholar] [CrossRef]
- Lahiri, D.; Dash, S.; Dutta, R.; Nag, M. Elucidating the effect of anti-biofilm activity of bioactive compounds extracted from plants. J. Biosci. 2019, 44, 52. [Google Scholar] [CrossRef]
- Čabarkapa, I.; Čolović, R.; Đuragić, O.; Popović, S.; Kokić, B.; Milanov, D.; Pezo, L. Anti-biofilm activities of essential oils rich in carvacrol and thymol against Salmonella Enteritidis. Biofouling 2019, 35, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Ashrafudoulla, M.; Rahaman Mizan, M.F.; Park, S.H.; Ha, S.-D. Antibiofilm activity of carvacrol against Listeria monocytogenes and Pseudomonas aeruginosa biofilm on MBEC™ biofilm device and polypropylene surface. LWT 2021, 147, 111575. [Google Scholar] [CrossRef]
- Rodrigues, J.B.D.S.; Souza, N.T.D.; Scarano, J.O.A.; Sousa, J.M.D.; Lira, M.C.; Figueiredo, R.C.B.Q.D.; de Souza, E.L.; Magnani, M. Efficacy of using oregano essential oil and carvacrol to remove young and mature Staphylococcus aureus biofilms on food-contact surfaces of stainless steel. LWT 2018, 93, 293–299. [Google Scholar] [CrossRef]
- Trevisan, D.A.C.; Silva, A.F.D.; Negri, M.; Abreu Filho, B.A.D.; Machinski Junior, M.; Patussi, E.V.; Campanerut-Sá, P.A.Z.; Mikcha, J.M.G. Antibacterial and antibiofilm activity of carvacrol against Salmonella enterica serotype Typhimurium. Braz. J. Pharm. Sci. 2018, 54, e17229. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum sensing and phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, T.; Wang, K.; Hou, C.; Cai, S.; Huang, Y.; Du, Z.; Huang, H.; Kong, J.; Chen, Y. Baicalein inhibits Staphylococcus aureus biofilm formation and the quorum sensing system in vitro. PLoS ONE 2016, 11, e0153468. [Google Scholar] [CrossRef]
- Silva, A.F.; dos Santos, A.R.; Coelho Trevisan, D.A.; Ribeiro, A.B.; Zanetti Campanerut-Sá, P.A.; Kukolj, C.; de Souza, E.M.; Cardoso, R.F.; Estivalet Svidzinski, T.I.; de Abreu Filho, B.A.; et al. Cinnamaldehyde induces changes in the protein profile of Salmonella Typhimurium biofilm. Res. Microbiol. 2018, 169, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Y.; Ding, X.; Wang, J.; Zhan, X. Inhibitory effects of reuterin on biofilm formation, quorum sensing and virulence genes of Clostridium perfringens. LWT 2022, 162, 113421. [Google Scholar] [CrossRef]
- Leiman, S.A.; May, J.M.; Lebar, M.D.; Kahne, D.; Kolter, R.; Losick, R. D-Amino acids indirectly inhibit biofilm formation in Bacillus subtilis by interfering with protein synthesis. J. Bacteriol. 2013, 195, 5391–5395. [Google Scholar] [CrossRef]
- Sanchez, C.J.; Akers, K.S.; Romano, D.R.; Woodbury, R.L.; Hardy, S.K.; Murray, C.K.; Wenke, J.C. D-Amino acids enhance the activity of antimicrobials against biofilms of clinical wound isolates of Staphylococcus aureus and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 4353–4361. [Google Scholar] [CrossRef]
- Shu, Q.; Lou, H.; Wei, T.; Liu, X.; Chen, Q. Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review. Pharmaceutics 2021, 13, 227. [Google Scholar] [CrossRef]
- Silveira, V.A.I.; Nishio, E.K.; Freitas, C.A.U.Q.; Amador, I.R.; Kobayashi, R.K.T.; Caretta, T.; Macedo, F.; Celligoi, M.A.P. Production and antimicrobial activity of sophorolipid against Clostridium perfringens and Campylobacter jejuni and their additive interaction with lactic acid. Biocatal. Agric. Biotechnol. 2019, 21, 101287. [Google Scholar] [CrossRef]
- Zhang, X.; Ashby, R.; Solaiman, D.K.Y.; Uknalis, J.; Fan, X. Inactivation of Salmonella spp. and Listeria spp. by palmitic, stearic, and oleic acid sophorolipids and thiamine dilauryl sulfate. Front. Microbiol. 2016, 7, 2076. [Google Scholar] [CrossRef]
- Elgamoudi, B.A.; Korolik, V. Campylobacter Biofilms: Potential of Natural Compounds to Disrupt Campylobacter jejuni Transmission. Int. J. Mol. Sci. 2021, 22, 12159. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Antibacterial Derivatives of Marine Algae: An Overview of Pharmacological Mechanisms and Applications. Mar. Drugs 2016, 14, 81. [Google Scholar] [CrossRef]
- Castillo, S.; Heredia, N.; García, S. 2(5H)-Furanone, epigallocatechin gallate, and a citric-based disinfectant disturb quorum-sensing activity and reduce motility and biofilm formation of Campylobacter jejuni. Folia Microbiol. 2015, 60, 89–95. [Google Scholar] [CrossRef]
- Cepas, V.; Gutiérrez-Del-Río, I.; López, Y.; Redondo-Blanco, S.; Gabasa, Y.; Iglesias, M.J.; Soengas, R.; Fernández-Lorenzo, A.; López-Ibáñez, S.; Villar, C.J.; et al. Microalgae and cyanobacteria strains as producers of lipids with antibacterial and antibiofilm activity. Mar. Drugs 2021, 19, 675. [Google Scholar] [CrossRef]
- Song, X.; Wang, L.; Liu, T.; Liu, Y.; Wu, X.; Liu, L. Mandarin (Citrus reticulata L.) essential oil incorporated into chitosan nanoparticles: Characterization, anti-biofilm properties and application in pork preservation. Int. J. Biol. Macromol. 2021, 185, 620–628. [Google Scholar] [CrossRef]
- Hakimi Alni, R.; Ghorban, K.; Dadmanesh, M. Combined effects of Allium sativum and Cuminum cyminum essential oils on planktonic and biofilm forms of Salmonella typhimurium isolates. 3 Biotech 2020, 10, 315. [Google Scholar] [CrossRef]
- Sharifi, A.; Nayeri Fasaei, B. Selected plant essential oils inhibit biofilm formation and luxS- and pfs-mediated quorum sensing by Escherichia coli O157:H7. Lett. Appl. Microbiol. 2022, 74, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Pelarti, S.; Karimi Zarehshuran, L.; Babaeekhou, L.; Ghane, M. Antibacterial, anti-biofilm and anti-quorum sensing activities of Artemisia dracunculus essential oil (EO): A study against Salmonella enterica serovar Typhimurium and Staphylococcus aureus. Arch. Microbiol. 2021, 203, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Karampoula, F.; Giaouris, E.; Deschamps, J.; Doulgeraki, A.I.; Nychas, G.J.; Dubois-Brissonnet, F. Hydrosol of Thymbra capitata is a highly efficient biocide against Salmonella enterica serovar Typhimurium biofilms. Appl. Environ. Microbiol. 2016, 82, 5309–5319. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Jin, W.; Wang, J.; Sun, Y.; Wu, X.; Liu, L. Antibacterial and anti-biofilm activities of peppermint essential oil against Staphylococcus aureus. LWT 2019, 101, 639–645. [Google Scholar] [CrossRef]
- Coimbra, A.; Carvalho, F.; Duarte, A.P.; Ferreira, S. Antimicrobial activity of Thymus zygis essential oil against Listeria monocytogenes and its application as food preservative. Innov. Food Sci. Emerg. Technol. 2022, 80, 103077. [Google Scholar] [CrossRef]
- Gao, Z.; Zhong, W.; Chen, K.; Tang, P.; Guo, J. Chemical composition and anti-biofilm activity of essential oil from Citrus medica L. var. sarcodactylis Swingle against Listeria monocytogenes. Ind. Crops Prod. 2020, 144, 112036. [Google Scholar] [CrossRef]
- Zhao, Y.; Qu, Y.; Tang, J.; Chen, J.; Liu, J. Tea catechin inhibits biofilm formation of methicillin-resistant S. aureus. J. Food Qual. 2021, 2021, 8873091. [Google Scholar] [CrossRef]
- Wang, B.; Wei, P.W.; Wan, S.; Yao, Y.; Song, C.R.; Song, P.P.; Xu, G.B.; Hu, Z.Q.; Zeng, Z.; Wang, C.; et al. Ginkgo biloba exocarp extracts inhibit S. aureus and MRSA by disrupting biofilms and affecting gene expression. J. Ethnopharmacol. 2021, 271, 113895. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M. Antimicrobial and Anti-Biofilm Activities of Citrus sinensis and Moringa oleifera against the pathogenic Pseudomonas aeruginosa and Staphylococcus aureus. Cureus 2020, 12, e12337. [Google Scholar] [CrossRef]
- Kannappan, A.; Gowrishankar, S.; Srinivasan, R.; Pandian, S.K.; Ravi, A.V. Antibiofilm activity of Vetiveria zizanioides root extract against methicillin-resistant Staphylococcus aureus. Microb. Pathog. 2017, 110, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Maia, D.S.V.; Haubert, L.; Kroning, I.S.; Soares, K.D.S.; Oliveira, T.L.; da Silva, W.P. Biofilm formation by Staphylococcus aureus isolated from food poisoning outbreaks and effect of Butia odorata Barb. Rodr. Extract on planktonic and biofilm cells. LWT 2020, 117, 108685. [Google Scholar] [CrossRef]
- Wu, B.; Liu, X.; Nakamoto, S.T.; Wall, M.; Li, Y. Antimicrobial activity of Ohelo Berry (Vaccinium calycinum) Juice against Listeria monocytogenes and its potential for milk preservation. Microorganisms 2022, 10, 548. [Google Scholar] [CrossRef] [PubMed]
- Banerji, R.; Mahamune, A.; Saroj, S.D. Aqueous extracts of spices inhibit biofilm in Listeria monocytogenes by downregulating release of eDNA. LWT 2022, 154, 112566. [Google Scholar] [CrossRef]
- Minami, M.; Takase, H.; Nakamura, M.; Makino, T. Methanol extract of Lonicera caerulea var. emphyllocalyx fruit has anti-motility and anti-biofilm activity against enteropathogenic Escherichia coli. Drug Discov. Ther. 2019, 13, 335–342. [Google Scholar] [CrossRef]
- Kim, G.; Xu, Y.J.; Farha, A.K.; Sui, Z.Q.; Corke, H. Bactericidal and antibiofilm properties of Rumex japonicus Houtt. on multidrug-resistant Staphylococcus aureus isolated from milk. J. Dairy Sci. 2022, 105, 2011–2024. [Google Scholar] [CrossRef]
- Kannappan, A.; Srinivasan, R.; Nivetha, A.; Annapoorani, A.; Pandian, S.K.; Ravi, A.V. Anti-virulence potential of 2-hydroxy-4-methoxybenzaldehyde against methicillin-resistant Staphylococcus aureus and its clinical isolates. Appl. Microbiol. Biotechnol. 2019, 103, 6747–6758. [Google Scholar] [CrossRef] [PubMed]
- Ming, D.; Wang, D.; Cao, F.; Xiang, H.; Mu, D.; Cao, J.; Li, B.; Zhong, L.; Dong, X.; Zhong, X.; et al. Kaempferol inhibits the primary attachment phase of biofilm formation in Staphylococcus aureus. Front. Microbiol. 2017, 8, 2263. [Google Scholar] [CrossRef]
- Bai, J.R.; Wu, Y.P.; Elena, G.; Zhong, K.; Gao, H. Insight into the effect of quinic acid on biofilm formed by Staphylococcus aureus. RSC Adv. 2019, 9, 3938–3945. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.H.; Wang, R.; Zhao, S.Q.; Chen, B.R.; Zeng, X.A. Inhibition of biofilm formation of foodborne Staphylococcus aureus by the citrus flavonoid naringenin. Foods 2021, 10, 2614. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.; Yu, H.H.; Kim, Y.J.; Lee, N.-K.; Paik, H.-D. The use of papain for the removal of biofilms formed by pathogenic Staphylococcus aureus and Campylobacter jejuni. LWT 2020, 127, 109383. [Google Scholar] [CrossRef]
- Wei, L.N.; Shi, C.Z.; Luo, C.X.; Hu, C.Y.; Meng, Y.H. Phloretin inhibits biofilm formation by affecting quorum sensing under different temperature. LWT 2020, 131, 109668. [Google Scholar] [CrossRef]
- Yum, S.-j.; Kwon, J.H.; Lee, K.-T.; Park, J.-T.; Jeong, H.-G. Efficacy of pristimerin against Staphylococcus aureus planktonic cultures and biofilms. LWT 2022, 164, 113627. [Google Scholar] [CrossRef]
- Kim, Y.K.; Roy, P.K.; Ashrafudoulla, M.; Nahar, S.; Toushik, S.H.; Hossain, M.I.; Mizan, M.F.R.; Park, S.H.; Ha, S.-D. Antibiofilm effects of quercetin against Salmonella enterica biofilm formation and virulence, stress response, and quorum-sensing gene expression. Food Control 2022, 137, 108964. [Google Scholar] [CrossRef]
- Al-Shabib, N.A.; Husain, F.M.; Ahmad, I.; Khan, M.S.; Khan, R.A.; Khan, J.M. Rutin inhibits mono and multi-species biofilm formation by foodborne drug resistant Escherichia coli and Staphylococcus aureus. Food Control 2017, 79, 325–332. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Park, I.; Khan, I.; Alshammari, F.H.; Kumar, P.; Chen, L.; Na, M.; Huh, Y.S.; Han, Y.-K.; Shukla, S. (−)-Tetrahydroberberrubine∙acetate accelerates antioxidant potential and inhibits food associated Bacillus cereus in rice. Food Chem. 2021, 339, 127902. [Google Scholar] [CrossRef]
- Zhao, Q.; Shi, Y.; Wang, X.; Huang, A. Characterization of a novel antimicrobial peptide from buffalo casein hydrolysate based on live bacteria adsorption. J. Dairy Sci. 2020, 103, 11116–11128. [Google Scholar] [CrossRef]
- Yang, S.; Yuan, Z.; Aweya, J.J.; Deng, S.; Weng, W.; Zhang, Y.; Liu, G. Antibacterial and antibiofilm activity of peptide PvGBP2 against pathogenic bacteria that contaminate Auricularia auricular culture bags. Food Sci. Hum. Wellness 2022, 11, 1607–1613. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, M.; Chen, Y.; Ouyang, J.; Wang, Y.; Yang, H.; Luo, X.; Zhang, D.; Lu, Y.; Yu, H.; et al. Antimicrobial, anti-biofilm properties of three naturally occurring antimicrobial peptides against spoilage bacteria, and their synergistic effect with chemical preservatives in food storage. Food Control 2021, 123, 107729. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kang, S.-S. Anti-Biofilm Activities of Manuka Honey against Escherichia coli O157:H7. Food Sci. Anim. Resour. 2020, 40, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Iodice, C.; Tommonaro, G.; Stanimirovic, B.; Ciric, A.; Glamoclija, J.; Nikolic, M.; De Rosa, S.; Sokovic, M. Further in vitro evaluation of antimicrobial activity of the marine sesquiterpene hydroquinone avarol. Curr. Pharm. Biotechnol. 2014, 15, 583–588. [Google Scholar] [CrossRef]
- Melian, C.; Segli, F.; Gonzalez, R.; Vignolo, G.; Castellano, P. Lactocin AL705 as quorum sensing inhibitor to control Listeria monocytogenes biofilm formation. J. Appl. Microbiol. 2019, 127, 911–920. [Google Scholar] [CrossRef]
- Chopra, L.; Singh, G.; Kumar Jena, K.; Sahoo, D.K. Sonorensin: A new bacteriocin with potential of an anti-biofilm agent and a food biopreservative. Sci. Rep. 2015, 5, 13412. [Google Scholar] [CrossRef]
- Wang, L.; Pang, X.; Zhao, J.; Jin, H.; Yang, X.; Fu, S.; Cheng, S.; Li, H.; Miao, C.; Man, C.; et al. Isolation and characteristics of new phage JK004 and application to control Cronobacter sakazakii on material surfaces and powdered infant formula. LWT 2022, 153, 112571. [Google Scholar] [CrossRef]
- Hintz, T.; Matthews, K.K.; Di, R. The Use of Plant Antimicrobial Compounds for Food Preservation. BioMed Res. Int. 2015, 2015, 246264. [Google Scholar] [CrossRef]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta (BBA) Biomembr. 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Driessen, A.J.M.; van den Hooven, H.W.; Kuiper, W.; Van de Camp, M.; Sahl, H.-G.; Konings, R.N.H.; Konings, W.N. Mechanistic studies of lantibiotic-induced permeabilization of phospholipid vesicles. Biochemistry 1995, 34, 1606–1614. [Google Scholar] [CrossRef] [Green Version]
- Brewer, M.S. Natural Antioxidants: Sources, Compounds, Mechanisms of Action, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Alvarez, M.; Moreira, M.d.R.; Roura, S.; Ayala-Zavala, J.; González-Aguilar, G.A. Using Natural Antimicrobials to Enhance the Safety and Quality of Fresh and Processed Fruits and Vegetables: TYPES of Antimicrobials. In Handbook of Natural Antimicrobials for Food Safety and Quality; Woodhead Publishing: Cambridge, UK, 2015; pp. 287–313. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxidative Med. Cell. Longev. 2009, 2, 897484. [Google Scholar] [CrossRef]
- González-Manzano, S.; Dueñas, M. Applications of Natural Products in Food. Foods 2021, 10, 300. [Google Scholar] [CrossRef]
- Bazinet, L.; Araya-Farias, M.; Doyen, A.; Trudel, D.; Têtu, B. Effect of process unit operations and long-term storage on catechin contents in EGCG-enriched tea drink. Food Res. Int. 2010, 43, 1692–1701. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, F.X.; Chan, J.M.; Wang, A.Z.; Langer, R.S.; Farokhzad, O.C. Nanoparticles in medicine: Therapeutic applications and developments. Clin. Pharmacol. Ther. 2008, 83, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Slavin, Y.N.; Asnis, J.; Häfeli, U.O.; Bach, H. Metal nanoparticles: Understanding the mechanisms behind antibacterial activity. J. Nanobiotechnol. 2017, 15, 65. [Google Scholar] [CrossRef]
- Singh, T.; Shukla, S.; Kumar, P.; Wahla, V.; Bajpai, V.K. Application of nanotechnology in food science: Perception and overview. Front. Microbiol. 2017, 8, 1501. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Delshadi, R.; Cacciotti, I.; Esfanjani, A.F.; Rezaei, A.; Tarhan, O.; Lee, C.C.; Assadpour, E.; Tomas, M.; Vahapoglu, B.; et al. Targeting foodborne pathogens via surface-functionalized nano-antimicrobials. Adv. Colloid Interface Sci. 2022, 302, 102622. [Google Scholar] [CrossRef] [PubMed]
- Pacha, P.A.; Munoz, M.A.; González-Rocha, G.; San Martín, I.; Quezada-Aguiluz, M.; Aguayo-Reyes, A.; Bello-Toledo, H.; Latorre, A.A. Molecular diversity of Staphylococcus aureus and the role of milking equipment adherences or biofilm as a source for bulk tank milk contamination. J. Dairy Sci. 2021, 104, 3522–3531. [Google Scholar] [CrossRef]
- Sharaf, M.; Sewid, A.H.; Hamouda, H.I.; Elharrif, M.G.; El-Demerdash, A.S.; Alharthi, A.; Hashim, N.; Hamad, A.A.; Selim, S.; Alkhalifah, D.H.M.; et al. Rhamnolipid-coated iron oxide nanoparticles as a novel multitarget candidate against major foodborne E. coli serotypes and methicillin-resistant S. aureus. Microbiol. Spectr. 2022, 10, e0025022. [Google Scholar] [CrossRef]
- Devlin, H.; Hiebner, D.; Barros, C.; Fulaz, S.; Quinn, L.; Vitale, S.; Casey, E. A high throughput method to investigate nanoparticle entrapment efficiencies in biofilms. Colloids Surf. B Biointerfaces 2020, 193, 111123. [Google Scholar] [CrossRef]
- Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Nanoparticle-biofilm interactions: The role of the EPS matrix. Trends Microbiol. 2019, 27, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Ikuma, K.; Decho, A.W.; Lau, B.L.T. When nanoparticles meet biofilms—Interactions guiding the environmental fate and accumulation of nanoparticles. Front. Microbiol. 2015, 6, 591. [Google Scholar] [CrossRef] [PubMed]
- Barros, C.H.N.; Devlin, H.; Hiebner, D.W.; Vitale, S.; Quinn, L.; Casey, E. Enhancing curcumin’s solubility and antibiofilm activity via silica surface modification. Nanoscale Adv. 2020, 2, 1694–1708. [Google Scholar] [CrossRef]
- Menon, S.; Rajeshkumar, S.; Kumar, V. A review on biogenic synthesis of gold nanoparticles, characterization, and its applications. Resour. Effic. Technol. 2017, 3, 516–527. [Google Scholar] [CrossRef]
- Sharma, D.; Kanchi, S.; Bisetty, K. Biogenic synthesis of nanoparticles: A review. Arab. J. Chem. 2019, 12, 3576–3600. [Google Scholar] [CrossRef]
- Muthulakshmi, L.; Suganya, K.; Murugan, M.; Annaraj, J.; Duraipandiyan, V.; Al Farraj, D.A.; Elshikh, M.S.; Juliet, A.; Pasupuleti, M.; Arockiaraj, J. Antibiofilm efficacy of novel biogenic silver nanoparticles from Terminalia catappa against food-borne Listeria monocytogenes ATCC 15,313 and mechanisms investigation in-vivo and in-vitro. J. King Saud Univ. Sci. 2022, 34, 102083. [Google Scholar] [CrossRef]
- Lotha, R.; Sundaramoorthy, N.S.; Shamprasad, B.R.; Nagarajan, S.; Sivasubramanian, A. Plant nutraceuticals (Quercetrin and Afzelin) capped silver nanoparticles exert potent antibiofilm effect against food borne pathogen Salmonella enterica serovar Typhi and curtail planktonic growth in zebrafish infection model. Microb. Pathog. 2018, 120, 109–118. [Google Scholar] [CrossRef]
- Zahoor, M.; Nazir, N.; Iftikhar, M.; Naz, S.; Zekker, I.; Burlakovs, J.; Uddin, F.; Kamran, A.W.; Kallistova, A.; Pimenov, N.; et al. A review on silver nanoparticles: Classification, various methods of synthesis, and their potential roles in biomedical applications and water treatment. Water 2021, 13, 2216. [Google Scholar] [CrossRef]
- Lotha, R.; Shamprasad, B.R.; Sundaramoorthy, N.S.; Nagarajan, S.; Sivasubramanian, A. Biogenic phytochemicals (cassinopin and isoquercetin) capped copper nanoparticles (ISQ/CAS@CuNPs) inhibits MRSA biofilms. Microb. Pathog. 2019, 132, 178–187. [Google Scholar] [CrossRef]
- Ali, K.; Ahmed, B.; Ansari, S.M.; Saquib, Q.; Al-Khedhairy, A.A.; Dwivedi, S.; Alshaeri, M.; Khan, M.S.; Musarrat, J. Comparative in situ ROS mediated killing of bacteria with bulk analogue, Eucalyptus leaf extract (ELE)-capped and bare surface copper oxide nanoparticles. Mater. Sci. Eng. C 2019, 100, 747–758. [Google Scholar] [CrossRef]
- Khan, F.; Park, S.-K.; Bamunuarachchi, N.I.; Oh, D.; Kim, Y.-M. Caffeine-loaded gold nanoparticles: Antibiofilm and anti-persister activities against pathogenic bacteria. Appl. Microbiol. Biotechnol. 2021, 105, 3717–3731. [Google Scholar] [CrossRef]
- Perveen, K.; Husain, F.M.; Qais, F.A.; Khan, A.; Razak, S.; Afsar, T.; Alam, P.; Almajwal, A.M.; Abulmeaty, M.M.A. Microwave-assisted rapid green synthesis of gold nanoparticles using seed extract of Trachyspermum ammi: ROS mediated biofilm inhibition and anticancer activity. Biomolecules 2021, 11, 197. [Google Scholar] [CrossRef]
- Malaikozhundan, B.; Lakshmi, V.N.; Krishnamoorthi, R. Copper oxide nanoparticles using Mentha spicata leaves as antibacterial, antibiofilm, free radical scavenging agent and efficient photocatalyst to degrade methylene blue dyes. Mater. Today Commun. 2022, 33, 104348. [Google Scholar] [CrossRef]
- Shamprasad, B.R.; Lotha, R.; Nagarajan, S.; Sivasubramanian, A. Metal nanoparticles functionalized with nutraceutical Kaempferitrin from edible Crotalaria juncea, exert potent antimicrobial and antibiofilm effects against Methicillin-resistant Staphylococcus aureus. Sci. Rep. 2022, 12, 7061. [Google Scholar] [CrossRef] [PubMed]
- El-Ghorab, A.H.; Behery, F.A.; Abdelgawad, M.A.; Alsohaimi, I.H.; Musa, A.; Mostafa, E.M.; Altaleb, H.A.; Althobaiti, I.O.; Hamza, M.; Elkomy, M.H.; et al. LC/MS profiling and gold nanoparticle formulation of major metabolites from Origanum majorana as antibacterial and antioxidant potentialities. Plants 2022, 11, 1871. [Google Scholar] [CrossRef] [PubMed]
- Senthamarai, M.D.; Malaikozhundan, B. Synergistic action of zinc oxide nanoparticle using the unripe fruit extract of Aegle marmelos (L.)—Antibacterial, antibiofilm, radical scavenging and ecotoxicological effects. Mater. Today Commun. 2022, 30, 103228. [Google Scholar] [CrossRef]
- Adebayo-Tayo, B.; Salaam, A.; Ajibade, A. Green synthesis of silver nanoparticle using Oscillatoria sp. extract, its antibacterial, antibiofilm potential and cytotoxicity activity. Heliyon 2019, 5, e02502. [Google Scholar] [CrossRef]
- Hayat, S.; Ashraf, A.; Zubair, M.; Aslam, B.; Siddique, M.H.; Khurshid, M.; Saqalein, M.; Khan, A.M.; Almatroudi, A.; Naeem, Z.; et al. Biofabrication of ZnO nanoparticles using Acacia arabica leaf extract and their antibiofilm and antioxidant potential against foodborne pathogens. PLoS ONE 2022, 17, e0259190. [Google Scholar] [CrossRef]
- Almatroudi, A.; Khadri, H.; Azam, M.; Rahmani, A.H.; Al Khaleefah, F.K.; Khateef, R.; Ansari, M.A.; Allemailem, K.S. Antibacterial, antibiofilm and anticancer activity of biologically synthesized silver nanoparticles using seed extract of Nigella sativa. Processes 2020, 8, 388. [Google Scholar] [CrossRef] [Green Version]
- Gherasim, O.; Popescu, R.C.; Grumezescu, V.; Mogoșanu, G.D.; Mogoantă, L.; Iordache, F.; Holban, A.M.; Vasile, B.Ș.; Bîrcă, A.C.; Oprea, O.-C.; et al. MAPLE coatings embedded with essential oil-conjugated magnetite for anti-biofilm applications. Materials 2021, 14, 1612. [Google Scholar] [CrossRef]
- Begines, B.; Ortiz, T.; Pérez-Aranda, M.; Martínez, G.; Merinero, M.; Argüelles-Arias, F.; Alcudia, A. Polymeric nanoparticles for drug delivery: Recent developments and future prospects. Nanomaterials 2020, 10, 1403. [Google Scholar] [CrossRef] [PubMed]
- Avramović, N.; Mandić, B.; Savić-Radojević, A.; Simić, T. Polymeric nanocarriers of drug delivery systems in cancer therapy. Pharmaceutics 2020, 12, 298. [Google Scholar] [CrossRef] [PubMed]
- Elbialy, N.S.; Aboushoushah, S.F.; Sofi, B.F.; Noorwali, A. Multifunctional curcumin-loaded mesoporous silica nanoparticles for cancer chemoprevention and therapy. Microporous Mesoporous Mater. 2020, 291, 109540. [Google Scholar] [CrossRef]
- Di, J.; Kim, J.; Hu, Q.; Jiang, X.; Gu, Z. Spatiotemporal drug delivery using laser-generated-focused ultrasound system. J. Control. Release 2015, 220, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Sirivisoot, S.; Harrison, B.S. Magnetically stimulated ciprofloxacin release from polymeric microspheres entrapping iron oxide nanoparticles. Int. J. Nanomed. 2015, 10, 4447–4458. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Gao, N.; Ren, J.; Qu, X. Transmutation of personal glucose meters into portable and highly sensitive microbial pathogen detection platform. Small 2015, 11, 4970–4975. [Google Scholar] [CrossRef]
- Gunasekaran, T.; Haile, T.; Nigusse, T.; Dhanaraju, M.D. Nanotechnology: An effective tool for enhancing bioavailability and bioactivity of phytomedicine. Asian Pac. J. Trop. Biomed. 2014, 4, S1–S7. [Google Scholar] [CrossRef]
- Pircalabioru, G.G.; Chifiriuc, M.-C. Nanoparticulate drug-delivery systems for fighting microbial biofilms: From bench to bedside. Future Microbiol. 2020, 15, 679–698. [Google Scholar] [CrossRef]
- Lee, N.-Y.; Ko, W.-C.; Hsueh, P.-R. Nanoparticles in the treatment of infections caused by multidrug-resistant organisms. Front. Pharmacol. 2019, 10, 1153. [Google Scholar] [CrossRef] [Green Version]
- Michalak, G.; Głuszek, K.; Piktel, E.; Deptuła, P.; Puszkarz, I.; Niemirowicz, K.; Bucki, R. Polymeric nanoparticles—A novel solution for delivery of antimicrobial agents. Med. Stud. 2016, 32, 56–62. [Google Scholar] [CrossRef]
- Perinelli, D.R.; Fagioli, L.; Campana, R.; Lam, J.K.W.; Baffone, W.; Palmieri, G.F.; Casettari, L.; Bonacucina, G. Chitosan-based nanosystems and their exploited antimicrobial activity. Eur. J. Pharm. Sci. 2018, 117, 8–20. [Google Scholar] [CrossRef]
- Siddhardha, B.; Pandey, U.; Kaviyarasu, K.; Pala, R.; Syed, A.; Bahkali, A.H.; Elgorban, A.M. Chrysin-loaded chitosan nanoparticles potentiates antibiofilm activity against Staphylococcus aureus. Pathogens 2020, 9, 115. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lin, Q.; Sheng, M.; Ding, T.; Li, B.; Gao, Y.; Tan, Y. Antibiofilm effect of cinnamaldehyde-chitosan nanoparticles against the biofilm of Staphylococcus aureus. Antibiotics 2022, 11, 1403. [Google Scholar] [CrossRef]
- Liu, T.; Liu, L. Fabrication and characterization of chitosan nanoemulsions loading thymol or thyme essential oil for the preservation of refrigerated pork. Int. J. Biol. Macromol. 2020, 162, 1509–1515. [Google Scholar] [CrossRef]
- Granata, G.; Stracquadanio, S.; Leonardi, M.; Napoli, E.; Malandrino, G.; Cafiso, V.; Stefani, S.; Geraci, C. Oregano and thyme essential oils encapsulated in chitosan nanoparticles as effective antimicrobial agents against foodborne pathogens. Molecules 2021, 26, 4055. [Google Scholar] [CrossRef]
- Cai, M.; Wang, Y.; Wang, R.; Li, M.; Zhang, W.; Yu, J.; Hua, R. Antibacterial and antibiofilm activities of chitosan nanoparticles loaded with Ocimum basilicum L. essential oil. Int. J. Biol. Macromol. 2022, 202, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Yu, H.; Kim, Y.-M. Bactericidal activity of usnic acid-chitosan nanoparticles against persister cells of biofilm-forming pathogenic bacteria. Mar. Drugs 2020, 18, 270. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, A.; Riviello, A.; Crocetta, V.; Giuseppe, F.D.; Pomponio, S.; Sulpizio, M.; Ilio, C.D.; Angelucci, S.; Barone, L.; Giulio, A.D.; et al. Evaluation of antibacterial and antibiofilm mechanisms by usnic acid against methicillin-resistant Staphylococcus aureus. Future Microbiol. 2016, 11, 1315–1338. [Google Scholar] [CrossRef]
- Folch-Cano, C.; Yazdani-Pedram, M.; Olea-Azar, C. Inclusion and functionalization of polymers with cyclodextrins: Current applications and future prospects. Molecules 2014, 19, 14066–14079. [Google Scholar] [CrossRef] [Green Version]
- Mohandoss, S.; Atchudan, R.; Edison, T.N.J.I.; Mishra, K.; Tamargo, R.J.I.; Palanisamy, S.; Yelithao, K.; You, S.; Napoleon, A.A.; Lee, Y.R. Enhancement of solubility, antibiofilm, and antioxidant activity of uridine by inclusion in β-cyclodextrin derivatives. J. Mol. Liq. 2020, 306, 112849. [Google Scholar] [CrossRef]
- Leclercq, L.; Tessier, J.; Douyère, G.; Nardello-Rataj, V.; Schmitzer, A.R. Phytochemical- and cyclodextrin-based pickering emulsions: Natural potentiators of antibacterial, antifungal, and antibiofilm activity. Langmuir 2020, 36, 4317–4323. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Garcia-Fernandez, M.J.; Lenoir, J.; De Meyer, L.; Remon, J.-P.; De Beer, T.; Concheiro, A.; Alvarez-Lorenzo, C.; Coenye, T. Dressings Loaded with cyclodextrin–hamamelitannin complexes increase Staphylococcus aureus susceptibility toward antibiotics both in single as well as in mixed biofilm communities. Macromol. Biosci. 2016, 16, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Huh, A.J.; Kwon, Y.J. “Nanoantibiotics”: A new paradigm for treating infectious diseases using nanomaterials in the antibiotics resistant era. J. Control. Release 2011, 156, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.S. Dendrimer nanotechnology for enhanced formulation and controlled delivery of resveratrol. Ann. N. Y. Acad. Sci. 2015, 1348, 134–140. [Google Scholar] [CrossRef]
- Wang, L.; Xu, X.; Zhang, Y.; Zhang, Y.; Zhu, Y.; Shi, J.; Sun, Y.; Huang, Q. Encapsulation of curcumin within poly(amidoamine) dendrimers for delivery to cancer cells. J. Mater. Sci. Mater. Med. 2013, 24, 2137–2144. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Caviglia, D. Prevention and eradication of biofilm by dendrimers: A possibility still little explored. Pharmaceutics 2022, 14, 2016. [Google Scholar] [CrossRef]
- Klajnert, B.; Janiszewska, J.; Urbanczyk-Lipkowska, Z.; Bryszewska, M.; Shcharbin, D.; Labieniec, M. Biological properties of low molecular mass peptide dendrimers. Int. J. Pharm. 2006, 309, 208–217. [Google Scholar] [CrossRef]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Préat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef]
- Gaspar, L.M.d.A.C.; Dórea, A.C.S.; Droppa-Almeida, D.; de Mélo Silva, I.S.; Montoro, F.E.; Alves, L.L.; Macedo, M.L.H.; Padilha, F.F. Development and characterization of PLGA nanoparticles containing antibiotics. J. Nanoparticle Res. 2018, 20, 289. [Google Scholar] [CrossRef]
- Anjum, A.; Chung, P.-Y.; Ng, S.-F. PLGA/xylitol nanoparticles enhance antibiofilm activity via penetration into biofilm extracellular polymeric substances. RSC Adv. 2019, 9, 14198–14208. [Google Scholar] [CrossRef]
- Da Costa, D.; Exbrayat-Héritier, C.; Rambaud, B.; Megy, S.; Terreux, R.; Verrier, B.; Primard, C. Surface charge modulation of rifampicin-loaded PLA nanoparticles to improve antibiotic delivery in Staphylococcus aureus biofilms. J. Nanobiotechnol. 2021, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- del Pozo-Rodríguez, A.; Solinís, M.Á.; Rodríguez-Gascón, A. Applications of lipid nanoparticles in gene therapy. Eur. J. Pharm. Biopharm. 2016, 109, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.; Muiruri, P.W.; Jones, G.H.; Scott, M.J.; Jones, M.N. The effect of grafted poly(ethylene glycol) on the electrophoretic properties of phospholipid liposomes and their adsorption to bacterial biofilms. Colloids Surf. A Physicochem. Eng. Asp. 2001, 194, 287–296. [Google Scholar] [CrossRef]
- Wang, D.-Y.; van der Mei, H.C.; Ren, Y.; Busscher, H.J.; Shi, L. Lipid-based antimicrobial delivery-systems for the treatment of bacterial infections. Front. Chem. 2020, 7, 872. [Google Scholar] [CrossRef]
- Ribeiro, L.N.d.M.; de Paula, E.; Rossi, D.A.; Martins, F.A.; de Melo, R.T.; Monteiro, G.P.; Breitkreitz, M.C.; Goulart, L.R.; Fonseca, B.B. Nanocarriers from natural lipids with in vitro activity against Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2021, 10, 571040. [Google Scholar] [CrossRef] [PubMed]
- Filipczak, N.; Pan, J.; Yalamarty, S.S.K.; Torchilin, V.P. Recent advancements in liposome technology. Adv. Drug Deliv. Rev. 2020, 156, 4–22. [Google Scholar] [CrossRef]
- Wang, Y. Liposome as a delivery system for the treatment of biofilm-mediated infections. J. Appl. Microbiol. 2021, 131, 2626–2639. [Google Scholar] [CrossRef]
- Cui, H.; Li, W.; Li, C.; Vittayapadung, S.; Lin, L. Liposome containing cinnamon oil with antibacterial activity against methicillin-resistant Staphylococcus aureus biofilm. Biofouling 2016, 32, 215–225. [Google Scholar] [CrossRef]
- Dos Santos Ramos, M.A.; Da Silva, P.B.; Spósito, L.; De Toledo, L.G.; Bonifácio, B.V.; Rodero, C.F.; Dos Santos, K.C.; Chorilli, M.; Bauab, T.M. Nanotechnology-based drug delivery systems for control of microbial biofilms: A review. Int. J. Nanomed. 2018, 13, 1179–1213. [Google Scholar] [CrossRef] [Green Version]
- Rukavina, Z.; Vanić, Ž. Current Trends in Development of Liposomes for Targeting Bacterial Biofilms. Pharmaceutics 2016, 8, 18. [Google Scholar] [CrossRef]
- Rossi, C.; Chaves-López, C.; Serio, A.; Casaccia, M.; Maggio, F.; Paparella, A. Effectiveness and mechanisms of essential oils for biofilm control on food-contact surfaces: An updated review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2172–2191. [Google Scholar] [CrossRef]
- Amrutha, B.; Sundar, K.; Shetty, P.H. Spice oil nanoemulsions: Potential natural inhibitors against pathogenic E. coli and Salmonella spp. from fresh fruits and vegetables. LWT Food Sci. Technol. 2017, 79, 152–159. [Google Scholar] [CrossRef]
- Prateeksha; Barik, S.K.; Singh, B.N. Nanoemulsion-loaded hydrogel coatings for inhibition of bacterial virulence and biofilm formation on solid surfaces. Sci. Rep. 2019, 9, 6520. [Google Scholar] [CrossRef]
- Wu, Y.; Long, Y.; Li, Q.-L.; Han, S.; Ma, J.; Yang, Y.-W.; Gao, H. Layer-by-layer (LBL) self-assembled biohybrid nanomaterials for efficient antibacterial applications. ACS Appl. Mater. Interfaces 2015, 7, 17255–17263. [Google Scholar] [CrossRef]
- Tang, F.; Li, L.; Chen, D. Mesoporous silica nanoparticles: Synthesis, biocompatibility and drug delivery. Adv. Mater. 2012, 24, 1504–1534. [Google Scholar] [CrossRef]
- Balaure, P.C.; Boarca, B.; Popescu, R.C.; Savu, D.; Trusca, R.; Vasile, B.Ș.; Grumezescu, A.M.; Holban, A.M.; Bolocan, A.; Andronescu, E. Bioactive mesoporous silica nanostructures with anti-microbial and anti-biofilm properties. Int. J. Pharm. 2017, 531, 35–46. [Google Scholar] [CrossRef]
- Li, J.; Zhong, W.; Zhang, K.; Wang, D.; Hu, J.; Chan-Park, M.B. Biguanide-derived polymeric nanoparticles kill MRSA biofilm and suppress infection in vivo. ACS Appl. Mater. Interfaces 2020, 12, 21231–21241. [Google Scholar] [CrossRef]
- Li, C.-H.; Chen, X.; Landis, R.F.; Geng, Y.; Makabenta, J.M.; Lemnios, W.; Gupta, A.; Rotello, V.M. Phytochemical-based nanocomposites for the treatment of bacterial biofilms. ACS Infect. Dis. 2019, 5, 1590–1596. [Google Scholar] [CrossRef]
- Cacciatore, F.A.; Dalmás, M.; Maders, C.; Ataíde Isaía, H.; Brandelli, A.; da Silva Malheiros, P. Carvacrol encapsulation into nanostructures: Characterization and antimicrobial activity against foodborne pathogens adhered to stainless steel. Food Res. Int. 2020, 133, 109143. [Google Scholar] [CrossRef]
- Perez, A.P.; Perez, N.; Lozano, C.M.S.; Altube, M.J.; de Farias, M.A.; Portugal, R.V.; Buzzola, F.; Morilla, M.J.; Romero, E.L. The anti MRSA biofilm activity of Thymus vulgaris essential oil in nanovesicles. Phytomedicine 2019, 57, 339–351. [Google Scholar] [CrossRef]
- Bhatia, E.; Sharma, S.; Jadhav, K.; Banerjee, R. Combinatorial liposomes of berberine and curcumin inhibit biofilm formation and intracellular methicillin resistant Staphylococcus aureus infections and associated inflammation. J. Mater. Chem. B 2021, 9, 864–875. [Google Scholar] [CrossRef]
- Pu, C.; Tang, W. The antibacterial and antibiofilm efficacies of a liposomal peptide originating from rice bran protein against Listeria monocytogenes. Food Funct. 2017, 8, 4159–4169. [Google Scholar] [CrossRef]
- Girish, V.M.; Liang, H.; Aguilan, J.T.; Nosanchuk, J.D.; Friedman, J.M.; Nacharaju, P. Anti-biofilm activity of garlic extract loaded nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2019, 20, 102009. [Google Scholar] [CrossRef]
- Luan, L.; Chi, Z.; Liu, C. Chinese white wax solid lipid nanoparticles as a novel nanocarrier of curcumin for inhibiting the formation of Staphylococcus aureus biofilms. Nanomaterials 2019, 9, 763. [Google Scholar] [CrossRef]
- Nemattalab, M.; Rohani, M.; Evazalipour, M.; Hesari, Z. Formulation of cinnamon (Cinnamomum verum) oil loaded solid lipid nanoparticles and evaluation of its antibacterial activity against multi-drug resistant Escherichia coli. BMC Complement. Med. Ther. 2022, 22, 289. [Google Scholar] [CrossRef]
- Nallasamy, P.; Ramalingam, T.; Nooruddin, T.; Shanmuganathan, R.; Arivalagan, P.; Natarajan, S. Polyherbal drug loaded starch nanoparticles as promising drug delivery system: Antimicrobial, antibiofilm and neuroprotective studies. Process Biochem. 2020, 92, 355–364. [Google Scholar] [CrossRef]
- Anjum, M.M.; Patel, K.K.; Dehari, D.; Pandey, N.; Tilak, R.; Agrawal, A.K.; Singh, S. Anacardic acid encapsulated solid lipid nanoparticles for Staphylococcus aureus biofilm therapy: Chitosan and DNase coating improves antimicrobial activity. Drug Deliv. Transl. Res. 2021, 11, 305–317. [Google Scholar] [CrossRef]
- Nosrati, M.; Ranjbar, R. Investigation of the antibacterial and biofilm inhibitory activities of Prangos acaulis (DC.) Bornm in nanoparticulated formulation. Nanotechnology 2022, 33, 385103. [Google Scholar] [CrossRef]
- Franklyne, J.S.; Andrew Ebenazer, L.; Mukherjee, A.; Natarajan, C. Cinnamon and clove oil nanoemulsions: Novel therapeutic options against vancomycin intermediate susceptible Staphylococcus aureus. Appl. Nanosci. 2019, 9, 1405–1415. [Google Scholar] [CrossRef]
- Thorn, C.R.; Carvalho-Wodarz, C.d.S.; Horstmann, J.C.; Lehr, C.-M.; Prestidge, C.A.; Thomas, N. Tobramycin liquid crystal nanoparticles eradicate cystic fibrosis-related Pseudomonas aeruginosa biofilms. Small 2021, 17, 2100531. [Google Scholar] [CrossRef]
- Dalcin, A.J.F.; Santos, C.G.; Gündel, S.S.; Roggia, I.; Raffin, R.P.; Ourique, A.F.; Santos, R.C.V.; Gomes, P. Anti biofilm effect of dihydromyricetin-loaded nanocapsules on urinary catheter infected by Pseudomonas aeruginosa. Colloids Surf. B Biointerfaces 2017, 156, 282–291. [Google Scholar] [CrossRef]
- Lima, R.A.; de Souza, S.L.X.; Lima, L.A.; Batista, A.L.X.; de Araújo, J.T.C.; Sousa, F.F.O.; Rolim, J.; Bandeira, T. Antimicrobial effect of anacardic acid-loaded zein nanoparticles loaded on Streptococcus mutans biofilms. Braz. J. Microbiol. 2020, 51, 1623–1630. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, L.; Su, L.; van der Mei, H.C.; Jutte, P.C.; Ren, Y.; Busscher, H.J. Nanotechnology-based antimicrobials and delivery systems for biofilm-infection control. Chem. Soc. Rev. 2019, 48, 428–446. [Google Scholar] [CrossRef]
- Peulen, T.O.; Wilkinson, K.J. Diffusion of nanoparticles in a biofilm. Environ. Sci. Technol. 2011, 45, 3367–3373. [Google Scholar] [CrossRef]
- MubarakAli, D.; Arunkumar, J.; Pooja, P.; Subramanian, G.; Thajuddin, N.; Alharbi, N.S. Synthesis and characterization of biocompatibility of tenorite nanoparticles and potential property against biofilm formation. Saudi Pharm. J. 2015, 23, 421–428. [Google Scholar] [CrossRef]
- Forier, K.; Messiaen, A.-S.; Raemdonck, K.; Nelis, H.; De Smedt, S.; Demeester, J.; Coenye, T.; Braeckmans, K. Probing the size limit for nanomedicine penetration into Burkholderia multivorans and Pseudomonas aeruginosa biofilms. J. Control. Release 2014, 195, 21–28. [Google Scholar] [CrossRef]
- Jiang, F.; Deng, Y.; Yeh, C.-K.; Sun, Y. Quaternized chitosans bind onto preexisting biofilms and eradicate pre-attached microorganisms. J. Mater. Chem. B 2014, 2, 8518–8527. [Google Scholar] [CrossRef]
- Fang, J.Y.; Chou, W.L.; Lin, C.F.; Sung, C.T.; Alalaiwe, A.; Yang, S.C. Facile biofilm penetration of cationic liposomes loaded with DNase I/Proteinase K to eradicate Cutibacterium acnes for treating cutaneous and catheter infections. Int. J. Nanomed. 2021, 16, 8121–8138. [Google Scholar] [CrossRef]
- Tan, Y.; Ma, S.; Liu, C.; Yu, W.; Han, F. Enhancing the stability and antibiofilm activity of DspB by immobilization on carboxymethyl chitosan nanoparticles. Microbiol. Res. 2015, 178, 35–41. [Google Scholar] [CrossRef]
- He, D.; Tan, Y.; Li, P.; Luo, Y.; Zhu, Y.; Yu, Y.; Chen, J.; Ning, N.; Zhang, S. Surface charge-convertible quaternary ammonium salt-based micelles for in vivo infection therapy. Chin. Chem. Lett. 2021, 32, 1743–1746. [Google Scholar] [CrossRef]
- Bandi, S.P.; Bhatnagar, S.; Venuganti, V.V.K. Advanced materials for drug delivery across mucosal barriers. Acta Biomater. 2021, 119, 13–29. [Google Scholar] [CrossRef]
- Horev, B.; Klein, M.I.; Hwang, G.; Li, Y.; Kim, D.; Koo, H.; Benoit, D.S. pH-activated nanoparticles for controlled topical delivery of farnesol to disrupt oral biofilm virulence. ACS Nano 2015, 9, 2390–2404. [Google Scholar] [CrossRef]
- Sims, K.R.; Liu, Y.; Hwang, G.; Jung, H.I.; Koo, H.; Benoit, D.S.W. Enhanced design and formulation of nanoparticles for anti-biofilm drug delivery. Nanoscale 2018, 11, 219–236. [Google Scholar] [CrossRef]
- Wang, Y.; Shukla, A. Bacteria-responsive biopolymer-coated nanoparticles for biofilm penetration and eradication. Biomater. Sci. 2022, 10, 2831–2843. [Google Scholar] [CrossRef]
- Liu, Y.; Ren, Y.; Li, Y.; Su, L.; Zhang, Y.; Huang, F.; Liu, J.; Liu, J.; van Kooten, T.G.; An, Y.; et al. Nanocarriers with conjugated antimicrobials to eradicate pathogenic biofilms evaluated in murine in vivo and human ex vivo infection models. Acta Biomater. 2018, 79, 331–343. [Google Scholar] [CrossRef]
- Ding, Y.; Hao, Y.; Yuan, Z.; Tao, B.; Chen, M.; Lin, C.; Liu, P.; Cai, K. A dual-functional implant with an enzyme-responsive effect for bacterial infection therapy and tissue regeneration. Biomater. Sci. 2020, 8, 1840–1854. [Google Scholar] [CrossRef]
- Vinchhi, P.; Rawal, S.U.; Patel, M.M. Chapter 13—External Stimuli-Responsive Drug Delivery Systems. In Drug Delivery Devices and Therapeutic Systems; Chappel, E., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 267–288. [Google Scholar]
- Wang, Y.; Lv, Q.; Chen, Y.; Xu, L.; Feng, M.; Xiong, Z.; Li, J.; Ren, J.; Liu, J.; Liu, B. Bilayer hydrogel dressing with lysozyme-enhanced photothermal therapy for biofilm eradication and accelerated chronic wound repair. Acta Pharm. Sin. B 2023, 13, 284–297. [Google Scholar] [CrossRef]
- Ding, M.; Zhao, W.; Song, L.-J.; Luan, S.-F. Stimuli-responsive nanocarriers for bacterial biofilm treatment. Rare Met. 2022, 41, 482–498. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Gupta, A.; Makabenta, J.M.V.; Park, J.; Amante, J.J.; Chattopadhyay, A.N.; Matuwana, D.; Kearney, C.J.; Rotello, V.M. Ultrasound-enhanced antibacterial activity of polymeric nanoparticles for eradicating bacterial biofilms. Adv. Healthc. Mater. 2022, 11, 2201060. [Google Scholar] [CrossRef]
- Ren, Y.; Liu, H.; Liu, X.; Zheng, Y.; Li, Z.; Li, C.; Yeung, K.W.K.; Zhu, S.; Liang, Y.; Cui, Z.; et al. Photoresponsive Materials for Antibacterial Applications. Cell Rep. Phys. Sci. 2020, 1, 100245. [Google Scholar] [CrossRef]
- Zhao, Y.; Dai, X.; Wei, X.; Yu, Y.; Chen, X.; Zhang, X.; Li, C. Near-infrared light-activated thermosensitive liposomes as efficient agents for photothermal and antibiotic synergistic therapy of bacterial biofilm. ACS Appl. Mater. Interfaces 2018, 10, 14426–14437. [Google Scholar] [CrossRef]
- Yu, Q.; Deng, T.; Lin, F.C.; Zhang, B.; Zink, J.I. Supramolecular assemblies of heterogeneous mesoporous silica nanoparticles to co-deliver antimicrobial peptides and antibiotics for synergistic eradication of pathogenic biofilms. ACS Nano 2020, 14, 5926–5937. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Thoendel, M.; Olakanmi, O.; Britigan, B.E.; Singh, P.K. The transition metal gallium disrupts Pseudomonas aeruginosa iron metabolism and has antimicrobial and antibiofilm activity. J. Clin. Investig. 2007, 117, 877–888. [Google Scholar] [CrossRef]
- Olakanmi, O.; Britigan, B.E.; Schlesinger, L.S. Gallium disrupts iron metabolism of mycobacteria residing within human macrophages. Infect. Immun. 2000, 68, 5619–5627. [Google Scholar] [CrossRef]
- Halwani, M.; Yebio, B.; Suntres, Z.E.; Alipour, M.; Azghani, A.O.; Omri, A. Co-encapsulation of gallium with gentamicin in liposomes enhances antimicrobial activity of gentamicin against Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2008, 62, 1291–1297. [Google Scholar] [CrossRef]
- Domenico, P.; Baldassarri, L.; Schoch, P.E.; Kaehler, K.; Sasatsu, M.; Cunha, B.A. Activities of bismuth thiols against staphylococci and staphylococcal biofilms. Antimicrob. Agents Chemother. 2001, 45, 1417–1421. [Google Scholar] [CrossRef]
- Domenico, P.; Reich, J.; Madonia, W.; Cunha, B.A. Resistance to bismuth among Gram-negative bacteria is dependent upon iron and its uptake. J. Antimicrob. Chemother. 1996, 38, 1031–1040. [Google Scholar] [CrossRef]
- Huang, C.T.; Stewart, P.S. Reduction of polysaccharide production in Pseudomonas aeruginosa biofilms by bismuth dimercaprol (BisBAL) treatment. J. Antimicrob. Chemother. 1999, 44, 601–605. [Google Scholar] [CrossRef]
- Folsom, J.P.; Baker, B.; Stewart, P.S. In vitro efficacy of bismuth thiols against biofilms formed by bacteria isolated from human chronic wounds. J. Appl. Microbiol. 2011, 111, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Halwani, M.; Blomme, S.; Suntres, Z.E.; Alipour, M.; Azghani, A.O.; Kumar, A.; Omri, A. Liposomal bismuth-ethanedithiol formulation enhances antimicrobial activity of tobramycin. Int. J. Pharm. 2008, 358, 278–284. [Google Scholar] [CrossRef]
- Guo, P.; Buttaro, B.A.; Xue, H.Y.; Tran, N.T.; Wong, H.L. Lipid-polymer hybrid nanoparticles carrying linezolid improve treatment of methicillin-resistant Staphylococcus aureus (MRSA) harbored inside bone cells and biofilms. Eur. J. Pharm. Biopharm. 2020, 151, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Shu, X.; Wang, Y.; Wang, D.; Wu, G.; Zhang, X.; Jin, Q.; Liu, J.; Wu, Z.; Xu, Z.; et al. Key progresses of MOE key laboratory of macromolecular synthesis and functionalization in 2020. Chin. Chem. Lett. 2022, 33, 1650–1658. [Google Scholar] [CrossRef]
- Yuan, Z.; Lin, C.; Dai, L.; He, Y.; Hu, J.; Xu, K.; Tao, B.; Liu, P.; Cai, K. Near-infrared light-activatable dual-action nanoparticle combats the established biofilms of methicillin-resistant Staphylococcus aureus and its accompanying inflammation. Small 2021, 17, e2007522. [Google Scholar] [CrossRef] [PubMed]
- de Annunzio, S.R.; de Freitas, L.M.; Blanco, A.L.; da Costa, M.M.; Carmona-Vargas, C.C.; de Oliveira, K.T.; Fontana, C.R. Susceptibility of Enterococcus faecalis and Propionibacterium acnes to antimicrobial photodynamic therapy. J. Photochem. Photobiol. B Biol. 2018, 178, 545–550. [Google Scholar] [CrossRef]
- Silva, A.F.; de Oliveira Silva, J.V.; Campanerut-Sá, P.A.Z.; Mikcha, J.M.G. Chapter 13—Photoinactivation of biofilms. In Recent Trends in Biofilm Science and Technology; Simoes, M., Borges, A., Chaves Simoes, L., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 295–306. [Google Scholar]
- Cossu, A.; Ercan, D.; Wang, Q.; Peer, W.A.; Nitin, N.; Tikekar, R.V. Antimicrobial effect of synergistic interaction between UV-A light and gallic acid against Escherichia coli O157:H7 in fresh produce wash water and biofilm. Innov. Food Sci. Emerg. Technol. 2016, 37, 44–52. [Google Scholar] [CrossRef]
- Parasuraman, P.; Antony, A.P.; Sharan, A.; Siddhardha, B.; Kasinathan, K.; Bahkali, N.A.; Dawoud, T.M.; Syed, A. Antimicrobial photodynamic activity of toluidine blue encapsulated in mesoporous silica nanoparticles against Pseudomonas aeruginosa and Staphylococcus aureus. Biofouling 2019, 35, 89–103. [Google Scholar] [CrossRef]
- Al-Mutairi, R.; Tovmasyan, A.; Batinic-Haberle, I.; Benov, L. Sublethal Photodynamic Treatment Does Not Lead to Development of Resistance. Front. Microbiol. 2018, 9, 1699. [Google Scholar] [CrossRef]
- Penders, J.; Stolzoff, M.; Hickey, D.J.; Andersson, M.; Webster, T.J. Shape-dependent antibacterial effects of non-cytotoxic gold nanoparticles. Int. J. Nanomed. 2017, 12, 2457–2468. [Google Scholar] [CrossRef]
- MubarakAli, D.; Arunachalam, K.; Lakshmanan, M.; Badar, B.; Kim, J.-W.; Lee, S.-Y. Unveiling antibiofilm property of hydroxyapatide on Pseudomonas aeruginosa: Synthesis and Strategy. Pharmaceutics 2023, 15, 463. [Google Scholar] [CrossRef]
- LewisOscar, F.; MubarakAli, D.; Nithya, C.; Priyanka, R.; Gopinath, V.; Alharbi, N.S.; Thajuddin, N. One pot synthesis and anti-biofilm potential of copper nanoparticles (CuNPs) against clinical strains of Pseudomonas aeruginosa. Biofouling 2015, 31, 379–391. [Google Scholar] [CrossRef]
- Chari, N.; Felix, L.; Davoodbasha, M.; Sulaiman Ali, A.; Nooruddin, T. In vitro and in vivo antibiofilm effect of copper nanoparticles against aquaculture pathogens. Biocatal. Agric. Biotechnol. 2017, 10, 336–341. [Google Scholar] [CrossRef]
- MubarakAli, D.; Manzoor, M.A.P.; Sabarinathan, A.; Anchana Devi, C.; Rekha, P.D.; Thajuddin, N.; Lee, S.-Y. An investigation of antibiofilm and cytotoxic property of MgO nanoparticles. Biocatal. Agric. Biotechnol. 2019, 18, 101069. [Google Scholar] [CrossRef]
- Singh, R.P. Chapter 11—Utility of Nanomaterials in Food Safety. In Food Safety and Human Health; Singh, R.L., Mondal, S., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 285–318. [Google Scholar]
- Brandelli, A. The interaction of nanostructured antimicrobials with biological systems: Cellular uptake, trafficking and potential toxicity. Food Sci. Hum. Wellness 2020, 9, 8–20. [Google Scholar] [CrossRef]
- Bahrami, A.; Delshadi, R.; Assadpour, E.; Jafari, S.M.; Williams, L. Antimicrobial-loaded nanocarriers for food packaging applications. Adv. Colloid Interface Sci. 2020, 278, 102140. [Google Scholar] [CrossRef] [PubMed]

| Foodborne Bacterial Pathogens | Selected Important Pathogenic Attributes and Essential Genes or Proteins Involved in Biofilm Formation | Favorable Conditions and Frequently Associated Food Products | Diseases Conditions | References |
|---|---|---|---|---|
| L. monocytogenes | Listerial adhesive proteins mediate adherence in L. monocytogenes. Further, internalins and phospholipases ensure their internalization in host cells. In addition, it produces other proteins like listeriolysin (causing infection) and actins (aiding mobility between host cells). Putative response regulator (DegU), flagellar proteins (FlaA and Mot), quorum sensing proteins (AgrBDCA), biofilm-associated protein (BapL), adhesin proteins (LAP), D-alanylation pathway (DtlABCD), internalins, phospholipases, phosphate-sensing two-component system (PhoPR), autoinducer-2 (LuxS), autolysin, SigB, relA, hpt, and SecA2 are reported to be involved in biofilm formation. | It can survive in harsh environments, such as varied pH (4.0–9.6) and temperature (−1.5 to 45 °C). It is frequently associated with dairy products, fresh produce, and ready-to-eat foods. | Ingestion of contaminated food leads to gastroenteritis or Invasive listeriosis. The invasive systemic disease targets immunocompromised hosts and lead to miscarriage and stillbirth in pregnant women. It can cause meningitis or encephalitis in newborns and the elderly. | [15,16,17] |
| Salmonella spp. | Adherence is mediated by the expression of proteins related to motility (fimbriae and flagella) and different adhesion proteins (BapA, SiiE, ShdA, MisL, and SadA). Bap and curli (CsgD) and cellulose (BcsA) biosynthesis are reported to be involved in biofilm formation. | It can grow at temperatures ranging between 8 and 45 °C and at a pH ranging between 4.0 and 9.5. Incidence, dissemination, and persistence of infections are often observed when consuming raw foods like fruits and vegetables. | Salmonella infection, often known as salmonellosis, is characterized by an early onset of fever, diarrhea, and stomach cramps within 12 to 72 h after exposure. | [18,19] |
| E. coli | Molecular determining factors responsible for surface colonization include exopolysaccharides, lipopolysaccharides, poly-N-acetyl glucosamine, colonic acid, lipoproteins (SslE), E. coli factor adherence 1 (Efa1), immunoglobulin binding protein (Eib), and fimbrial (AAF, Fim) adhesins. It also produces heat-stable (STa, STb) and heat-labile (LTp/h, LT-IIa, LT-IIb) enterotoxins. | E. coli thrives in aerobic and anaerobic conditions and temperatures ranging from 23 to 40 °C and at a pH of 6.5 to 7.5. Contaminated ground meat, dairy, and fresh vegetables are the causes of most outbreaks. | E. coli, a sign of feces in food and water, suggests inadequate hygiene. Food infected with heat-stable enterotoxins from pathogenic E. coli may induce food poisoning. | [20,21] |
| S. aureus | Produce enzymes that degrade phospholipids, elastin, DNA, and hyaluronic acid, and facilitate tissue penetration. Express different pore-forming toxins to evade host immune response, such as leukocidins, phenol-soluble modulins, protein A, Eap, staphyloxanthin, staphylococcal complement inhibitor, and capsular polysaccharides. Express toxins like Toxic shock syndrome toxin-1, α-toxin, enterotoxins, exfoliative toxins A and B, and lipoteichoic acid. Microbial surface components that recognize adhesive matrix molecules include surface proteins such as bone sialoprotein-binding protein, fibronectin-binding proteins, clumping factors, and fibrinogen-binding protein. Produce several proteins involved in biofilm formation, such as SasG, CidA, Bap, Spa, and polysaccharide intracellular adhesions (ica operon). | Grow at a temperature ranging from 15 to 45 °C, with pH ranging between 4.0 and 9.8. Staphylococcal food poisoning happens with consuming dairy products from animals suffering from S. aureus mastitis. In addition, there are a lot of people who carry the bacteria. This is primarily because of improper food handling. | Food pathogenic S. aureus strains produce enterotoxins. Due to their stability, heat tolerance, and capacity to withstand freezing and drying, these enterotoxins are a significant food business risk. These enterotoxins cause staphylococcal food poisoning outbreaks when consumed. | [22,23] |
| Campylobacter spp. | Express CadF, JlpA, PEB1, and CapA for different surface adherence. Express a wide range of flagellar protein required for host cell adhesion, biofilm formation, and secretion of invasive proteins that suppresses the host immune response. In addition, they produce toxins (CdtA, CdtB, and CdtC). | Grow between 37 °C and 42 °C (optimally at 41.5 °C), with fresh chicken meat as the most implicated food type. | Among the Campylobacter spp., C. jejuni and C. coli are the most significant. Campylobacteriosis causes diarrhea, stomach discomfort, fever, headache, nausea, and vomiting | [24,25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arunachalam, K.; Krishnan, G.P.; Sethuraman, S.; Abraham, S.V.P.I.; Krishnan, S.T.; Venkateswar, A.; Arunkumar, J.; Shi, C.; MubarakAli, D. Exploring Possible Ways to Enhance the Potential and Use of Natural Products through Nanotechnology in the Battle against Biofilms of Foodborne Bacterial Pathogens. Pathogens 2023, 12, 270. https://doi.org/10.3390/pathogens12020270
Arunachalam K, Krishnan GP, Sethuraman S, Abraham SVPI, Krishnan ST, Venkateswar A, Arunkumar J, Shi C, MubarakAli D. Exploring Possible Ways to Enhance the Potential and Use of Natural Products through Nanotechnology in the Battle against Biofilms of Foodborne Bacterial Pathogens. Pathogens. 2023; 12(2):270. https://doi.org/10.3390/pathogens12020270
Chicago/Turabian StyleArunachalam, Kannappan, Ganesh Prasath Krishnan, Sathya Sethuraman, Sybiya Vasantha Packiavathy Issac Abraham, Swetha Thirukannamangai Krishnan, Aakanksha Venkateswar, Jagadeesan Arunkumar, Chunlei Shi, and Davoodbasha MubarakAli. 2023. "Exploring Possible Ways to Enhance the Potential and Use of Natural Products through Nanotechnology in the Battle against Biofilms of Foodborne Bacterial Pathogens" Pathogens 12, no. 2: 270. https://doi.org/10.3390/pathogens12020270