

Assessing the Relationship between the Gut Microbiota and Inflammatory Bowel Disease Therapeutics: A Systematic Review
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Study Selection and Eligibility Criteria
2.3. Outcome Assessment
2.4. Data Extraction and Analysis
2.5. Data Synthesis and Statistical Methods
3. Results
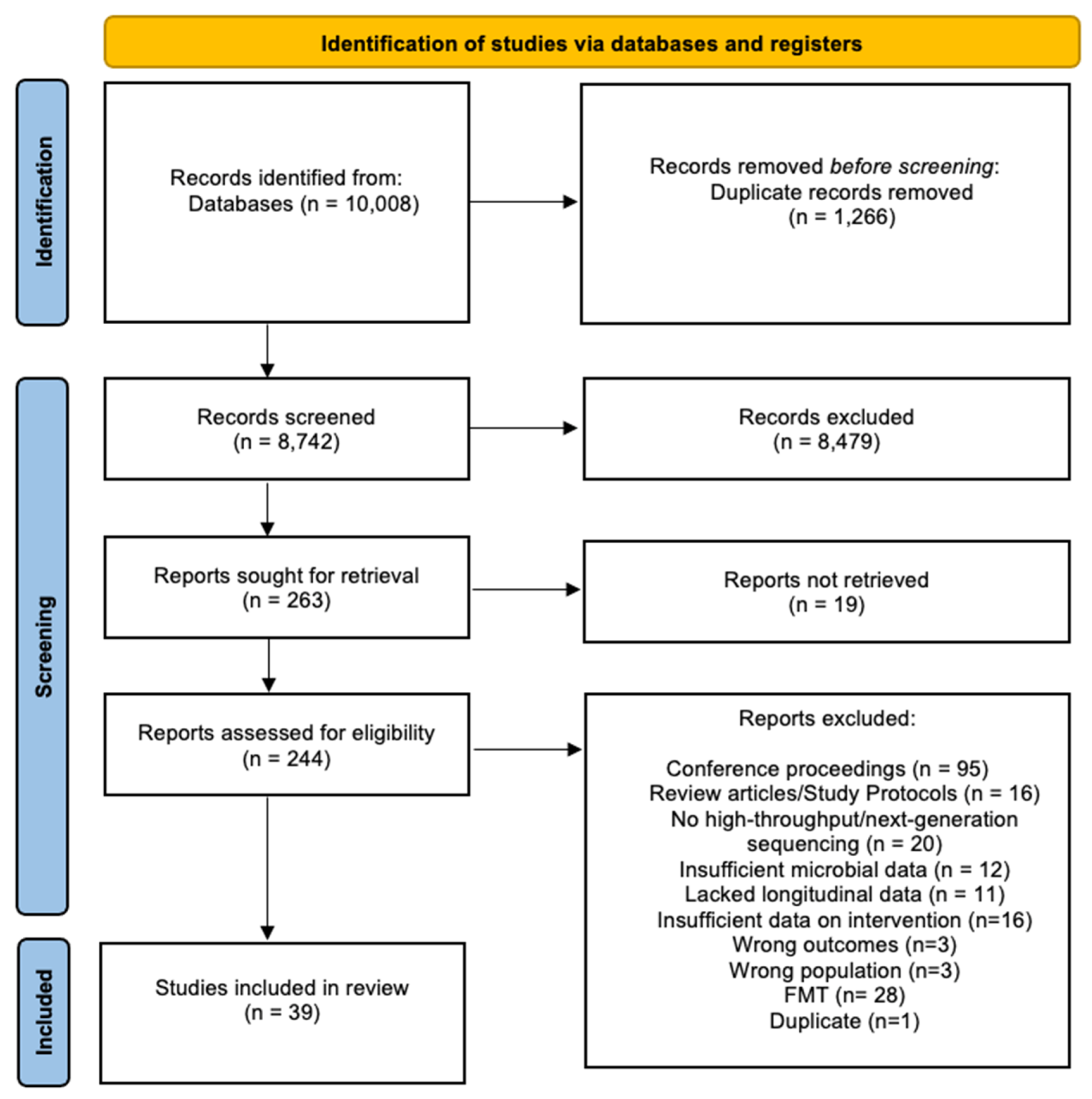
3.1. Search Results
3.2. Description of Included Studies

{kind=link}
{kind=link}
| Disease Subtype | Study | Study Type | Country, Single-(SC) /Multi-Centre (MC) | Diagnosis (# Subjects) | Age Group | Age Group | Duration of Disease (in Years) (Mean SD) | Intervention (# of Subjects); Route, Dose | Length of Intervention (Sampling Points (Weeks)) | Sample Type | Length of Follow-up (Weeks + Sampling Points) | Method of Microbial Composition Analysis | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Age (Years) Mean (SD) | Age (Years) Median/IQR | ||||||||||||
| Biological Therapies | |||||||||||||
| CD CD | Wang et al. [39] | Prospective cohort study (PCS) | China, SC | CD (n = 11) HC (n = 16) | Paediatric | CD: ND (range 4–17) | ND | ND | Infliximab (IFX; n = 11); IV, 5mg/kg | 0, 2, 6 + every 8 weeks | Faeces | ND | 16S rRNA sequencing analysis (16S SA) |
| Doherty et al. [41] | Randomised controlled trial (RCT) | U.S., MC | CD (n = 306) | Adult | CD: 39 (SEM 13) | ND | CD: 12 (SEM 8.8) | Ustekinumab (UST, n = 232); IV, ND (induction therapy (IT)) UST; SC, 270 mg or 90 mg SC (maintenance therapy (MT)) | 8 weeks (ND; IT) 8 weeks (8, 16; MT) | Faeces | 0, 4, 6, 22 | 16S SA | |
| Ribaldone et al. [53] | PCS | Italy, SC | CD (n = 20) | Adult | ND | CD: 52.5 (range 26–69) | CD: 14.5, ND | Adalimumab (ADA, n = 20); ND | 6 months (ND) | Faeces | 0, 26 | Metagenomic (MGS) and 16S SA | |
| Kowalska-Duplaga et al. [54] | PCS | Poland, ND | CD (n = 18) HC (n = 18) | Paediatric | ND | ND | ND | IFX (n = 18); 5 mg/kg, IT | 0, 2, 6 | Faeces | 0, 14 and 6–8 weeks post drug) | 16S SA | |
| Zhuang et al. [40] | PCS | China, SC | CD (n = 49) | Adult | CD: 25 (range 18–48) | ND | CD: 2 (median range 0.5–17) | IFX (n = 49); IV, ND | 0, 2, 6, 14, 22, 30 | Faeces | 0, 6, 30 | 16S SA | |
| Salamon et al. [55] | PCS | Poland, SC | CD (n = 61) HC (n = 17) | Paediatric | CD on IFX: 13.09 (3.76) HC: 11.73 (2.88) | ND | IFX (n = 13); IV 5 mg/kg | 0, 2, 6 | Faeces | 10 weeks (0, 10); 4 weeks after 3rd dose | qPCR analysis | ||
| Wang et al. [38] | PCS | China, MC | CD (n = 18) HC (n = 12) | Paediatric | ND | CD: 12 (11,14) HC: 12 (11,13) | ND | IFX (n = 18); IV 5–10 mg/kg | 0, 2, 6 + every 8 weeks | Faeces | >6 weeks (0, 6 or 30 weeks; after 3rd or 6th dose) | 16S SA | |
| Ventin-Holmberg et al. [56] | PCS | Finland, SC | CD-R (n = 12) CD-NR (n = 18) | Paediatric | ND | CD-R: 13 CD-NR: 14 (ND) | Median CD-R: 1.4 CD-NR: 0.3 | IFX (n = 30), IV, ND | 0, 2, 6 | Faeces | 0, 2, 6 | 16S SA | |
| IBD IBD | Kolho et al. [43] | PCS | Finland, SC | UC (n = 26) CD (n = 36) IBDU (n = 6) HC (n = 26) | Paediatric | ND | IBD: 15.5 (range 9.6–18.3) HC: 13.4 (range 9.7–18.3) | UC, CD, IBDU: 3.5 (median range 0–10.7) | IFX (n = 31); ND ADA (n = 1); ND | ND | Faeces | >6 weeks (0, 2, 6 + 1 during MT) | Phylogenetic microarray and qPCR analysis |
| Ananthakrishnan et al. [42] | PCS | U.S., SC | UC (n = 43) CD (n = 42) | Adult | ND | ND | Remission (n = 31): 9.2 (7.2) No remission (n = 54):15.7 (12.7) | Vedolizumab (VEDO, n = 85); IV, 300mg infusion | 0, 2, 6 + every 8 weeks | Faeces | 0, 6, 14, 30, 54 | MGS | |
| Zhou et al. [57] | PCS | China, SC | UC (n = 51) CD (n = 16) HC (n = 73) | Adult | UC: 41.75 (14.36) CD: 31.81 (12.67) HC: 30.07 (6.36) | ND | ND | IFX (n = 16); 5 mg/kg | 0, 2, 6, 14, 22, 30 | Faeces | 0, 30 | 16S SA | |
| Aden et al. [58] | PCS | Germany, SC | Discovery Cohort (DC): UC (n = 4), CD (n = 8), HC (n = 19) Validation Cohort (VC): UC (n = 13), CD (n = 10), HC (n = 99) | Adult | DC- IBD: 45.4 (4.5) HC: 26.37 (0.5) VC; ND | ND | ND | IFX (n = 10); ND Etanercept (n = 12); ND VEDO (n = 13); ND | ND | Faeces | 0, 2, 6, 30 0, 2, 6, 14 | 16S SA | |
| Olbjorn et al. [50] | PCS | Norway, MC | HC (n = 70) non-IBD (n = 50) CD (n = 80) UC (n = 27) IBDU (n = 3) | Paediatric | ND | ND | ND | Immunomodulators (n = 98); ND Anti-TNF therapy (n =6 4); ND | ND | Faeces | 0, 78 | 16S SA | |
| Dovrolis et al. [59] | PCS | Greece, SC | UC (n = 6) CD (n = 14) HC (n = 9) | Adult | ND | ND | ND | IFX (n = 14); IV, 5 mg/kg | 0, 2, 6 + every 8 weeks | Mucosal Biopsies | 12–20 weeks (0, 12–20) | qPCR and 16S SA | |
| Ding et al. [48] | PCS | ND | CD (n = 76) UC (n = 10) HC (n = 13) | Adult | ND | CD: 38.49 14.61 (19–82) UC: 25.52 17.34 (19–70) HC: 30.5 2.94(27–35) | ND | IFX (n = 66); ND (IT) ADA (n = 10); ND (IT) | ND | Faeces | 16 months (3 monthly) | 16S SA | |
| Schierova et al. [60] | PCS | Czech Republic, SC | CD (n = 34, with 17 at endpoint (EP)) UC (n = 18, with 10 at EP) HC (n=37) | Adult | CD: 35.0 (26.5, 44.0) UC: 31.0 (26.0, 41.3) HC: 36.5 (28.8, 41.3) | ND | CD: 26 (22.5, 35) UC: 27 (21.5, 31) | Anti-TNF therapy (CD (n = 17), ND UC (n = 10), ND) | 38 | Faeces | 0, 2, 8, 14, 20, 26, 32, 38) | 16S SA | |
| Sanchis-Artero et al. [61] | Prospective observational study | Spain, MC | CD (n = 27) HC (n = 16) | Adult | CD: 41.4 (17.4) HC: 29.3 (7.2) | ND | ND | Anti-TNF therapy (n = 27), ND | 26 | Faeces | 0, 13, 26 | 16S SA | |
| Ventin-Holmberg et al. [49] | PCS | Finland, SC | CD (n = 25, with 19 at EP) UC (n = 47 with 33 at EP) | Adult | ND | 31 (24–45) | Median 2 (0–7) | IFX (n = 72, ND, ND) | 52 | Faeces | 0, 2, 6, 12, 52 | 16S SA | |
| Effenberger et al. [62] | Prospective comparative study | Austria, SC | CD (n = 24) UC (n = 12) | Adult | CD: 33.9 (+/−12.9) UC: 44.1 (+/−14.5) | ND | ND | anti-TNF therapy | 30 | Faeces | 0, 12, 30 | 16S SA | |
| Park et al. [63] | PCS | South Korea, MC | CD (n-10) UC (n = 9) HC (n = 19) | Adult | ND | IBD: 33 (23–52) HC: 31 (28–34) | ND | IFX (CD n = 7; UC n = 4) ADA (CD n = 3; UCn = 2) Golimumab (UC n = 3) | 13 | Faeces, saliva, serum, urine | 0, 13 | 16S SA | |
| Non-Biological Therapies | |||||||||||||
| CD | Pigneur et al. [34] | RCT | France, ND | CD (n = 19) | Paediatric | CD on corticosteroids: 13.7 (1.8) HC: ND | ND | ND | Corticosteroids (n = 6); ND, 1 mg/kg/ day (up to max of 60 mg) | 4 daily IT + tapered over 3 months | Faeces | 0, 8 | 16S SA |
| Sprockett et al. [36] | RCT | Europe Canada, Israel, MC | CD (n = 67) | Paediatric | Metronidazole (MET): 13.5 (3.1) Metronidazole + Azithromycin (MET+AZ): 14.2 (3.1) | ND | MET: 0.7 (1) MET+AZ: 1.1 (1.1) | MET; n = 36); ND, 20 mg/kg twice daily (max 1000 mg/day) MET+AZ; n = 31 of which AZ, 7.5 mg/kg once a day (max 500 mg/day) | 8 weeks (daily) + 4 weeks if lacked response | Faeces | 0, 4, 8, 12 | 16S SA | |
| UC | Ishikawa et al. [37] | PCS | Japan, SC | UC (n = 41) | Adult | UC on AFM: 44.7 (14.9) | ND | UC on FMT: 7.0 (8.0) | AFM monotherapy (n = 20); | 2 weeks (daily) | Faeces | 0, 4, 8 | 16S SA |
| Schierova et al. [33] | RCT | Czech Republic, SC | UC (n = 16) | Adult | UC on 5-ASA: 40 (range 31–66) | ND | ND | Mesalazine (n = 8); enema, 4 g | 6 weeks (daily for 2 weeks, every alternate day till week 6) | Faeces | 0, 2, 4, 6, 12 | 16S SA | |
| IBD | Hart et al. [35] | PCS | Canada, SC | UC (n = 10) CD (n = 20) | Paediatric | ND except CS ≤12 years (n = 7) CS ≥13 years (n = 7) | ND | ND | Methyprednisolone (n = 14); IV, 1 mg/kg/day (max 40 mg/day) Transition to oral corticosteroids 1 mg/kg/day PO x 2 weeks followed by progressive wean by 5 mg/wk | 8 weeks (daily) | Faeces | 0, 0.5, 1, 2, 4, 6, 8, 12 | 16S SA |
| Effenberger et al. [62] | Prospective comparative study | Austria, SC | CD (n = 19) UC (n = 10) | Adult | CD: 49.2 (+/- 15.5) UC: 45.0 (+/−10.7) | ND | ND | Azathioprine | 20 | Faeces | 0, 12, 30 | 16S SA | |
| Nutritional therapies | |||||||||||||
| CD | Kaakoush et al. [64] | PCS | Australia SC | CD (n = 5) HC (n = 5) | Paediatric | CD: 9.88 (ND) HC: ND | 9.7 | de novo presenting | EEN (OSMOLITE) | 8–12 | Faecal | 0, 12 weeks post EEN | 16S SA |
| Quince et al. [65] | PCS | Scotland SC | CD (n = 23) HC (n = 21) | Paediatric | CD: ND (range 6.9–14.7) HC: ND (range 4.6–16.9) | NR | NR | EEN (Modulen) | 8 | Faecal | −1, 16, 32, 54 | 16S SA | |
| Tang et al. [66] | PCS | China | CD (n = 31) HC (n = 12) | Paediatric | HC n = 12; 10.45 ± 2.39; CD RE n = 17; 12.02 ± 1.88; CD NRE n = 8; 9.84 ± 4.51 | NR | de novo presenting | EEN (oral) | 8 | Faecal | 8 weeks | 16S SA | |
| Dunn et al. [67] | PCS | Canada | CD (n = 10) HC (n = 5) | Paediatric | CD: 12.27 | CD: 12 | mainly de novo presenting | EEN; (nasogastric/gastric) | 12 | Faecal | 12 weeks | 16S SA | |
| Costa-Santos et al. [68] | PCS | Portugal | CD patients receive pre-operative EEN = 10; CD patients receive immediate surgery = 5 | Adult | CD: 45.4 ± 19.1 | NR | 9 years (0–33 years) | Pre-operative EEN (oral) | 15–70 days | Faecal | 24 weeks | 16S SA | |
| Diederen et al. [69] | PCS | Netherlands | CD (n = 27) HC (n = 18) | Paediatric | NR | CD: 14 (12–15) HC: 13 (11–16) | de novo presenting | EEN + concomitant thiopurines | 6 | Faecal | 6 weeks | 16S SA | |
| Tang et al. [66] | Prospective comparative study | China, SC | CD (n = 31) HC (n = 12) | Paediatric | CD:11.3 (3.5) HC:10.5 (2.4) | ND | ND | EEN (n = 31); oral, ND | 0, 2, 8 | Faeces | 0, 2, 6, 8 | 16S SA | |
| Jiang et al. [70] | Prospective study | China, SC | CD (n = 7) | Adult | 27.9 (12.4) | ND | ND | Enteral nutritional powder (n = 7); oral, 250 mL | 0, 8 | Faeces | 0, 8 | 16S SA | |
| Suskind et al. [71] | RCT | U.S. | CD (n = 10) | Paediatric | 14.3 ±2.9 (7–18) | NR | NR | Specific carbohydrate diet (SCD) | 12 | Faecal | NR | 16S SA | |
| UC | Wilson et al. [72] | Prospective study | U.K., SC | UC (n = 17) | Adult | 35 (10) | ND | ND | GOS supplement (n = 17); oral, 2.8 g | 0, 6 | Faeces | 0, 6 | 16S SA |
| Chen et al. [73] | Prospective comparative study | China, SC | UC (n = 14) HC (n = 14) | Adult | UC and HC: 42.4 (15.0) | ND | ND | GMA (n = 28); intravenous, ND | 5 “sessions”; (baseline and endpoint) (n = 5) 10 “sessions”; (baseline and endpoint) (n = 8) | Faeces | baseline and endpoint | 16S SA | |
| Valcheva et al. [51] | PCS | Canada | UC (n = 25) Inulin 7.5g/d n = 12. Inulin 15g/d n = 13 | Adult | Inulin 7.5g/d = 36 Inulin 15g/d = 39 | NR | NR | Prebiotic (inulin) | 9 | Faecal | NR | 16S SA | |
| IBD | Suskind et al. [52] | Observational study | U.S., MC | CD = 9, UC = 3 | Paediatric | 12.8 ± 2.2.(10–17) | NR | 1.3 ± 1.6 years (0–5) | Specific carbohydrate diet (SCD) | 12 | Faecal | NR | 16S SA |
| Olendzki et al. [74] | Prospective comparative study | U.S., SC | CD (n = 14) UC (n = 7) | Adult | 40.5 (12.8) | ND | ND | Dietary intervention (n = 25); oral | 8 | Faeces | 0, 8 | MGS | |
3.3. Impact of IBD Treatment on the Gut Microbiota
3.3.1. Biologics
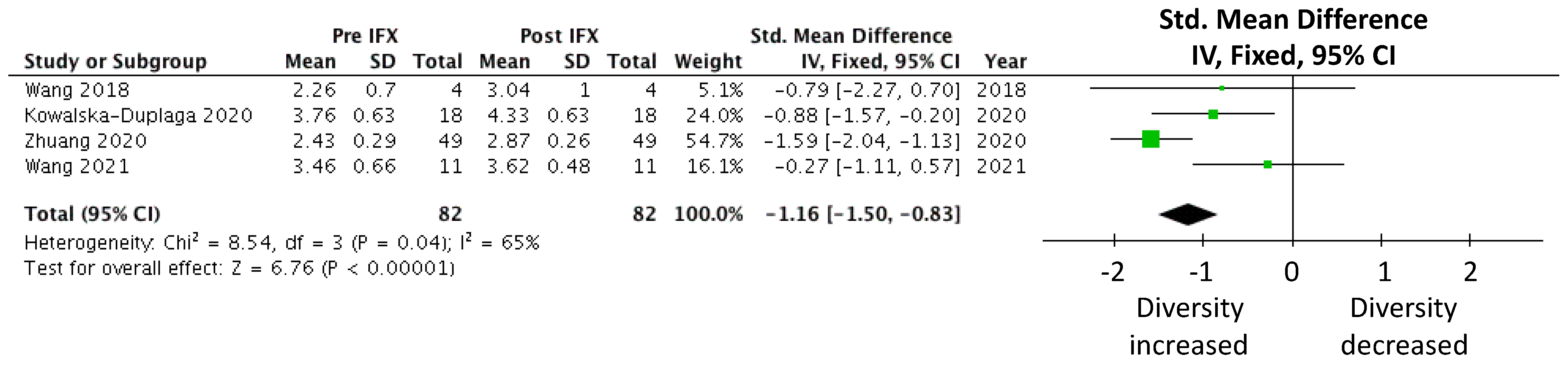
Microbial Diversity
| Disease Subtype | Reference/No. of Subjects | Alpha Diversity Changes | Pre- Treatment (Week) | Post-Treatment (Week) | |||
|---|---|---|---|---|---|---|---|
| Biological therapies | |||||||
| CD | Wang et al. [38] CD (n = 4) HC (n = 16) | Week | 0 * | 14 * | 22 * | 30 * | 38 * |
| IFX Shannon#○ | 2.26 | 3.04 | 3.63 | 3.27 | 2.99 | ||
| IFX Observed Species#◊ | 148 | 219 | 321.5 | 281 | 202 | ||
| IFX Simpson#○ | 0.824 | 0.912 | 0.940 | 0.918 | 0.907 | ||
| IFX Inverse Simpson#○ | 5.68 | 11.4 | 18.3 | 12.1 | 10.8 | ||
| HC Shannon#○ | 3.49 | ||||||
| HC Observed Species#◊ | 393 | ||||||
| HC Simpson#○ | 0.898 | ||||||
| HC Inverse Simpson#○ | 10.00 | ||||||
| Kowalska-Duplaga et al. [54] CD (n = 18) HC (n = 18) | Week | 0 | 14 | ||||
| IFX Shannon#○ | 3.762 | 4.326 | |||||
| IFX Observed Species#◊ | 46.5 | 64 | |||||
| IFX Faith PD#□ | 6.5723 | 7.65354 | |||||
| IFX Pielou#○ | 0.70198 | 0.6995 | |||||
| HC Shannon#○ | 4.664 | ||||||
| HC Observed Species#◊ | 64 | ||||||
| HC Faith PD#□ | 8.1631 | ||||||
| HC Pielou#○ | 0.7739 | ||||||
| Zhuang et al. [40] CD (n = 49) | Week | 0 | 6 | 30 | |||
| IFX Shannon#○ | 2.43 | 2.61 | 2.87 | ||||
| IFX Observed Species#◊ | 113 | 145 | 166.5 | ||||
| IFX Chao1#◊ | 148.1 | 183.1 | 209.7 | ||||
| IFX Good’s Coverage#◊ | 0.999 | 0.999 | 0.999 | ||||
| Wang et al. [39] CD (n = 11) HC (n = 12) | Week | 0 | T1^ | ||||
| IFX Shannon#○ | 3.46 | 3.62 | |||||
| IFX Chao1#◊ | 489.97 | 506.2 | |||||
| IFX Simpson#○ | 0.891 | 0.924 | |||||
| IFX Inverse Simpson#○ | 9.19 | 13.17 | |||||
| HC Shannon○ | 3.7 | - | |||||
| HC Chao1◊ | 475 | - | |||||
| HC Simpson○ | 0.94 | - | |||||
| HC Inverse Simpson○ | 14.5 | - | |||||
| Zhou et al. [57] CD R (n = 9) CD Relapse (n = 7) HC (n = 11) | Week | 0 | 30 | ||||
| IFX R Shannon○ | 5.5 | 6.25 | |||||
| IFX R PD□ | 15 | 19 | |||||
| IFX Relapse Shannon○ | 5.5 | 5.6 | |||||
| IFX Relapse PD□ | 15 | 16 | |||||
| HC Shannon○ | 5.6 | - | |||||
| HC PD□ | 16 | - | |||||
| Doherty et al. [41] CD R (n = 18) CD NR (n = 30) | Week | 0 | 4 | 6 | 22 | ||
| UST R Inverse Simpson#○ | 6.65 | 9.44 | 8.42 | 10.7 | |||
| UST NR Inverse Simpson○ | 5.25 | 5.0 | 5.5 | 6.5 | |||
| Dovrolis et al. [59] CD R (n = 5) CD NR (n = 5) | Week | 0 | 12–20 | ||||
| CD IFX R Pielou○ | 0.74 | 0.73 | |||||
| CD IFX NR Pielou○ | 0.74 | 0.725 | |||||
| Sanchis-Artero et al. [61] CD R (n = 13) CD NR (n = 14) HC (n = 16) | Week | 0 | 24 | ||||
| CD R anti-TNF Shannon○ | 5.65 | 6.15 | |||||
| CD NR anti-TNF Shannon○ | 5.65 | 5.2 | |||||
| CD R anti-TNF Chao1◊ | 92.5 | 112.5 | |||||
| CD NR anti-TNF Chao1◊ | 92.5 | 68.8 | |||||
| HC Shannon○ | 6.33 | ND | |||||
| HC Chao1◊ | 116.3 | ND | |||||
| UC | Dovrolis et al. [59] UC R (n = 2) UC NR (n = 2) | Week | 0 | 12–20 | |||
| UC IFX R Pielou○ | 0.74 | 0.69 | |||||
| UC IFX NR Pielou○ | 0.74 | 0.78 | |||||
| IBD | Kolho et al. [43] IBD R (n = 6) IBD NR (n = 5) | Week | 0 | 2 | 6 | ||
| IFX/ADA Responder Inverse Simpson○ | 173 | 188 | 175 | ||||
| IFX/ADA Non-Responder Inverse Simpson○ | 133 | 113 | 128 | ||||
| Aden et al. [58] IBD (n = 12) HC (n = 19) | Week | 0 | 2 | 6 | 30 | ||
| Etanercept Shannon○ | 2.4 | 2.5 | 2.7 | 2.7 | |||
| Etanercept Observed Species◊ | 64 | 64 | 61 | 70 | |||
| Etanercept Chao1◊ | 75 | 76 | 80 | 86 | |||
| Etanercept PD□ | 6 | 6.5 | 6 | 7.5 | |||
| HC Shannon○ | 3.2 | ||||||
| HC Observed Species◊ | 72 | ||||||
| HC Chao1◊ | 86 | ||||||
| HC PD□ | 7.5 | ||||||
| Schierova et al. [60] CD (n = 26) UC (n = 18) HC (n = 37) | Week | 0 | 20 | 38 | |||
| CD Anti-TNF Shannon○ | 5.65 | 5.7 | 5.9 | ||||
| UC Anti-TNF Shannon○ | 5.2 | 5.85 | 5.85 | ||||
| HC Shannon○ | 6.15 | ND | ND | ||||
| Park et al. [63] IBD (n = 19) HC (n = 19) | Week | 0 | 12 | ||||
| IBD Anti-TNF Shannon○ | 3.28 | 3.35 | |||||
| IBD Anti-TNF Simpson○ | 0.068 | 0.068 | |||||
| IBD Anti-TNF Chao1◊ | 290 | 315 | |||||
| IBD Anti-TNF ACE◊ | 305 | 325 | |||||
| IBD Anti-TNF Jackknife | 295 | 330 | |||||
| IBD Anti-TNF NPShannon○ | 3.29 | 3.36 | |||||
| HC Shannon○ | 3.73 | ND | |||||
| HC Simpson○ | 0.05 | ND | |||||
| HC Chao1◊ | 465 | ND | |||||
| HC ACE◊ | 485 | ND | |||||
| HC Jackknife | 503 | ND | |||||
| HC NPShannon○ | 3.73 | ND | |||||
| Ventin-Holmberg et al. [49] CD R (n = 13) CD PR (n = 4) CD NR (n = 6) UC R (n = 31) UC PR (n = 8) UC NR (n = 8) | Week | 0 | 2 | 6 | 12 | 52 | |
| CD R IFX Simpson○ | 9 | 9 | 12 | 8.75 | 10.5 | ||
| CD NR IFX Simpson○ | 7 | 8.5 | 8.3 | 9 | ND | ||
| CD PR IFX Simpson○ | 5.5 | 5.5 | 10.3 | 8 | 8 | ||
| CD R IFX OTU◊ | 97.5 | 105 | 107.5 | 107.5 | 110 | ||
| CD NR IFX OTU◊ | 90 | 95 | 97.5 | 95 | ND | ||
| CD PR IFX OTU◊ | 55 | 77.5 | 107.5 | 95 | 107.5 | ||
| UC R IFX Simpson○ | 11.25 | 11 | 10.5 | 10.5 | 11 | ||
| UC NR IFX Simpson○ | 7 | 10.75 | 8 | 10.25 | 4.75 | ||
| UC PR IFX Simpson○ | 9.75 | 8.5 | 10 | 10 | 10.5 | ||
| UC R IFX OTU◊ | 105 | 115 | 107.5 | 105 | 105 | ||
| UC NR IFX OTU◊ | 90 | 105 | 97.5 | 105 | 102.5 | ||
| UC PR IFX OTU◊ | 107.5 | 112.5 | 110 | 105 | 105 | ||
| Ventin-Holmberg et al. [56] IBD R (n = 10) IBD NR (n-16) | Week | 0 | 2 | 6 | |||
| IBD R IFX Simpson○ | 8.5 | 7 | 6.5 | ||||
| IBD NR IFX Inverse Simpson○ | 6 | 6.5 | 7 | ||||
| IBD R IFX OTU◊ | 75 | 72.5 | 72.5 | ||||
| IBD NR IFX OTU◊ | 65 | 66 | 70 | ||||
| Effenberger et al. [62] IBD R (n = 18) IBD NR (n = 18) | Week | 0 | 98 days | ||||
| R Anti-TNF Shannon○ | 0.4 | 0.2 | |||||
| NR Anti-TNF Shannon○ | −0.2 | 0.175 | |||||
| Non-biological therapies | |||||||
| CD | Sprockett et al. [36] ^1 MET (n = 36) MET+AZ (n = 31) MET/MET+AZ (n = 11) | Week | 0 | 4 | 8 | 12 | |
| CD MET Faith’s PD□ | 18.5 | 11.5 | 12 | 16 | |||
| CD MET + AZ Faith’s PD□ | 17.5 | 11 | 10 | 13 | |||
| CD MET/MET+AZ Faith’s PD□ | 18.5 | 15.5 | 6.5 | 9.5 | |||
| UC | Schierova et al. [33] UC R (n = 4) UC NR (n = 4) | Week | 0 | 2 | |||
| UC R 5-ASA Shannon○ | 6.6 | 6.9 | |||||
| UC NR 5-ASA Shannon○ | 6.65 | 6.85 | |||||
| IBD | Hart et al. [35] ^2 UC (n = 10) CD (n = 4) | Week | 0 | 12 | |||
| Corticosteroids Shannon○ | 2.85 | 3.6 | |||||
| Effenberger et al. [62] IBD R (n = 15) IBD NR (n = 14) | Week | 0 | 14 | ||||
| R AZA Shannon○ | −0.2 | −0.2 | |||||
| NR AZA Shannon○ | −0.05 | −0.85 | |||||
| Nutritional therapies | |||||||
| CD | Kaakoush et al. [64] CD (n = 5) HC (n = 5) | Week | 0 | 8 | 26 | ||
| EEN CD Shannon#○ | 2.25 ± 0.24 | ND | ND | ||||
| HC Shannon#○ | 2.75 ± 0.14 | ND | ND | ||||
| Quince et al. [65] CD (n = 23) HC (n = 21) | Week | 0 | 2 | 4 | 8 | Free diet | |
| EEN CD Shannon#○ | 2.88 ± 0.597 | 2.82 ± 0.492 | 2.49 ± 0.731 | 2.84 ± 0.513 | 3.04 ± 0.686 | ||
| HC Shannon○ | ND | ||||||
| Tang et al. [66] HC (n = 12) CD Remission (n = 17) CD Non-remission (n = 8) | Week | 0 | 8 | ||||
| EEN CD-R Shannon#○ | 1.84 ± 0.16 | 2.45 ± 0.1 | |||||
| EEN CD-NR Shannon#○ | 1.33 ± 0.23 | 1.76 ± 0.23 | |||||
| HC Shannon#○ | 3.1 ± 0.2 | ND | |||||
| Dunn et al. [67] CD (n = 10 at baseline, n = 9 at 12 weeks) HC (n = 5) | Week | 0 | 12 | ||||
| EEN CD Sustained Remission EEN Chao | ~1300 | ~1250 | |||||
| EEN CD Non-Sustained Remission Chao◊ | ~1000 | ~ 700 | |||||
| HC Chao◊ | ~2000 | ND | |||||
| Costa-Santos et al. [68] CD (n = 15) | Week | 0 | 8 | ||||
| CD Faith’s PD#□ | 8 ± 2.3 | 5.2 ± 1.5 | |||||
| Diederen et al. [69] CD (n = 27) HC (n = 18) | Week | 0 | 1–5 | 6 | 9 | ||
| EEN CD Inv Simpsons#○ | 19.40 | 11.57 | 12.85 | 16.87 | |||
| HC Inv Simpsons#○ | 20.37 | ||||||
| Suskind et al. [52] CD (n = 8) | Week | 0 | 12 | ||||
| SCD CD Shannon#○ | 2.62(2.4–3.1) | 2.7(2.3–3.1) | |||||
| Suskind et al. [71] CD (n = 5) | Week | 0 | 2 | 12 | |||
| SCD CD Inverse Simpson#○ | 103 (89–132) | 115 (95–130) | 112 (107–147) | ||||
| Tang et al. [66] CD (n = 31) HC (n = 12) | Week | 0 | 2 | 8 | |||
| HC Shannon#○ | 3.1 | ND | ND | ||||
| EEN NR CD Shannon#○ | 1.33 | ND | 1.76 | ||||
| EEN R CD Shannon#○ | 1.84 | ND | 2.45 | ||||
| Observed Species#◊ | 172 | ND | ND | ||||
| Jiang et al. [70] CD (n = 7) | Week | 0 | 8 | ||||
| Shannon#○ | ND | ||||||
| Observed Species#◊ | 292 | 390 | |||||
| UC | Wilson et al. [72] UC (n = 17) | Week | 0 | 6 | |||
| Shannon#○ | 2.5 | 2.2 | |||||
| Chao1#◊ | 50.4 | 35.6 | |||||
| Observed Species#◊ | 178 | ND | |||||
| Chen et al. [73] UC (n = 14) HC (n = 14) | “Sessions” | 0 | 5 | 10 | |||
| UC Shannon#○ | ND | ||||||
| HC Shannon#○ | ND | ||||||
| UC Observed Species#◊ | 523 | 751 | 521 | ||||
| IBD | Olendzki et al. [74] CD (n = 14) UC (n = 7) | Week | 0 | 6 | |||
| Shannon#○ | CD = 2.1 UC = 2.3 | ||||||
Microbial Composition Changes
Microbial Predictors of Treatment Outcomes
3.3.2. Non-Biological Therapies
Microbial Diversity
Microbial Composition
3.3.3. Nutritional Therapies
Microbial Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McQuaid, K.R. Inflammatory Bowel Disease. In Current Medical Diagnosis & Treatment 2022; Papadakis, M.A., McPhee, S.J., Rabow, M.W., McQuaid, K.R., Eds.; McGraw-Hill Education: New York, NY, USA, 2022. [Google Scholar]
- Alatab, S.; Sepanlou, S.G.; Ikuta, K.; Vahedi, H.; Bisignano, C.; Safiri, S.; Sadeghi, A. The global, regional, and national burden of inflammatory bowel disease in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 17–30. [Google Scholar] [CrossRef]
- Lichtenstein, G.R.; Shahabi, A.; Seabury, S.A.; Lakdawalla, D.N.; Espinosa, O.D.; Green, S.; Brauer, M.; Baldassano, R.N. Lifetime Economic Burden of Crohn’s Disease and Ulcerative Colitis by Age at Diagnosis. Clin. Gastroenterol. Hepatol. 2020, 18, 889–897.e810. [Google Scholar] [CrossRef]
- Peyrin-Biroulet, L.; Sandborn, W.; Sands, B.E.; Reinisch, W.; Bemelman, W.; Bryant, R.V.; D’Haens, G.; Dotan, I.; Dubinsky, M.; Feagan, B.; et al. Selecting Therapeutic Targets in Inflammatory Bowel Disease (STRIDE): Determining Therapeutic Goals for Treat-to-Target. Am. J. Gastroenterol. 2015, 110, 1324–1338. [Google Scholar] [CrossRef]
- Privitera, G.; Pugliese, D.; Lopetuso, L.R.; Scaldaferri, F.; Neri, M.; Guidi, L.; Gasbarrini, A.; Armuzzi, A. Novel trends with biologics in inflammatory bowel disease: Sequential and combined approaches. Ther. Adv. Gastroenterol. 2021, 14, 17562848211006669. [Google Scholar] [CrossRef]
- Colombel, J.F.; Narula, N.; Peyrin-Biroulet, L. Management Strategies to Improve Outcomes of Patients with Inflammatory Bowel Diseases. Gastroenterology 2017, 152, 351–361.e355. [Google Scholar] [CrossRef]
- Berg, D.R.; Colombel, J.F.; Ungaro, R. The Role of Early Biologic Therapy in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2019, 25, 1896–1905. [Google Scholar] [CrossRef]
- Khanna, R.; Bressler, B.; Levesque, B.G.; Zou, G.; Stitt, L.W.; Greenberg, G.R.; Panaccione, R.; Bitton, A.; Paré, P.; Vermeire, S.; et al. Early combined immunosuppression for the management of Crohn’s disease (REACT): A cluster randomised controlled trial. Lancet 2015, 386, 1825–1834. [Google Scholar] [CrossRef]
- de Jong, M.E.; Smits, L.J.T.; van Ruijven, B.; den Broeder, N.; Russel, M.; Römkens, T.E.H.; West, R.L.; Jansen, J.M.; Hoentjen, F. Increased Discontinuation Rates of Anti-TNF Therapy in Elderly Inflammatory Bowel Disease Patients. J. Crohns Colitis 2020, 14, 888–895. [Google Scholar] [CrossRef]
- Qiu, Y.; Chen, B.L.; Mao, R.; Zhang, S.H.; He, Y.; Zeng, Z.R.; Ben-Horin, S.; Chen, M.H. Systematic review with meta-analysis: Loss of response and requirement of anti-TNFα dose intensification in Crohn’s disease. J. Gastroenterol. 2017, 52, 535–554. [Google Scholar] [CrossRef]
- Ben-Horin, S.; Chowers, Y. Tailoring anti-TNF therapy in IBD: Drug levels and disease activity. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 243–255. [Google Scholar] [CrossRef]
- Hold, G.L.; Smith, M.; Grange, C.; Watt, E.R.; El-Omar, E.M.; Mukhopadhya, I. Role of the gut microbiota in inflammatory bowel disease pathogenesis: What have we learnt in the past 10 years? World J. Gastroenterol. 2014, 20, 1192–1210. [Google Scholar] [CrossRef]
- Yue, B.; Luo, X.; Yu, Z.; Mani, S.; Wang, Z.; Dou, W. Inflammatory Bowel Disease: A Potential Result from the Collusion between Gut Microbiota and Mucosal Immune System. Microorganisms 2019, 7, 440. [Google Scholar] [CrossRef]
- Yamashiro, Y. Gut Microbiota in Health and Disease. Ann. Nutr. Metab. 2017, 71, 242–246. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A microbial signature for Crohn’s disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef]
- Halfvarson, J.; Brislawn, C.J.; Lamendella, R.; Vázquez-Baeza, Y.; Walters, W.A.; Bramer, L.M.; D’Amato, M.; Bonfiglio, F.; McDonald, D.; Gonzalez, A.; et al. Dynamics of the human gut microbiome in inflammatory bowel disease. Nat. Microbiol. 2017, 2, 17004. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef]
- Sokol, H.; Seksik, P. The intestinal microbiota in inflammatory bowel diseases: Time to connect with the host. Curr. Opin. Gastroenterol. 2010, 26, 327–331. [Google Scholar] [CrossRef]
- Joossens, M.; Huys, G.; Cnockaert, M.; De Preter, V.; Verbeke, K.; Rutgeerts, P.; Vandamme, P.; Vermeire, S. Dysbiosis of the faecal microbiota in patients with Crohn’s disease and their unaffected relatives. Gut 2011, 60, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Seksik, P.; Furet, J.P.; Firmesse, O.; Nion-Larmurier, I.; Beaugerie, L.; Cosnes, J.; Corthier, G.; Marteau, P.; Doré, J. Low counts of Faecalibacterium prausnitzii in colitis microbiota. Inflamm. Bowel Dis. 2009, 15, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych, I.; Castro Sangrador, J.; Dordević, D.; Rozehnalová, M.; Černý, M.; Fafula, R.; Vítězová, M.; Rittmann, S.K.R. Evaluation of Physiological Parameters of Intestinal Sulfate-Reducing Bacteria Isolated from Patients Suffering from IBD and Healthy People. J. Clin. Med. 2020, 9, 1920. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych, I.; Dordević, D.; Vítězová, M. Possible synergy effect of hydrogen sulfide and acetate produced by sulfate-reducing bacteria on inflammatory bowel disease development. J. Adv. Res. 2021, 27, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Britton, G.J.; Contijoch, E.J.; Mogno, I.; Vennaro, O.H.; Llewellyn, S.R.; Ng, R.; Li, Z.; Mortha, A.; Merad, M.; Das, A.; et al. Microbiotas from Humans with Inflammatory Bowel Disease Alter the Balance of Gut Th17 and RORγt(+) Regulatory T Cells and Exacerbate Colitis in Mice. Immunity 2019, 50, 212–224.e214. [Google Scholar] [CrossRef]
- Sokol, H.; Landman, C.; Seksik, P.; Berard, L.; Montil, M.; Nion-Larmurier, I.; Bourrier, A.; Le Gall, G.; Lalande, V.; De Rougemont, A.; et al. Fecal microbiota transplantation to maintain remission in Crohn’s disease: A pilot randomized controlled study. Microbiome 2020, 8, 12. [Google Scholar] [CrossRef]
- Haiser, H.J.; Seim, K.L.; Balskus, E.P.; Turnbaugh, P.J. Mechanistic insight into digoxin inactivation by Eggerthella lenta augments our understanding of its pharmacokinetics. Gut Microbes 2014, 5, 233–238. [Google Scholar] [CrossRef]
- Rekdal, V.M.; Bess, E.N.; Bisanz, J.E.; Turnbaugh, P.J.; Balskus, E.P. Discovery and inhibition of an interspecies gut bacterial pathway for Levodopa metabolism. Science 2019, 364, eaau6323. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Weiner, B.; Ford, C.B.; Sellman, B.R.; Hammond, S.A.; Freeman, D.J.; Dennis, P.; Soria, J.C.; Wortman, J.R.; Henn, M.R. Intervention strategies for microbial therapeutics in cancer immunotherapy. Immunooncol. Technol. 2020, 6, 9–17. [Google Scholar] [CrossRef]
- Chaput, N.; Lepage, P.; Coutzac, C.; Soularue, E.; Le Roux, K.; Monot, C.; Boselli, L.; Routier, E.; Cassard, L.; Collins, M.; et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 2017, 28, 1368–1379. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Schierová, D.; Březina, J.; Mrázek, J.; Fliegerová, K.O.; Kvasnová, S.; Bajer, L.; Drastich, P. Gut Microbiome Changes in Patients with Active Left-Sided Ulcerative Colitis after Fecal Microbiome Transplantation and Topical 5-aminosalicylic Acid Therapy. Cells 2020, 9, 2283. [Google Scholar] [CrossRef] [PubMed]
- Pigneur, B.; Lepage, P.; Mondot, S.; Schmitz, J.; Goulet, O.; Doré, J.; Ruemmele, F.M. Mucosal Healing and Bacterial Composition in Response to Enteral Nutrition Vs Steroid-based Induction Therapy-A Randomised Prospective Clinical Trial in Children With Crohn’s Disease. J. Crohns Colitis 2019, 13, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Hart, L.; Farbod, Y.; Szamosi, J.C.; Yamamoto, M.; Britz-McKibbin, P.; Halgren, C.; Zachos, M.; Pai, N. Effect of Exclusive Enteral Nutrition and Corticosteroid Induction Therapy on the Gut Microbiota of Pediatric Patients with Inflammatory Bowel Disease. Nutrients 2020, 12, 1691. [Google Scholar] [CrossRef] [PubMed]
- Sprockett, D.; Fischer, N.; Boneh, R.S.; Turner, D.; Kierkus, J.; Sladek, M.; Escher, J.C.; Wine, E.; Yerushalmi, B.; Dias, J.A.; et al. Treatment-Specific Composition of the Gut Microbiota Is Associated with Disease Remission in a Pediatric Crohn’s Disease Cohort. Inflamm. Bowel Dis. 2019, 25, 1927–1938. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, D.; Sasaki, T.; Osada, T.; Kuwahara-Arai, K.; Haga, K.; Shibuya, T.; Hiramatsu, K.; Watanabe, S. Changes in Intestinal Microbiota Following Combination Therapy with Fecal Microbial Transplantation and Antibiotics for Ulcerative Colitis. Inflamm. Bowel Dis. 2017, 23, 116–125. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, X.; Ghozlane, A.; Hu, H.; Li, X.; Xiao, Y.; Li, D.; Yu, G.; Zhang, T. Characteristics of Faecal Microbiota in Paediatric Crohn’s Disease and Their Dynamic Changes During Infliximab Therapy. J. Crohns Colitis 2018, 12, 337–346. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, X.; Zhang, X.; Xiao, F.; Hu, H.; Li, X.; Dong, F.; Sun, M.; Xiao, Y.; Ge, T.; et al. Microbial and metabolic features associated with outcome of infliximab therapy in pediatric Crohn’s disease. Gut Microbes 2021, 13, 1865708. [Google Scholar] [CrossRef]
- Zhuang, X.; Tian, Z.; Feng, R.; Li, M.; Li, T.; Zhou, G.; Qiu, Y.; Chen, B.; He, Y.; Chen, M.; et al. Fecal Microbiota Alterations Associated with Clinical and Endoscopic Response to Infliximab Therapy in Crohn’s Disease. Inflamm. Bowel Dis. 2020, 26, 1636–1647. [Google Scholar] [CrossRef]
- Doherty, M.K.; Ding, T.; Koumpouras, C.; Telesco, S.E.; Monast, C.; Das, A.; Brodmerkel, C.; Schloss, P.D. Fecal Microbiota Signatures Are Associated with Response to Ustekinumab Therapy among Crohn’s Disease Patients. mBio 2018, 9, e02120-17. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Luo, C.; Yajnik, V.; Khalili, H.; Garber, J.J.; Stevens, B.W.; Cleland, T.; Xavier, R.J. Gut Microbiome Function Predicts Response to Anti-integrin Biologic Therapy in Inflammatory Bowel Diseases. Cell Host Microbe 2017, 21, 603–610.e603. [Google Scholar] [CrossRef] [PubMed]
- Kolho, K.L.; Korpela, K.; Jaakkola, T.; Pichai, M.V.; Zoetendal, E.G.; Salonen, A.; de Vos, W.M. Fecal Microbiota in Pediatric Inflammatory Bowel Disease and Its Relation to Inflammation. Am. J. Gastroenterol. 2015, 110, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Wells, G.A.; Shea, B.; O'Connell, D.; Peterson, J.; Welcon, V.; Losos, M.; Tugwell, P. The Newcastle-Ottawa Scale (NOS) for assessing the quality of nonrandomised studies in meta-analyses. Available online: https://www.ohri.ca/programs/clinical_epidemiology/oxford.asp (accessed on 19 October 2022).
- Higgins, J.P.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Sutton, A.J.; Abrams, K.R.; Jones, D.R.; Sheldon, T.A.; Song, F. Methods for Meta-Analysis in Medical Research; John Wiley & Sons, Ltd.: New York, NY, USA, 2000. [Google Scholar]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef]
- Ding, N.S.; McDonald, J.A.K.; Perdones-Montero, A.; Rees, D.N.; Adegbola, S.O.; Misra, R.; Hendy, P.; Penez, L.; Marchesi, J.R.; Holmes, E.; et al. Metabonomics and the Gut Microbiome Associated with Primary Response to Anti-TNF Therapy in Crohn’s Disease. J. Crohns Colitis 2020, 14, 1090–1102. [Google Scholar] [CrossRef] [PubMed]
- Ventin-Holmberg, R.; Eberl, A.; Saqib, S.; Korpela, K.; Virtanen, S.; Sipponen, T.; Salonen, A.; Saavalainen, P.; Nissilä, E. Bacterial and Fungal Profiles as Markers of Infliximab Drug Response in Inflammatory Bowel Disease. J. Crohns Colitis 2021, 15, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Olbjørn, C.; Cvancarova Småstuen, M.; Thiis-Evensen, E.; Nakstad, B.; Vatn, M.H.; Jahnsen, J.; Ricanek, P.; Vatn, S.; Moen, A.E.F.; Tannæs, T.M.; et al. Fecal microbiota profiles in treatment-naïve pediatric inflammatory bowel disease–associations with disease phenotype, treatment, and outcome. Clin. Exp. Gastroenterol. 2019, 12, 37–49. [Google Scholar] [CrossRef]
- Valcheva, R.; Koleva, P.; Martínez, I.; Walter, J.; Gänzle, M.G.; Dieleman, L.A. Inulin-type fructans improve active ulcerative colitis associated with microbiota changes and increased short-chain fatty acids levels. Gut Microbes 2019, 10, 334–357. [Google Scholar] [CrossRef]
- Suskind, D.L.; Cohen, S.A.; Brittnacher, M.J.; Wahbeh, G.; Lee, D.; Shaffer, M.L.; Braly, K.; Hayden, H.S.; Klein, J.; Gold, B.; et al. Clinical and Fecal Microbial Changes With Diet Therapy in Active Inflammatory Bowel Disease. J. Clin. Gastroenterol. 2018, 52, 155–163. [Google Scholar] [CrossRef]
- Ribaldone, D.G.; Caviglia, G.P.; Abdulle, A.; Pellicano, R.; Ditto, M.C.; Morino, M.; Fusaro, E.; Saracco, G.M.; Bugianesi, E.; Astegiano, M. Adalimumab Therapy Improves Intestinal Dysbiosis in Crohn’s Disease. J. Clin. Med. 2019, 8, 1646. [Google Scholar] [CrossRef]
- Kowalska-Duplaga, K.; Kapusta, P.; Gosiewski, T.; Sroka-Oleksiak, A.; Ludwig-Słomczyńska, A.H.; Wołkow, P.P.; Fyderek, K. Changes in the Intestinal Microbiota Are Seen Following Treatment with Infliximab in Children with Crohn’s Disease. J. Clin. Med. 2020, 9, 687. [Google Scholar] [CrossRef] [PubMed]
- Salamon, D.; Gosiewski, T.; Krawczyk, A.; Sroka-Oleksiak, A.; Duplaga, M.; Fyderek, K.; Kowalska-Duplaga, K. Quantitative changes in selected bacteria in the stool during the treatment of Crohn’s disease. Adv. Med. Sci. 2020, 65, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Ventin-Holmberg, R.; Höyhtyä, M.; Saqib, S.; Korpela, K.; Nikkonen, A.; Salonen, A.; de Vos, W.M.; Kolho, K.L. The gut fungal and bacterial microbiota in pediatric patients with inflammatory bowel disease introduced to treatment with anti-tumor necrosis factor-α. Sci. Rep. 2022, 12, 6654. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, Z.Z.; He, Y.; Yang, Y.; Liu, L.; Lin, Q.; Nie, Y.; Li, M.; Zhi, F.; Liu, S.; et al. Gut Microbiota Offers Universal Biomarkers across Ethnicity in Inflammatory Bowel Disease Diagnosis and Infliximab Response Prediction. mSystems 2018, 3, e00188-17. [Google Scholar] [CrossRef] [PubMed]
- Aden, K.; Rehman, A.; Waschina, S.; Pan, W.H.; Walker, A.; Lucio, M.; Nunez, A.M.; Bharti, R.; Zimmerman, J.; Bethge, J.; et al. Metabolic Functions of Gut Microbes Associate With Efficacy of Tumor Necrosis Factor Antagonists in Patients With Inflammatory Bowel Diseases. Gastroenterology 2019, 157, 1279–1292.e1211. [Google Scholar] [CrossRef]
- Dovrolis, N.; Michalopoulos, G.; Theodoropoulos, G.E.; Arvanitidis, K.; Kolios, G.; Sechi, L.A.; Eliopoulos, A.G.; Gazouli, M. The Interplay between Mucosal Microbiota Composition and Host Gene-Expression is Linked with Infliximab Response in Inflammatory Bowel Diseases. Microorganisms 2020, 8, 438. [Google Scholar] [CrossRef]
- Schierova, D.; Roubalova, R.; Kolar, M.; Stehlikova, Z.; Rob, F.; Jackova, Z.; Coufal, S.; Thon, T.; Mihula, M.; Modrak, M.; et al. Fecal Microbiome Changes and Specific Anti-Bacterial Response in Patients with IBD during Anti-TNF Therapy. Cells 2021, 10, 3188. [Google Scholar] [CrossRef]
- Sanchis-Artero, L.; Martínez-Blanch, J.F.; Manresa-Vera, S.; Cortés-Castell, E.; Valls-Gandia, M.; Iborra, M.; Paredes-Arquiola, J.M.; Boscá-Watts, M.; Huguet, J.M.; Gil-Borrás, R.; et al. Evaluation of changes in intestinal microbiota in Crohn’s disease patients after anti-TNF alpha treatment. Sci. Rep. 2021, 11, 10016. [Google Scholar] [CrossRef]
- Effenberger, M.; Reider, S.; Waschina, S.; Bronowski, C.; Enrich, B.; Adolph, T.E.; Koch, R.; Moschen, A.R.; Rosenstiel, P.; Aden, K.; et al. Microbial Butyrate Synthesis Indicates Therapeutic Efficacy of Azathioprine in IBD Patients. J. Crohns Colitis 2021, 15, 88–98. [Google Scholar] [CrossRef]
- Park, Y.E.; Moon, H.S.; Yong, D.; Seo, H.; Yang, J.; Shin, T.S.; Kim, Y.K.; Kim, J.R.; Lee, Y.N.; Kim, Y.H.; et al. Microbial changes in stool, saliva, serum, and urine before and after anti-TNF-α therapy in patients with inflammatory bowel diseases. Sci. Rep. 2022, 12, 6359. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Day, A.S.; Leach, S.T.; Lemberg, D.A.; Nielsen, S.; Mitchell, H.M. Effect of exclusive enteral nutrition on the microbiota of children with newly diagnosed Crohn’s disease. Clin. Transl. Gastroenterol. 2015, 6, e71. [Google Scholar] [CrossRef] [PubMed]
- Quince, C.; Ijaz, U.Z.; Loman, N.; Eren, A.M.; Saulnier, D.; Russell, J.; Haig, S.J.; Calus, S.T.; Quick, J.; Barclay, A.; et al. Extensive Modulation of the Fecal Metagenome in Children with Crohn’s Disease During Exclusive Enteral Nutrition. Am. J. Gastroenterol. 2015, 110, 1718–1729; quiz 1730. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Huang, Y.; Shi, P.; Wang, Y.; Zhang, Y.; Xue, A.; Tang, Z.; Hu, W.; Sun, H.; Zhang, P.; et al. Effect of Exclusive Enteral Nutrition on the Disease Process, Nutrition Status, and Gastrointestinal Microbiota for Chinese Children with Crohn’s Disease. J. Parenter. Enter. Nutr. 2021, 45, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.A.; Moore-Connors, J.; MacIntyre, B.; Stadnyk, A.W.; Thomas, N.A.; Noble, A.; Mahdi, G.; Rashid, M.; Otley, A.R.; Bielawski, J.P.; et al. Early Changes in Microbial Community Structure Are Associated with Sustained Remission after Nutritional Treatment of Pediatric Crohn’s Disease. Inflamm. Bowel Dis. 2016, 22, 2853–2862. [Google Scholar] [CrossRef]
- Costa-Santos, M.P.; Palmela, C.; Torres, J.; Ferreira, A.; Velho, S.; Ourô, S.; Glória, L.; Gordo, I.; Maio, R.; Cravo, M. Preoperative enteral nutrition in adults with complicated Crohn’s disease: Effect on disease outcomes and gut microbiota. Nutrition 2020, 70s, 100009. [Google Scholar] [CrossRef]
- Diederen, K.; Li, J.V.; Donachie, G.E.; de Meij, T.G.; de Waart, D.R.; Hakvoort, T.B.M.; Kindermann, A.; Wagner, J.; Auyeung, V.; Te Velde, A.A.; et al. Exclusive enteral nutrition mediates gut microbial and metabolic changes that are associated with remission in children with Crohn’s disease. Sci. Rep. 2020, 10, 18879. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, L.; Chen, Y.; Chen, H. Exclusive enteral nutrition remodels the intestinal flora in patients with active Crohn’s disease. BMC Gastroenterol. 2022, 22, 212. [Google Scholar] [CrossRef]
- Suskind, D.L.; Lee, D.; Kim, Y.M.; Wahbeh, G.; Singh, N.; Braly, K.; Nuding, M.; Nicora, C.D.; Purvine, S.O.; Lipton, M.S.; et al. The Specific Carbohydrate Diet and Diet Modification as Induction Therapy for Pediatric Crohn’s Disease: A Randomized Diet Controlled Trial. Nutrients 2020, 12, 3749. [Google Scholar] [CrossRef]
- Wilson, B.; Eyice, Ö.; Koumoutsos, I.; Lomer, M.C.; Irving, P.M.; Lindsay, J.O.; Whelan, K. Prebiotic Galactooligosaccharide Supplementation in Adults with Ulcerative Colitis: Exploring the Impact on Peripheral Blood Gene Expression, Gut Microbiota, and Clinical Symptoms. Nutrients 2021, 13, 3598. [Google Scholar] [CrossRef]
- Chen, X.; Lou, L.; Tang, H.; Tan, X.; Bi, J.; Wu, H.; Li, N.; Wang, Y.; Mao, J. Adsorptive granulomonocytapheresis alters the gut bacterial microbiota in patients with active ulcerative colitis. J. Clin. Apher. 2021, 36, 454–464. [Google Scholar] [CrossRef]
- Olendzki, B.; Bucci, V.; Cawley, C.; Maserati, R.; McManus, M.; Olednzki, E.; Madziar, C.; Chiang, D.; Ward, D.V.; Pellish, R.; et al. Dietary manipulation of the gut microbiome in inflammatory bowel disease patients: Pilot study. Gut Microbes 2022, 14, 2046244. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.D.; Van Domselaar, G.; Bernstein, C.N. The Gut Microbiota in Immune-Mediated Inflammatory Diseases. Front. Microbiol. 2016, 7, 1081. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Béguet-Crespel, F.; Marinelli, L.; Jamet, A.; Ledue, F.; Blottière, H.M.; Lapaque, N. Butyrate produced by gut commensal bacteria activates TGF-beta1 expression through the transcription factor SP1 in human intestinal epithelial cells. Sci. Rep. 2018, 8, 9742. [Google Scholar] [CrossRef]
- Paramsothy, S.; Nielsen, S.; Kamm, M.A.; Deshpande, N.P.; Faith, J.J.; Clemente, J.C.; Paramsothy, R.; Walsh, A.J.; van den Bogaerde, J.; Samuel, D.; et al. Specific Bacteria and Metabolites Associated with Response to Fecal Microbiota Transplantation in Patients with Ulcerative Colitis. Gastroenterology 2019, 156, 1440–1454.e1442. [Google Scholar] [CrossRef]


| Disease Subtype | Reference | Population | Microbiota Changes due to Treatment |
|---|---|---|---|
| Biological therapies | |||
| CD CD CD | Wang et al. [38] | Paediatric (P) CD (n = 4) | Following IFX treatment:
|
| Zhou et al. [57] | Adult (A) CD (n = 16) | Following IFX treatment:
| |
| Dovrolis et al. [59] | A -CD (n = 10) | Following IFX treatment, CD patients:
| |
| Kowalska-Duplaga et al. [54] | P -CD (n = 18) | Following IFX treatment:
| |
| Salamon et al. [55] | P -CD (n = 13) | Following IFX treatment:
| |
| Zhuang et al. [40] | A -CD (n = 49) | Following IFX treatment:
| |
| Wang et al. [39] | P -CD (n = 11) | Following IFX treatment:
| |
| Ribaldone et al. [53] | A -CD (n = 20) | Following ADA treatment:
| |
| Ananthakrishnan et al. [42] | A -CD (n = 42) | Following Vedolizumab treatment: In CD remission patients (n = 10):
| |
| Doherty et al. [41] | A-CD(n = 306) | Following UST treatment, in subjects in remission 6 weeks after induction compared to those with active disease:
| |
| Sanchis-Artero et al. [61] | CD R (n = 13) CD NR(n = 14) | Following IFX/ADA treatment:
| |
| CD | Ventin-Holmberg et al. [56] | CD R (n = 12) CD NR(n = 18) | Following IFX treatment:
|
| UC | Dovrolis et al. [59] | A -UC (n = 4) | Following IFX treatment, in UC patients:
|
| IBD | Kolho et al. [43] | P -IBD (n = 11) | Following IFX/ADA treatment:
|
| Aden et al. [58] | A-IBD (n = 12) | Following anti-TNF Etanercept treatment:
| |
| Ding et al. [48] | Adult CD (n = 76) UC (n = 10) | Following anti-TNF-alpha treatment:
| |
| Olbjorn et al. [50] | P -CD (n = 22) UC (n = 9) | Following anti-TNF-alpha treatment:
| |
| Schierova et al. [60] | CD (n = 17) UC (n= 10) | Following IFX/ADA treatment:
| |
| Ventin-Holmberg et al. [49] | CD R (n = 13) CD PR (n = 4) CD NR (n = 6) UC R (n = 31) UC PR (n = 8) UC NR (n = 8) | Following IFX treatment:
| |
| Effenberger et al. [62] | CD R (n = 13) CD NR(n = 11) UC R (n = 7) UC NR (n = 5) | Following anti-TNF treatment:
| |
| IBD | Park et al. [63] | UC (n = 10) CD (n = 9) | Following anti-TNF treatment:
|
| Non-biological therapies | |||
| CD | Pigneur et al. [34] | P -CD (n = 4) | Following Corticosteroid treatment:
|
| Sprockett et al. [36] | P -CD (n = 74) | Following antibiotic treatment: Metronidazole:
| |
| UC | Hart et al. [35] | P -UC (n = 10) | Following corticosteroid treatment:
|
| Ishikawa et al. [37] | A -UC (n = 19) | Following amoxicillin, fosfomycin and metronidazole treatment:
| |
| Schierova et al. [33] | UC (n = 8) | Following 5-ASA treatment, no changes in faecal microbiome observed. | |
| IBD | Hart et al. [35] | Paediatric CD (n = 4) UC (n = 10) | Following corticosteroid treatment:
|
| Effenberger et al. [62] | CD R (n = 10) CD NR (n = 9) UC R (n = 5) UC NR (n = 5) | Following AZA treatment:
| |
| Nutritional therapies | |||
| CD | Kaakoush et al. [64] | Paediatric CD (n = 5) HC (n = 5) | Following EEN:
|
| Quince et al. [65] | Paediatric CD (n = 23) HC (n = 21) | Following EEN:
| |
| Tang et al. [66] | Paediatric CD remission (n = 17) CD non-remission (n = 8) HC (n = 12) | Following EEN:
| |
| Dunn et al. [67] | Paediatric CD (n = 10) HC (n = 5) | Following EEN:
| |
| Costa-Santos et al. [68] | A -CD (n = 10) | Following EEN:
| |
| Diederen et al. [69] | P -CD (n = 43) | Following EEN:
| |
| Suskind et al. [71] | P -CD (n = 5) | Following SCD:
| |
| Tang et al. [66] | CD (n = 31) HC (n = 12) | Following EEN treatment:
| |
| Jiang et al. [70] | CD (n = 7) | Following EEN treatment:
| |
| UC | Valcheva et al. [51] | A -UC (n = 25) | Following prebiotics (inulin-type fructans):
|
| Wilson et al. [72] | UC (n = 17) | Following GOS prebiotic treatment:
| |
| Chen et al. [73] | UC (n = 14) HC (n = 14) | Following GMA treatment:
| |
| IBD | Olendzki et al. [74] | CD (n = 14) UC (n = 7) | After dietary intervention:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mah, C.; Jayawardana, T.; Leong, G.; Koentgen, S.; Lemberg, D.; Connor, S.J.; Rokkas, T.; Grimm, M.C.; Leach, S.T.; Hold, G.L. Assessing the Relationship between the Gut Microbiota and Inflammatory Bowel Disease Therapeutics: A Systematic Review. Pathogens 2023, 12, 262. https://doi.org/10.3390/pathogens12020262
Mah C, Jayawardana T, Leong G, Koentgen S, Lemberg D, Connor SJ, Rokkas T, Grimm MC, Leach ST, Hold GL. Assessing the Relationship between the Gut Microbiota and Inflammatory Bowel Disease Therapeutics: A Systematic Review. Pathogens. 2023; 12(2):262. https://doi.org/10.3390/pathogens12020262
Chicago/Turabian StyleMah, Cassandra, Thisun Jayawardana, Gary Leong, Sabrina Koentgen, Daniel Lemberg, Susan J. Connor, Theodore Rokkas, Michael C. Grimm, Steven T. Leach, and Georgina L. Hold. 2023. "Assessing the Relationship between the Gut Microbiota and Inflammatory Bowel Disease Therapeutics: A Systematic Review" Pathogens 12, no. 2: 262. https://doi.org/10.3390/pathogens12020262