

Compositional Differences of Meconium Microbiomes of Preterm and Term Infants, and Infants That Developed Necrotizing Enterocolitis or Feeding Intolerance
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Collection
2.2. Fecal Microbiome Analysis; DNA Extraction and Amplification
2.3. Sequencing and Data Analysis
2.4. Definitions
2.5. Statistical Analysis
3. Results
3.1. Clinical Characteristics
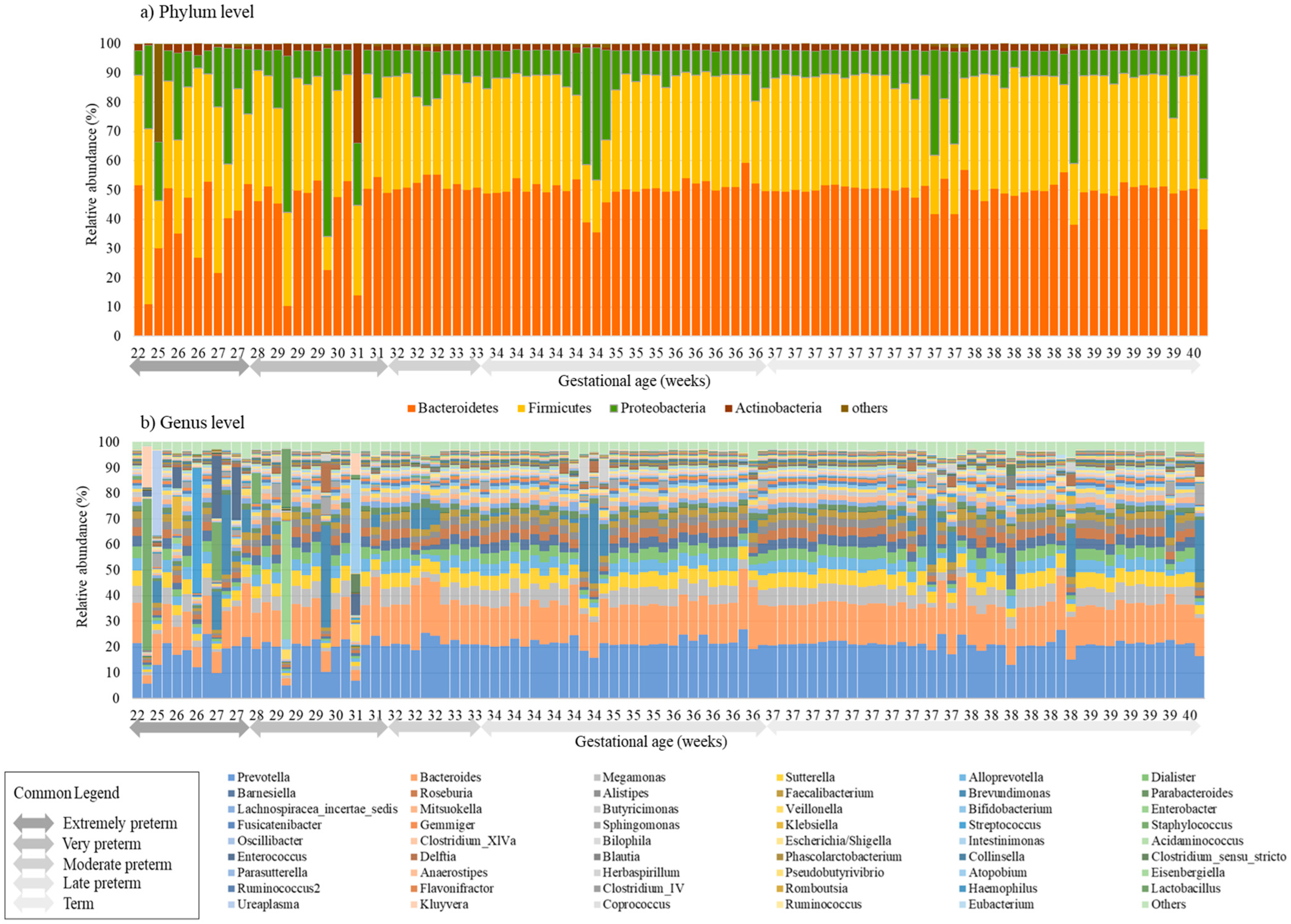
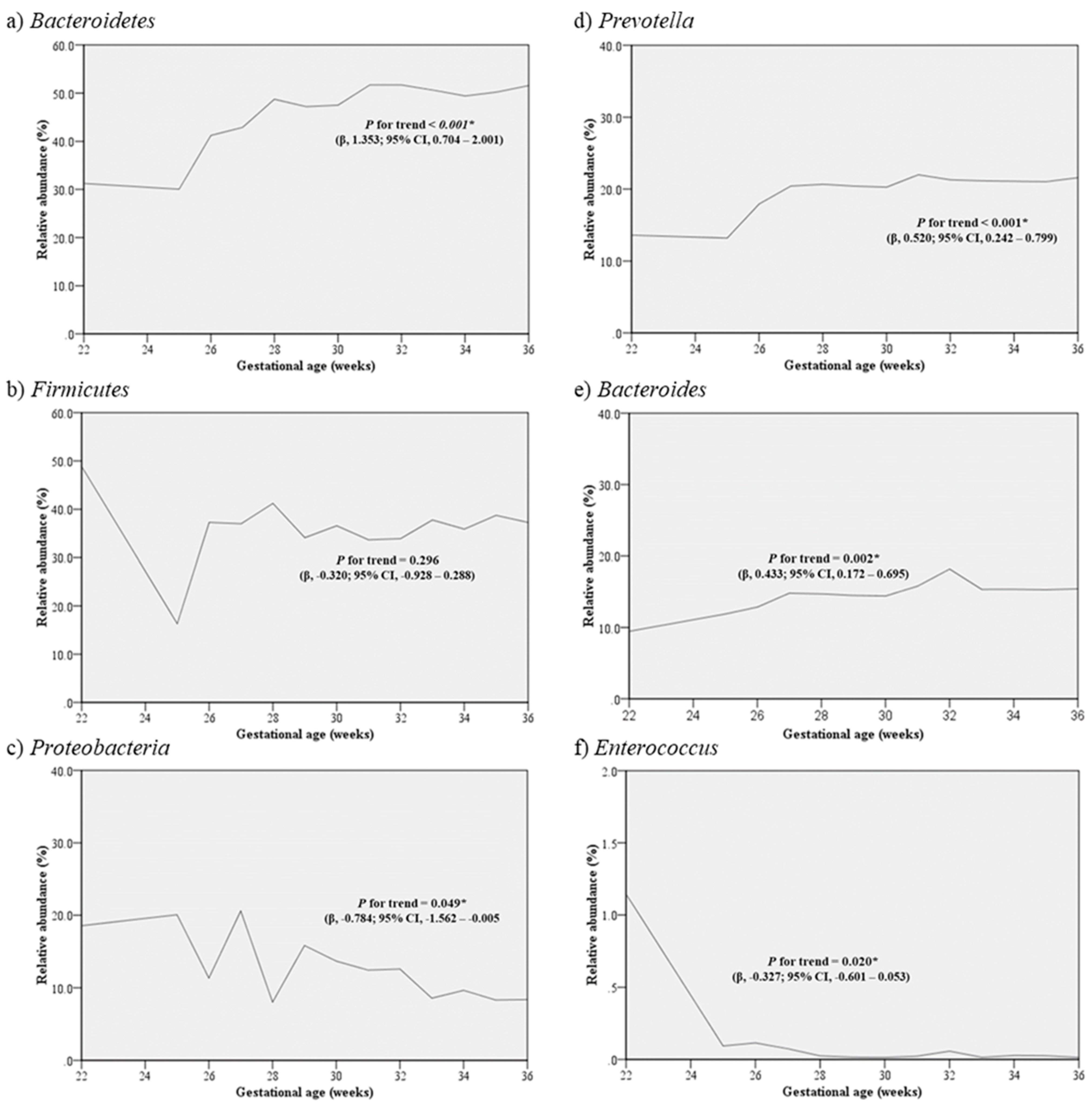
3.2. Distribution of Microbiota according to Gestational Age
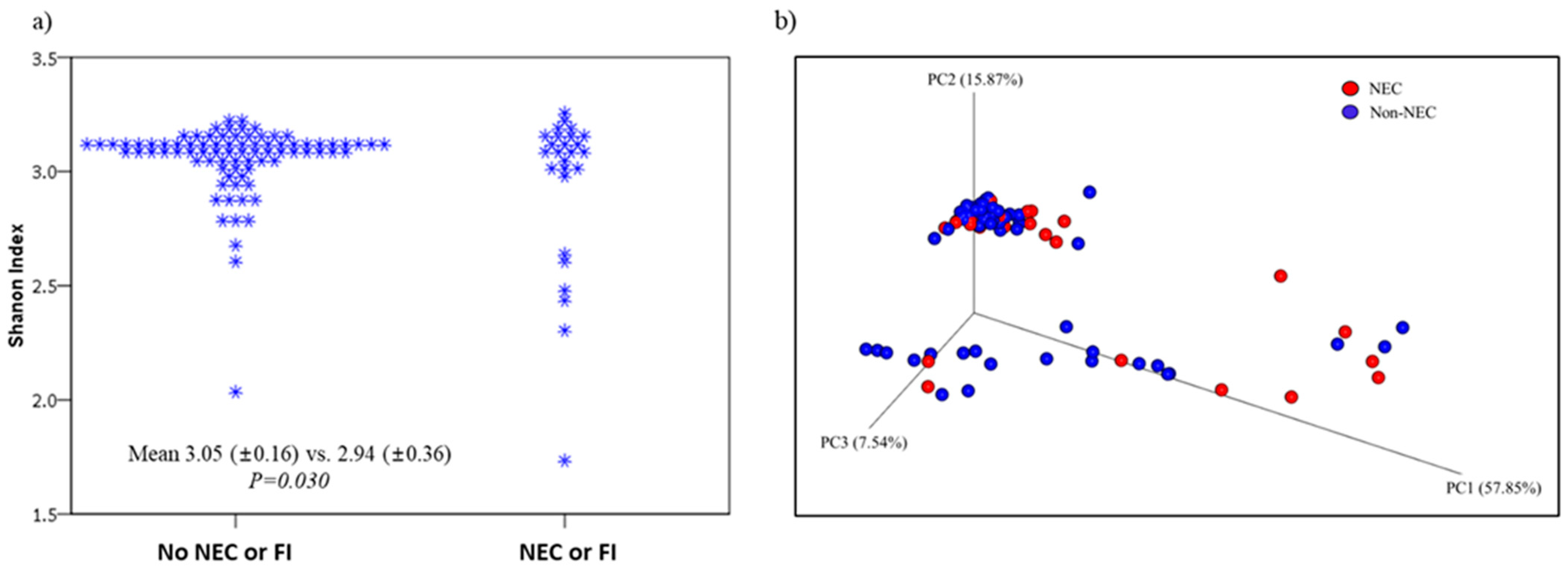
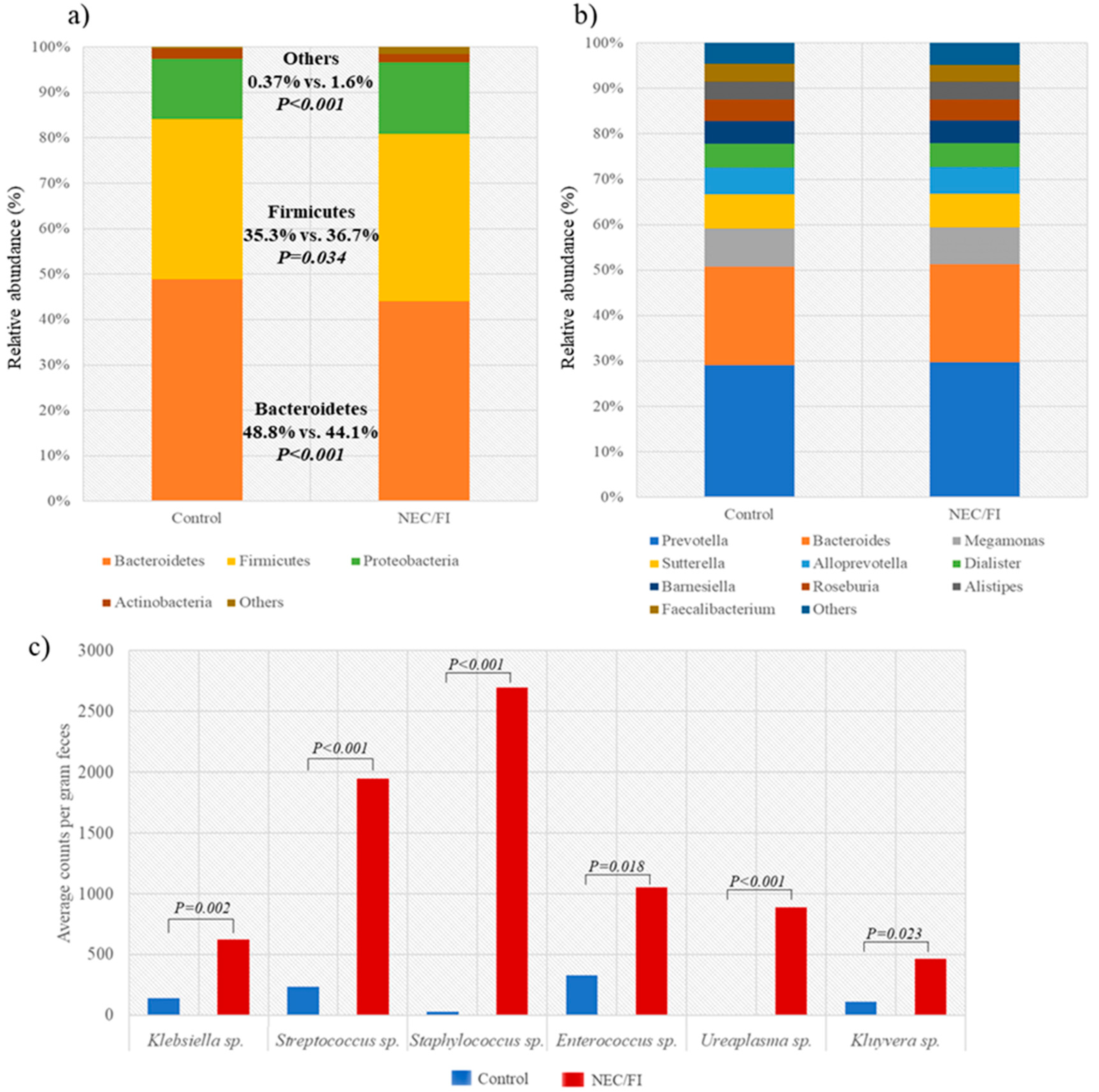
3.3. Comparison of Microbiomes and Clinical Outcomes between NEC/FI Group Versus Control Group
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chung, S.H.; Bae, C.W. Improvement in the Survival Rates of Very Low Birth Weight Infants after the Establishment of the Korean Neonatal Network: Comparison between the 2000s and 2010s. J. Korean Med. Sci. 2017, 32, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.J. Epidemiology of necrotizing enterocolitis. Clin. Perinatol. 1994, 21, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.H.; Gordon, P.; Walker, W.M.; Laughon, M.; Smith, P.B.; Spitzer, A.R. Characteristics of patients who die of necrotizing enterocolitis. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2012, 32, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Schnabl, K.L.; Van Aerde, J.E.; Thomson, A.B.; Clandinin, M.T. Necrotizing enterocolitis: A multifactorial disease with no cure. World J. Gastroenterol. 2008, 14, 2142–2161. [Google Scholar] [CrossRef]
- Thompson, A.M.; Bizzarro, M.J. Necrotizing enterocolitis in newborns: Pathogenesis, prevention and management. Drugs 2008, 68, 1227–1238. [Google Scholar] [CrossRef]
- Neu, J.; Walker, W.A. Necrotizing enterocolitis. New Engl. J. Med. 2011, 364, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Perez-Muñoz, M.E.; Arrieta, M.C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, K.M.; Gerlach, M.J.; Adam, T.; Heimesaat, M.M.; Rossi, L.; Surette, M.G.; Sloboda, D.M.; Braun, T. Fetal meconium does not have a detectable microbiota before birth. Nat. Microbiol. 2021, 6, 865–873. [Google Scholar] [CrossRef]
- Stinson, L.F.; Boyce, M.C.; Payne, M.S.; Keelan, J.A. The Not-so-Sterile Womb: Evidence That the Human Fetus Is Exposed to Bacteria Prior to Birth. Front. Microbiol. 2019, 10, 1124. [Google Scholar] [CrossRef]
- He, Q.; Kwok, L.Y.; Xi, X.; Zhong, Z.; Ma, T.; Xu, H.; Meng, H.; Zhao, F.; Zhang, H. The meconium microbiota shares more features with the amniotic fluid microbiota than the maternal fecal and vaginal microbiota. Gut Microbes 2020, 12, 1794266. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Forster, S.C.; Tsaliki, E.; Vervier, K.; Strang, A.; Simpson, N.; Kumar, N.; Stares, M.D.; Rodger, A.; Brocklehurst, P.; et al. Stunted microbiota and opportunistic pathogen colonization in caesarean-section birth. Nature 2019, 574, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Dalby, M.J.; Hall, L.J. Recent advances in understanding the neonatal microbiome. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Klopp, J.; Ferretti, P.; Meyer, C.U.; Hilbert, K.; Haiß, A.; Marißen, J.; Henneke, P.; Hudalla, H.; Pirr, S.; Viemann, D.; et al. Meconium Microbiome of Very Preterm Infants across Germany. mSphere. 2022, 7, e00808-21. [Google Scholar] [CrossRef]
- Shi, Y.-C.; Guo, H.; Chen, J.; Sun, G.; Ren, R.-R.; Guo, M.-Z.; Peng, L.-H.; Yang, Y.-S. Initial meconium microbiome in Chinese neonates delivered naturally or by cesarean section. Sci. Rep. 2018, 8, 3255. [Google Scholar] [CrossRef] [Green Version]
- González-Rivera, R.; Culverhouse, R.C.; Hamvas, A.; Tarr, P.I.; Warner, B.B. The age of necrotizing enterocolitis onset: An application of Sartwell's incubation period model. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2011, 31, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Hodzic, Z.; Bolock, A.M.; Good, M. The Role of Mucosal Immunity in the Pathogenesis of Necrotizing Enterocolitis. Front. Pediatr. 2017, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Niño, D.F.; Sodhi, C.P.; Hackam, D.J. Necrotizing enterocolitis: New insights into pathogenesis and mechanisms. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 590–600. [Google Scholar] [CrossRef] [Green Version]
- 16S Metagenomic Sequencing Library Preparation. Available online: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 20 July 2022).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, M.J.; Ternberg, J.L.; Feigin, R.D.; Keating, J.P.; Marshall, R.; Barton, L.; Brotherton, T. Neonatal necrotizing enterocolitis. Therapeutic decisions based upon clinical staging. Ann. Surg. 1978, 187, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Carlos, M.A.; Babyn, P.S.; Marcon, M.A.; Moore, A.M. Changes in gastric emptying in early postnatal life. J. Pediatr. 1997, 130, 931–937. [Google Scholar] [CrossRef] [PubMed]
- WHO Factsheet. Preterm Birth. Available online: https://www.who.int/news-room/fact-sheets/detail/preterm-birth (accessed on 31 November 2022).
- Samuels, N.; van de Graaf, R.A.; de Jonge, R.C.J.; Reiss, I.K.M.; Vermeulen, M.J. Risk factors for necrotizing enterocolitis in neonates: A systematic review of prognostic studies. BMC Pediatr. 2017, 17, 105. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neish, A.S. Microbes in gastrointestinal health and disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Scarpellini, E.; Ianiro, G.; Attili, F.; Bassanelli, C.; De Santis, A.; Gasbarrini, A. The human gut microbiota and virome: Potential therapeutic implications. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2015, 47, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Ringel-Kulka, T.; Heikamp-de Jong, I.; Ringel, Y.; Carroll, I.; de Vos, W.M.; Salojärvi, J.; Satokari, R. Discordant temporal development of bacterial phyla and the emergence of core in the fecal microbiota of young children. ISME J. 2016, 10, 1002–1014. [Google Scholar] [CrossRef] [Green Version]
- Jalanka-Tuovinen, J.; Salonen, A.; Nikkilä, J.; Immonen, O.; Kekkonen, R.; Lahti, L.; Palva, A.; de Vos, W.M. Intestinal microbiota in healthy adults: Temporal analysis reveals individual and common core and relation to intestinal symptoms. PloS ONE 2011, 6, e23035. [Google Scholar] [CrossRef]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4578–4585. [Google Scholar] [CrossRef] [Green Version]
- Asnicar, F.; Manara, S.; Zolfo, M.; Truong, D.T.; Scholz, M.; Armanini, F.; Ferretti, P.; Gorfer, V.; Pedrotti, A.; Tett, A.; et al. Studying Vertical Microbiome Transmission from Mothers to Infants by Strain-Level Metagenomic Profiling. mSystems 2017, 2, e00164-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korpela, K.; Costea, P.; Coelho, L.P.; Kandels-Lewis, S.; Willemsen, G.; Boomsma, D.I.; Segata, N.; Bork, P. Selective maternal seeding and environment shape the human gut microbiome. Genome Res. 2018, 28, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, J.; Marques, C.; Teixeira, D.; Durão, C.; Faria, A.; Brito, S.; Cardoso, M.; Macedo, I.; Pereira, E.; Tomé, T.; et al. Extremely preterm neonates have more Lactobacillus in meconium than very preterm neonates—The in utero microbial colonization hypothesis. Gut Microbes 2020, 12, 1785804. [Google Scholar] [CrossRef]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low diversity gut microbiota dysbiosis: Drivers, functional implications and recovery. Curr. Opin. Microbiol. 2018, 44, 34–40. [Google Scholar] [CrossRef]
- Hamilton, M.J.; Weingarden, A.R.; Unno, T.; Khoruts, A.; Sadowsky, M.J. High-throughput DNA sequence analysis reveals stable engraftment of gut microbiota following transplantation of previously frozen fecal bacteria. Gut Microbes 2013, 4, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Stombaugh, J.; Gonzalez, A.; Ackermann, G.; Wendel, D.; Vázquez-Baeza, Y.; Jansson, J.K.; Gordon, J.I.; Knight, R. Meta-analyses of studies of the human microbiota. Genome Res. 2013, 23, 1704–1714. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Frank, D.N.; Horch, M.; Chau, S.; Ir, D.; Horch, E.A.; Tretina, K.; van Besien, K.; Lozupone, C.A.; Nguyen, V.H. Associations between acute gastrointestinal GvHD and the baseline gut microbiota of allogeneic hematopoietic stem cell transplant recipients and donors. Bone Marrow Transplant. 2017, 52, 1643–1650. [Google Scholar] [CrossRef] [Green Version]
- Terrazzan Nutricionist, A.C.; Procianoy, R.S.; Roesch, L.F.W.; Corso, A.L.; Dobbler, P.T.; Silveira, R.C. Meconium microbiome and its relation to neonatal growth and head circumference catch-up in preterm infants. PloS ONE 2020, 15, e0238632. [Google Scholar] [CrossRef]
- Liu, L.; Ao, D.; Cai, X.; Huang, P.; Cai, N.; Lin, S.; Wu, B. Early gut microbiota in very low and extremely low birth weight preterm infants with feeding intolerance: A prospective case-control study. J. Microbiol. 2022, 60, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Sharif, S.; Meader, N.; Oddie, S.J.; Rojas-Reyes, M.X.; McGuire, W. Probiotics to prevent necrotising enterocolitis in very preterm or very low birth weight infants. Cochrane Database Syst Rev 2020, 10, CD005496. [Google Scholar] [CrossRef] [PubMed]
- Amandito, R.; Malik, A.; Rohsiswatmo, R. Metagenomic profiles of the early life microbiome of Indonesian inpatient neonates and their influence on clinical characteristics. Sci. Rep. 2022, 12, 9413. [Google Scholar] [CrossRef]
- Jiang, L.; Stärkel, P.; Fan, J.G.; Fouts, D.E.; Bacher, P.; Schnabl, B. The gut mycobiome: A novel player in chronic liver diseases. J. Gastroenterol. 2021, 56, 1–11. [Google Scholar] [CrossRef] [PubMed]
- James, S.A.; Phillips, S.; Telatin, A.; Baker, D.; Ansorge, R.; Clarke, P.; Hall, L.J.; Carding, S.R. Preterm Infants Harbour a Rapidly Changing Mycobiota That Includes Candida Pathobionts. J. Fungi 2020, 6, 273. [Google Scholar] [CrossRef]
- Kim, Y.S.; Unno, T.; Kim, B.Y.; Park, M.S. Sex Differences in Gut Microbiota. World J. Men’s Health 2020, 38, 48–60. [Google Scholar] [CrossRef]
- Lay, C.; Rigottier-Gois, L.; Holmstrøm, K.; Rajilic, M.; Vaughan, E.E.; de Vos, W.M.; Collins, M.D.; Thiel, R.; Namsolleck, P.; Blaut, M.; et al. Colonic microbiota signatures across five northern European countries. Appl. Environ. Microbiol. 2005, 71, 4153–4155. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Participants (%) | P Value | |||
|---|---|---|---|---|
| Total N = 108 | Preterm N = 63 | Term N = 45 | ||
| GA, weeks (mean ± SD) | 34.7 ± 4.2 | 31.6 ± 3.7 | 37.9 ± 1.0 | <0.001 |
| Birth weight, g (mean ± SD) | 2,305.9 ± 863.0 | 1,807.4 ± 773.0 | 3,003.8 ± 339.3 | <0.001 |
| Mother DM | 11 (10.2) | 8 (12.7) | 3 (6.7) | 0.354 |
| IVF | 6 (5.6) | 5 (7.9) | 1 (2.2) | 0.397 |
| Male | 96 (88.9) | 60 (95.2) | 36 (80.0) | 0.026 |
| Caesarean section | 90 (83.3) | 58 (92.1) | 32 (71.1) | 0.007 |
| Small for gestational age | 6 (5.6) | 3 (4.8) | 3 (6.7) | 0.692 |
| Chorioamnionitis | 2 (1.9) | 2 (3.2) | 0 | 0.509 |
| PROM, hrs | 14 (13.0) | 11 (17.5) | 2 (4.4) | 0.069 |
| Placental abruption | 6 (5.6) | 6 (9.5) | 0 | 0.040 |
| Oligohydramnios | 10 (9.3) | 10 (15.9) | 0 | 0.005 |
| Antenatal steroid use | 43 (39.8) | 43 (68.3) | 0 | <0.001 |
| Congenital infection | 7 (6.5) | 7 (11.1) | 0 | 0.040 |
| Need for resuscitation at birth | 50 (46.3) | 40 (63.5) | 10 (22.2) | <0.001 |
| NEC/FI | 26 (24.1) | 22 (34.9) | 4 (8.9) | 0.003 |
| Univariate | P Value | Multivariate | P Value | |||
|---|---|---|---|---|---|---|
| OR | 95% CI | OR | 95% CI | |||
| GA, weeks (mean ± SD) | 0.716 | 0.624–0.820 | <0.001 | 0.736 | 0.491–1.104 | 0.138 |
| Birth weight, g (mean ± SD) | 0.998 | 0.998–0.999 | <0.001 | 0.999 | 0.997–1.001 | 0.162 |
| Caesarean section | 1.716 | 0.455–6.470 | 0.425 | |||
| PROM | 3.214 | 0.971–10.638 | 0.056 | |||
| Preeclampsia | 0.893 | 0.174–4.590 | 0.892 | |||
| Placental abruption | 3.435 | 0.649–18.179 | 0.147 | |||
| Oligohydramnios | 3.667 | 0.970–13.866 | 0.056 | |||
| IUGR | 3.435 | 0.649–18.179 | 0.147 | |||
| Chorioamnionitis | - | - | 0.999 | |||
| Antenatal steroid use | 4.068 | 1.602–10.335 | 0.003 | 0.376 | 0.074–1.913 | 0.239 |
| Congenital infection | 9.524 | 1.725–52.593 | 0.010 | 1.275 | 0.179–9.097 | 0.808 |
| RDS | 4.227 | 1.673–10.682 | 0.002 | 0.207 | 0.033–1.309 | 0.094 |
| Massive pulmonary hemorrhage | - | - | 0.999 | |||
| Air leak | 1.283 | 0.234–7.043 | 0.774 | |||
| Sepsis | - | - | 0.998 | |||
| IVH > grade II | 2.187 | 1.126–4.247 | 0.021 | 0.595 | 0.227–1.560 | 0.291 |
| PDA | 2.194 | 0.346–13.909 | 0.404 | |||
| CLD | - | - | 0.998 | |||
| PAH | 3.333 | 0.446–24.936 | 0.241 | |||
| No breast milk given enterally prior to NEC/FI | 0.406 | 0.147–1.119 | 0.081 | |||
| Antibiotics use prior to NEC/FI | - | - | 0.999 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.M.; Kim, S.; Hwang-Bo, S.; Yoo, I.H.; Seo, Y.-M.; Oh, M.Y.; Im, S.-A.; Youn, Y.-A. Compositional Differences of Meconium Microbiomes of Preterm and Term Infants, and Infants That Developed Necrotizing Enterocolitis or Feeding Intolerance. Pathogens 2023, 12, 55. https://doi.org/10.3390/pathogens12010055
Kang HM, Kim S, Hwang-Bo S, Yoo IH, Seo Y-M, Oh MY, Im S-A, Youn Y-A. Compositional Differences of Meconium Microbiomes of Preterm and Term Infants, and Infants That Developed Necrotizing Enterocolitis or Feeding Intolerance. Pathogens. 2023; 12(1):55. https://doi.org/10.3390/pathogens12010055
Chicago/Turabian StyleKang, Hyun Mi, Sol Kim, Seok Hwang-Bo, In Hyuk Yoo, Yu-Mi Seo, Moon Yeon Oh, Soo-Ah Im, and Young-Ah Youn. 2023. "Compositional Differences of Meconium Microbiomes of Preterm and Term Infants, and Infants That Developed Necrotizing Enterocolitis or Feeding Intolerance" Pathogens 12, no. 1: 55. https://doi.org/10.3390/pathogens12010055