
Biomonitoring of Indoor Air Fungal or Chemical Toxins with Caenorhabditis elegans nematodes
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Stress-Responsive C. elegans Reporter Strains Sense Fungal Toxins
2.2. C. elegans Responses Correlate with Those Obtained from Single Cell-Based Assays
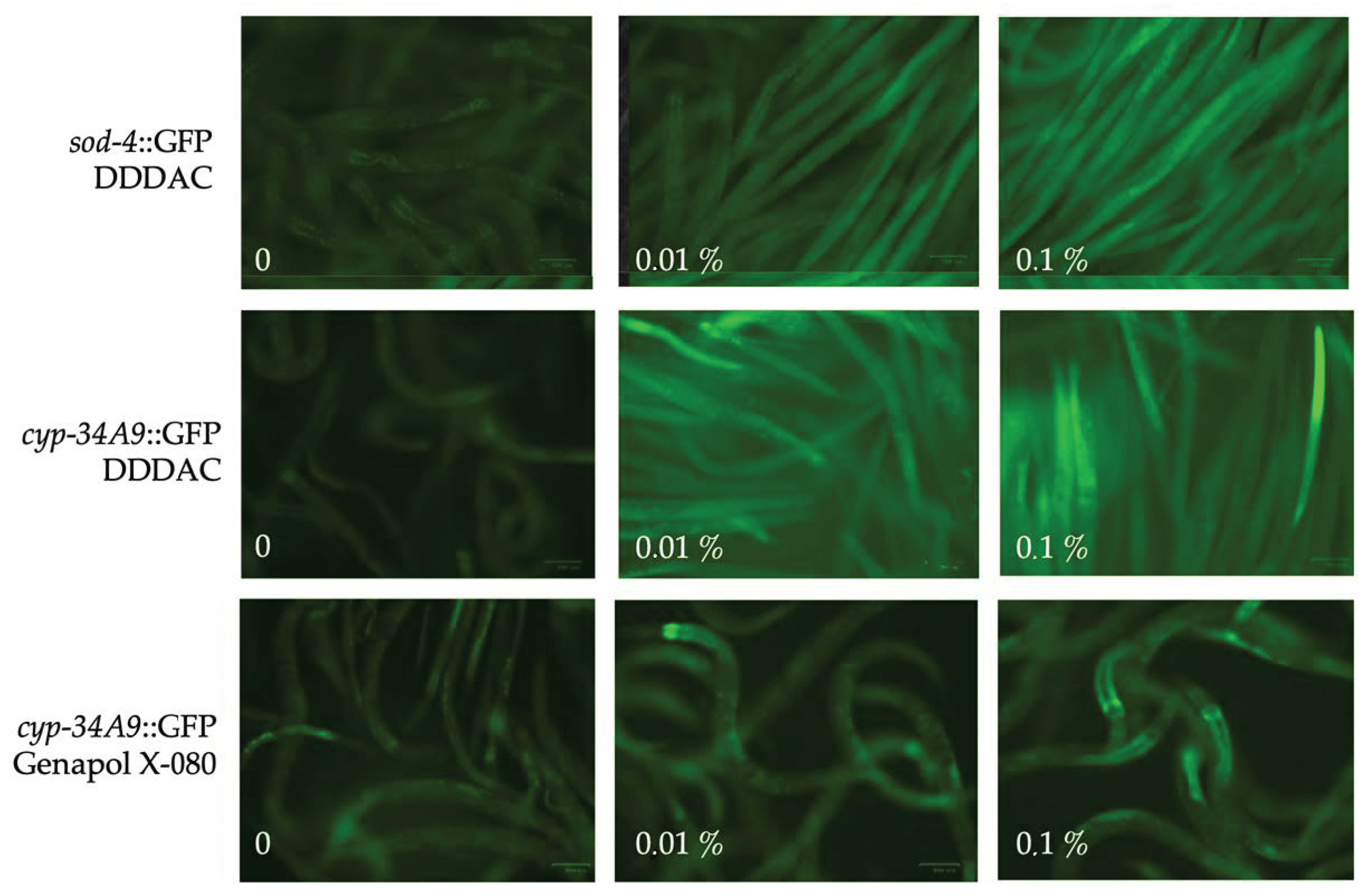
2.3. Indoor Chemicals Reduce C. elegans Motility in a Dose-Dependent Fashion
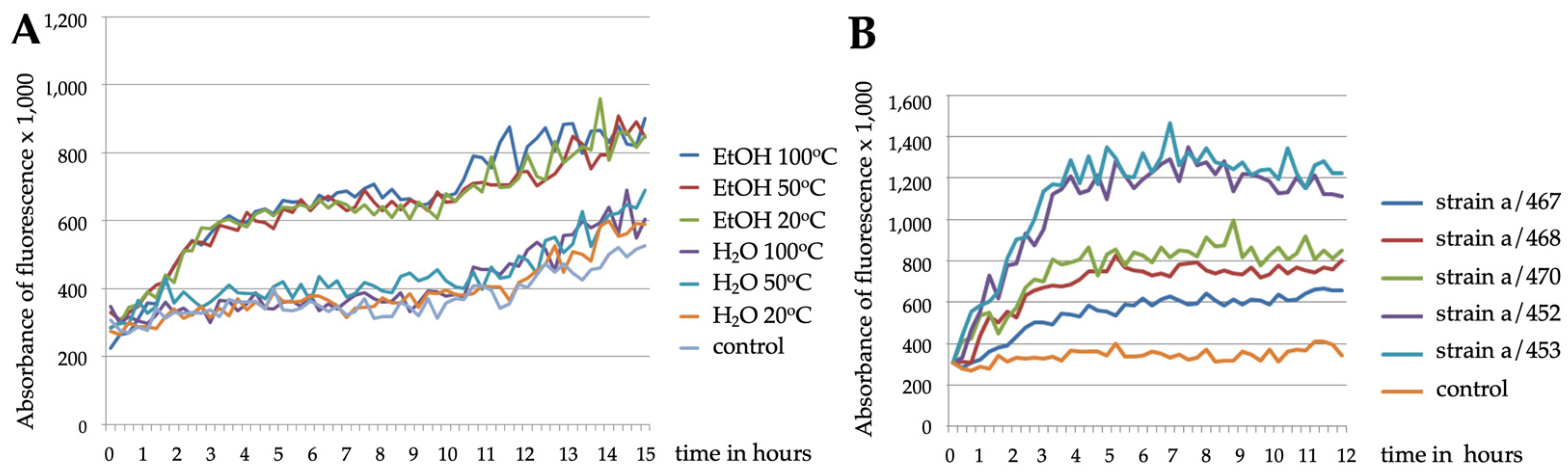
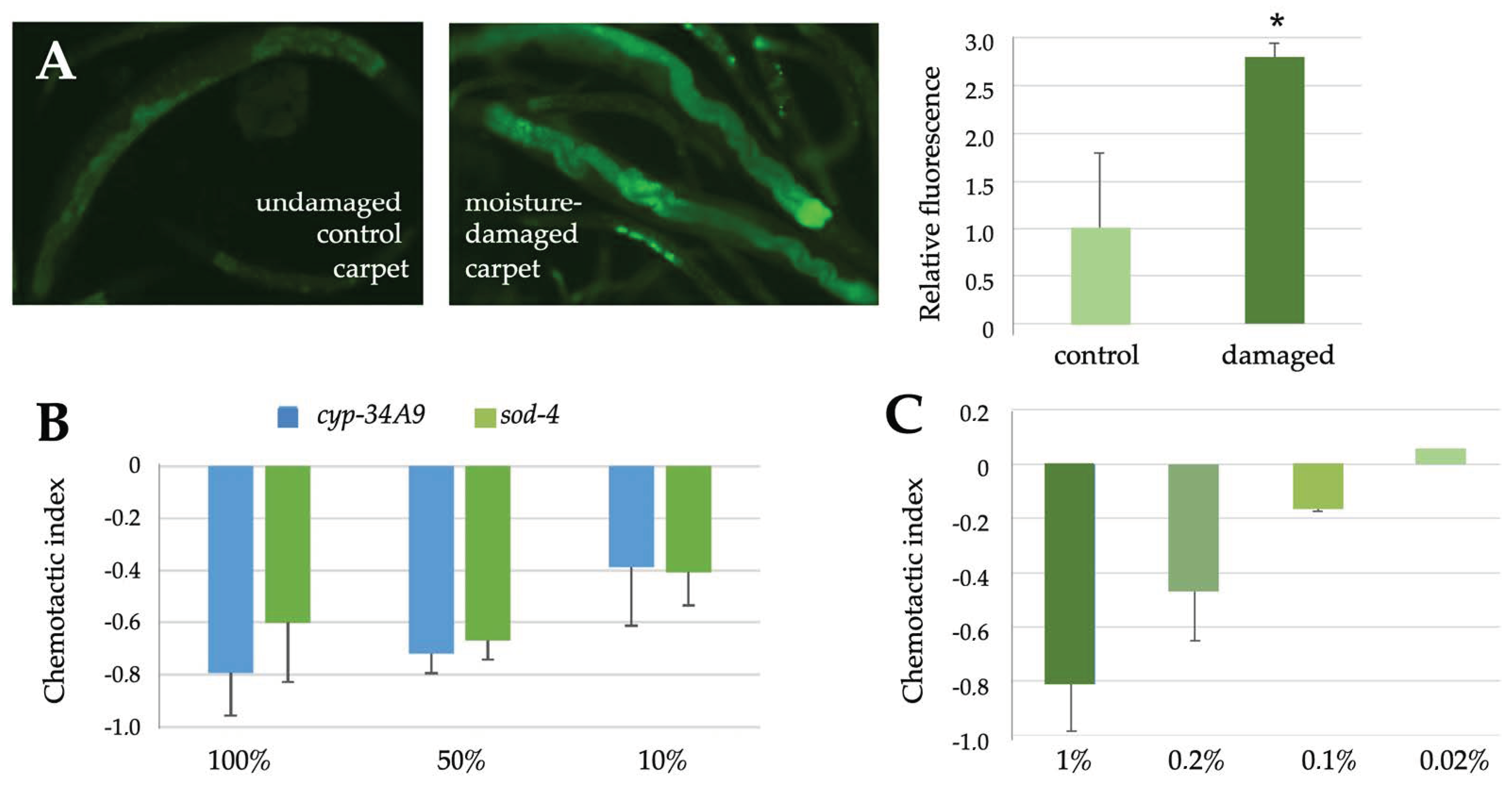
2.4. C. elegans Reporter Strains Also Respond to Airborne Exposure to Fungi
3. Discussion
4. Conclusions
5. Materials and methods
5.1. Experimental Design
5.2. C. elegans Nematode Strains and Their Maintenance
5.3. Spectrometric Analyses
5.4. Microscopic Analyses
5.5. Motility Measurements
5.6. Chemotactic Assays
5.7. In Vitro and Ex Vivo Assays
5.8. Preparation of Extracts or Collection of Liquid Exudates from Fungal Pure Cultures
5.9. Other Materials
5.10. VOC Analyses
5.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization; Regional Office for Europe. WHO Guidelines for Indoor Air Quality: Dampness and Mould. 2009. Available online: https://apps.who.int/iris/handle/10665/164348 (accessed on 22 July 2020).
- World Health Organization; Regional Office for Europe; European Centre for Environment and Health. Effects of Air Pollution on Children’s Health and Development: A Review of the Evidence. 2005. Available online: https://apps.who.int/iris/handle/10665/107652 (accessed on 22 July 2020).
- Moses, L.; Morrissey, K.; Sharpe, R.; Taylor, T. Exposure to indoor mouldy odour increases the risk of asthma in older adults living in social housing. Int. J. Environ. Res. Public Health 2019, 16, 2600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eurostat. Total Population Living in a Dwelling with a Leaking Roof, Damp Walls, Floors or Foundation, or Rot in Window Frames or Floor—EU-SILC Survey. 2020. Available online: https://ec.europa.eu/eurostat/en/web/products-datasets/-/ILC_MDHO01 (accessed on 6 January 2021).
- Dai, D.; Prussin, A.J., 2nd; Marr, L.C.; Vikesland, P.J.; Edwards, M.A.; Pruden, A. Factors shaping the human exposome in the built environment: Opportunities for engineering control. Environ. Sci. Technol. 2017, 51, 7759–7774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kembel, S.W.; Jones, E.; Kline, J.; Northcutt, D.; Stenson, J.; Womack, A.M.; Bohannan, B.J.; Brown, G.Z.; Green, J.L. Architectural design influences the diversity and structure of the built environment microbiome. ISME J. 2012, 6, 1469–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Kausar, M.A.; Singh, A.B.; Singh, R. Biological contaminants in the indoor air environment and their impacts on human health. Air Qual. Atmos. Health 2021, 14, 1723–1736. [Google Scholar] [CrossRef] [PubMed]
- Mendell, M.J.; Kumagai, K. Observation-based metrics for residential dampness and mold with dose–response relationships to health: A review. Indoor Air 2017, 27, 506–517. [Google Scholar] [CrossRef]
- Mendell, M.; Macher, J.; Kumagai, K. Measured moisture in buildings and adverse health effects: A review. Indoor Air 2018, 28, 488–499. [Google Scholar] [CrossRef]
- Miller, D.; McMullin, D. Fungal secondary metabolites as harmful indoor air contaminants: 10 years on. Appl. Microbiol. Biotechnol. 2014, 98, 9953–9966. [Google Scholar] [CrossRef]
- Holme, J.; Øya, E.; Afanou, A.; Øvrevik, W.; Eduard, W. Characterization and pro-inflammatory potential of indoor mold particles. Indoor Air 2020, 30, 662–681. [Google Scholar] [CrossRef]
- Fisk, W.; Chan, W.; Johnson, A. Does dampness and mold in schools affect health? Results of a meta-analysis. Indoor Air 2019, 29, 895–902. [Google Scholar] [CrossRef]
- Gallon, V.; Le Cann, P.; Sanchez, M.; Dematteo, C.; Le Bot, B. Emissions of VOCs, SVOCs, and mold during the construction process: Contribution to indoor air quality and future occupants’ exposure. Indoor Air 2020, 30, 691–710. [Google Scholar] [CrossRef]
- Wang, M.; Tan, G.; Eljaszewicz, A.; Meng, Y.; Wawrzyniak, P.; Acharya, S.; Altunbulakli, C.; Westermann, P.; Dreher, A.; Yan, L.; et al. Laundry detergents and detergent residue after rinsing directly disrupt tight junction barrier integrity in human bronchial epithelial cells. J. Allergy Clin. Immunol. 2019, 143, 1892–1903. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Park, E.K.; Young, T.M.; Hammock, B.D. Occurrence of endocrine-disrupting chemicals in indoor dust. Sci. Total Environ. 2008, 404, 26–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonvallot, N.; Mandin, C.; Mercier, F.; Le Bot, B.; Glorennec, P. Health ranking of ingested semi-volatile organic compounds in house dust: An application to France. Indoor Air 2010, 20, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Nazaroff, W.W.; Wescher, C. Cleaning products and air fresheners: Exposure to primary and secondary air pollutants. Atmos. Environ. 2004, 38, 2841–2865. [Google Scholar] [CrossRef]
- Inácio, A.S.; Costa, G.N.; Domingues, N.S.; Santos, M.S.; Moreno, A.J.M.; Vaz, W.L.C.; Vieira, O.V. Mitochondrial dysfunction is the focus of quaternary ammonium surfactant toxicity to mammalian epithelial cells. Antimicrob. Agents Chemother. 2013, 57, 62631–62639. [Google Scholar] [CrossRef] [Green Version]
- Dewey, H.; Jones, J.; Keating, M.; Budhathoki-Upret, J. Increased use of disinfectants during the COVID-19 pandemic and its potential impacts on health and safety. ACS Chem. Health Saf. 2022, 29, 27–38. [Google Scholar] [CrossRef]
- Anderson, S.E.; Wells, J.; Fedorowicz, A.; Butterworth, L.F.; Meade, B.; Munson, A.E. Evaluation of the contact and respiratory sensitization potential of volatile organic compounds generated by simulated indoor air chemistry. Toxicol. Sci. 2007, 97, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Carslaw, N. A mechanistic study of limonene oxidation products and pathways following cleaning activities. Atmos. Environ. 2013, 80, 507–513. [Google Scholar] [CrossRef]
- Mannerström, M.; Dvorakova, M.; Svobodova, L.; Rucki, M.; Kotal, F.; Vavrous, A.; Vrbíková, V.; Kejlova, K.; Jirova, D.; Heinonen, T. New approach methods for assessing indoor air toxicity. Curr. Res. Toxicol. 2022, 3, 100090. [Google Scholar] [CrossRef]
- Järnström, H.; Saarela, K.; Kalliokoski, P.; Pasanen, A.L. Comparison of VOC and ammonia emissions from individual PVC materials, adhesives and from complete structures. Environ. Int. 2008, 34, 420–427. [Google Scholar] [CrossRef]
- Movia, D.; Bruni-Favier, S.; Prina-Mello, A. In vitro alternatives to acute inhalation toxicity studies in animal models—A perspective. Front. Bioeng. Biotechnol. 2020, 8, 549. [Google Scholar] [CrossRef] [PubMed]
- Balls, M. Alternatives to laboratory animals: Trends in replacement and the three Rs. Altern. Lab. Anim. 2022, 50, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Atosuo, J.; Lehtinen, J.; Vojtek, L.; Lilius, E.P. Escherichia coli K-12 (pEGFPluxABCDEamp): A tool for analysis of bacterial killing by antibacterial agents and human complement activities on a real-time basis. Luminescence 2013, 28, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Hyvönen, S.M.; Lohi, J.J.; Rasanen, L.A.; Heinonen, T.; Mannerström, M.; Vaali, K.; Tuuminen, T. Association of toxic indoor air with multi-organ symptoms in pupils attending a moisture-damaged school in Finland. Am. J. Clin. Exp. Immunol. 2020, 9, 101–113. [Google Scholar]
- Nordberg, M.E.; Täubel, M.; Jalava, P.I.; BéruBé, K.; Tervahauta, A.; Hyvärinen, A.; Huttunen, K. Human airway construct model is suitable for studying transcriptome changes associated with indoor air particulate matter toxicity. Indoor Air 2020, 30, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Nordberg, M.E.; Täubel, M.; Heikkinen, S.; Jalkanen, K.; Köliö, A.; Stranger, M.; Leppänen, H.; Hyvärinen, A.; Huttunen, K. Toxicological transcriptome of human airway constructs after exposure to indoor air particulate matter: In search of relevant pathways of moisture damage-associated health effects. Environ. Int. 2022, 158, 106997. [Google Scholar] [CrossRef]
- Salin, J.; Ohtonen, P.; Syrjälä, H. Teacher’s work-related non-literature-known building-related symptoms are also connected to indoor toxicity: A cross-sectional study. Indoor Air 2021, 31, 1533–1539. [Google Scholar] [CrossRef]
- Salo, J. Tracking Diversity, Metabolic Activity, and Bioactive Metabolites of the Building Mycobiota—Examples and Novel Findings. Ph.D. Thesis, Aalto University, Helsinki, Finland, 2022. [Google Scholar]
- Tuuminen, T.; Andersson, A.; Hyvönen, S.; Lohi, J.; Vaali, K. Indoor air non-toxicity should be proven with special techniques prior claiming that it may cause a variety of mental disorders. J. Hyg. Environ. Health 2020, 229, 113545. [Google Scholar] [CrossRef]
- Nordin, S. Mechanisms underlying nontoxic indoor air health problems: A review. Int. J. Hyg. Environ. Health 2020, 226, 113489. [Google Scholar] [CrossRef]
- Candido, E.P.M.; Jones, D. Transgenic Caenorhabditis elegans strains as biosensors. Trends Biotechnol. 1996, 14, 125. [Google Scholar]
- Bargmann, C.I.; Hartwieg, E.; Horvitz, H.R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 1993, 74, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Bargmann, C.I. Chemosensation in C. elegans. WormBook 2006, 25, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Anbalagan, C.; Lafayette, I.; Antoniu-Kourounioti, M.; Haque, M.; King, J.; Baillie, J.; Gurierrez, C.; Rodriguez Martin, J.A.; De Pomerai, D. Transgenic nematodes as biosensors for metal stress in soil pore water samples. Ecotoxicology 2012, 21, 439–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbalagan, C.; Lafayette, I.; Antoniu-Kourounioti, M.; Gurierrez, C.; Martin, J.A.; Chowdhuri, D.K.; De Pomerai, D.I. Use of transgenic GFP reporter strains of the nematode Caenorhabditis elegans to investigate the patterns of stress responses induced by pesticides and by organic extracts from agricultural soils. Ecotoxicology 2013, 22, 72–85. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.E.; Lewis, C.W.; Anderson, J.G.; Solomons, G.L. Mycotoxins in Human Health; Report EUR 16048 EN; European Commission, Directorate-General XII: Brussels, Belgium, 1994. [Google Scholar]
- Bullerman, L.B.; Bianchini, A. Stability of mycotoxins during food processing. Int. J. Food Microbiol. 2007, 119, 140–146. [Google Scholar] [CrossRef]
- Fogle, M.R.; Douglas, D.R.; Jumper, C.A.; Straus, D.C. Heat stability of chaetoglobosins A and C. Can. J. Microbiol. 2008, 54, 423–425. [Google Scholar] [CrossRef]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [Green Version]
- Castagnoli, E.; Salo, J.; Toivonen, M.S.; Marik, T.; Mikkola, R.; Kredics, L.; Vicente-Carrillo, A.; Nagy, S.; Andersson, M.T.; Andersson, M.A.; et al. An evaluation of boar spermatozoa as a biosensor for the detection of sublethal and lethal toxicity. Toxins 2018, 10, 463. [Google Scholar] [CrossRef] [Green Version]
- Gareis, M.; Gareis, E.M. Guttation droplets of Penicillium nordicum and Penicillium verrucosum contain high concentrations of the mycotoxins ochratoxin A and B. Mycopathologia 2007, 163, 207–214. [Google Scholar] [CrossRef]
- Salo, M.J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A, B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Nagy, S.; Mikkola, R.; Kurnitski, J.; Salonen, H. Toxicities of detergents used in cleaning chemicals and hygiene products in a test battery of ex vivo and in vitro assays. In Sisäilmastoseminaari; SIY Report; Sisäilmayhdistys Ry: Espoo, Finland, 2019; Volume 37, pp. 297–302. Available online: https://www.sisailmauutiset.fi/Sisailmastoseminaari_2019.pdf (accessed on 15 January 2023).
- Valvira, Finnish National Supervisory Authority for Welfare and Health. Asumisterveysasetuksen Soveltamisohje Osa III, §15, Ohje 8/2016. Available online: https://www.valvira.fi/documents/14444/261239/Asumisterveysasetuksen+soveltamisohje+osa+III.pdf/997eeca1-53f7-4d4e-bb7a-df6ef7ee0e9c (accessed on 16 December 2022). (In Finnish).
- Andersen, B.; Frisvad, J.C.; Søondergaard, I.; Rasmussen, I.S.; Larsen, L.S. Associations between fungal species and water-damaged building materials. Appl. Environ. Microbiol. 2011, 77, 4180–4188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valvira, Finnish National Supervisory Authority for Welfare and Health. Asumisterveysasetuksen Soveltamisohje Osa IV, §20, Ohje 8/2016. Available online: https://www.valvira.fi/documents/14444/261239/Asumisterveysasetuksen+soveltamisohje+osa+IV.pdf/cdfaaa39-d2e5-4bd6-b9e9-6d9c0f60bff6 (accessed on 16 December 2022). (In Finnish).
- Horner, W.E.; Worthan, A.G.; Morey, P.R. Air- and dustborne mycoflora in houses free of water damage and fungal growth. Appl. Environ. Microbiol. 2004, 70, 6394–6400. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, S.; Schäfer, C. Toxin production by Stachybotrys chartarum genotype S on different culture media. J. Fungi 2020, 6, 159. [Google Scholar] [CrossRef] [PubMed]
- Wargenau, A.; Schulz, S.; Hakimzadeh, A.; Tufenkji, N. Evaluating the cell membrane penetration potential of lipid-soluble compounds using supported phospholipid bilayers. Anal. Chem. 2018, 90, 11174–11178. [Google Scholar] [CrossRef] [Green Version]
- Popova, A.A.; Koksharova, O.A.; Lipasova, V.A.; Zaitseva, J.V.; Katkova-Zhukotskaya, O.A.; Eremina, S.; Mironov, A.; Chernin, L.S.; Khmel, I.A. Inhibitory and toxic effects of volatiles emitted by strains of Pseudomonas and Serratia on growth and survival of selected micro-organisms, Caenorhabditis elegans and Drosophila melanogaster. BioMed Res. Int. 2014, 2014, 125704. [Google Scholar] [CrossRef] [Green Version]
- EU LCI Working Group. 2021. Available online: https://ec.europa.eu/docsroom/documents/49239 (accessed on 17 December 2022).
- Nørgaard, A.W.; Kudal, J.D.; Koefoed-Sørensen, V.; Koponen, J.K.; Wolkoff, P. Ozone-initiated VOC and particle emissions from a cleaning agent and an air freshener: Risk assessment of acute airway effects. Environ. Int. 2014, 68, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Salo, J.; Kedves, O.; Kredics, L.; Druzhinina, I.; Kurnitski, J.; Salonen, H. Bioreactivity, guttation and agents influencing surface tension of water emitted by actively growing indoor mould isolates. Microorganisms 2020, 8, 1940. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Kalichamy, K.S.; Ikkala, K.; Pörsti, J.; Santio, N.M.; Tuomaala, J.; Jha, S.; Holmberg, C.I.; Koskinen, P.J. PIM-related kinases selectively regulate olfactory sensations in Caenorhabditis elegans. eNeuro 2019, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Mikkola, R.; Rasimus, S.; Hoornstra, D.; Salin, P.; Rahkila, R.; Heikkinen, M.; Mattila, S.; Peltola, J.; Kalso, S.; et al. Boar spermatozoa as a biosensor for detecting toxic substances in indoor dust and aerosols. Toxicol. Vitr. 2010, 24, 2041–2052. [Google Scholar] [CrossRef]
- Mikkola, R.; Andersson, M.A.; Kredics, L.; Grigoriev, P.A.; Sundell, N.; Salkinoja-Salonen, M.S. 20-residue and 11-residue peptaibols from the fungus Trichoderma longibrachiatum are synergistic in forming Na+/K+-permeable channels and adverse action towards mammalian cells. FEBS J. 2012, 279, 4172–4190. [Google Scholar] [CrossRef] [PubMed]
- Salo, J.; Kedves, O.; Mikkola, R.; Kredics, L.; Andersson, M.; Kurnitski, J.; Salonen, H. Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the diversity tracking of mycotoxin-producing Chaetomium-like isolates obtained in buildings in Finland. Toxins 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2019; Volume 2, p. 481. [Google Scholar]
- Kawai, K.; Nozawa, Y.; Maebayashi, Y.; Yamazaki, M.; Hamasaki, T. Averufin, an anthraquinone mycotoxin possessing a potent uncoupling effect on mitochondrial respiration. Appl. Environ. Microbiol. 1984, 47, 481–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewangan, J.; Srivastava, S.; Mishra, S.; Pandey, P.K.; Divakar, A.; Rath, S.K. Chetomin induces apoptosis in human triple-negative breast cancer cells by promoting calcium overload and mitochondrial dysfunction. Biochem. Biophys. Res. Commun. 2018, 495, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Min, S.; Wang, X.; Du, Q.; Gong, H.; Yang, Y.; Wang, T.; Wu, N.; Liu, X.; Li, W.; Zhao, C.; et al. Chetomin, a Hsp90/HIF1α pathway inhibitor, effectively targets lung cancer stem cells and nonstem cells. Cancer Biol. Ther. 2020, 21, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Choi, G.J.; Jang, K.S.; Lim, H.K.; Kim, H.T.; Cho, K.Y.; Kim, J.C. Antifungal activity against plant pathogenic fungi of chaetoviridins isolated from Chaetomium globosum. FEMS Microbiol. Lett. 2005, 252, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Mikkola, R.; Andersson, M.A.; Hautaniemi, M.; Salkinoja-Salonen, M.S. Toxic indole alkaloids avrainvillamide and stephacidin B produced by a biocide tolerant indoor mold Aspergillus westerdijkiae. Toxicon 2015, 99, 58–67. [Google Scholar] [CrossRef]
- Zingales, V.; Fernández-Franzón, M.; Ruiz, M.J. Sterigmatocystin: Occurrence, toxicity and molecular mechanisms of action—A review. Food Chem. Toxicol. 2020, 146, 111802. [Google Scholar] [CrossRef]
- Andersson, M.A.; Salo, J.; Mikkola, R.; Marik, T.; Kredics, L.; Kurnitski, J.; Salonen, H. Melinacidin-producing Acrostalagmus luteoalbus, a major constituent of mixed mycobiota contaminating insulation material in an outdoor wall. Pathogens 2021, 10, 843. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models/Toxicity of Exposure Agents | Strongly Toxic +++ | Moderately Toxic ++ | Mildly Toxic + | Nontoxic - |
|---|---|---|---|---|
| Relative increse in Ce fluorescence | >2 x | 1.5–2 x | 1.25–1.5 x | <1.25 x |
| Decrease in cell proliferation (EC50) | <5 µg/mL | 5–15 µg/mL | 15–50 µg/mL | >50 µg/mL |
| Decrease in sperm motility (EC50) | <5 µg/mL | 5–10 µg/mL | 10–25 µg/mL | >25 µg/mL |
| Strain Code | Species/Genus | Mycotoxins | C. elegans | Cells | Sperm |
|---|---|---|---|---|---|
| POB8 | Agrostalagmus luteoalbus1 | melinacidins | + | ++ | ++ |
| K20 | Aspergillus versicolor1 | sterigmatocystin | ++ | +++ | ++ |
| SL3 | Aspergillus versicolor2 | sterigmatocystin, averufin | + | +++ | ++ |
| MH1 | Chaetomium1 | chaetoglobosin | +++ | + | ++ |
| OT7 | Chaetomium cochliodes1 | chaetomin, chaetoviridins | ++ | +++ | ++ |
| ABCD | Chaetomium globosum1 | chaetoglobosin, chaetoviridins | +++ | + | ++ |
| MH5 | Chaetomium globosum1 | chaetoglobosin, chaetoviridins | +++ | + | ++ |
| TR | Penicillium sp. 1 | nontoxic | - | - | - |
| RcP61 | Penicillium expansum2 | chaetoglobosin, communesins | +++ | +++ | +++ |
| SJ40 | Trichoderma citrinoviride1 | trilongins | - | + | +++ |
| THG | Trichoderma longibrachiatum1 | trilongins | + | + | +++ |
| NJ14 | Trichoderma trixiae2 | ND | + | ++ | +++ |
| Strain Code | Species/Genus | Ce sod-4 | Ce cyp-34A9 | Cells | Sperm |
|---|---|---|---|---|---|
| A21 | Acrenium sp. | + | + | - | ++ |
| BA36 | Aspergillus sp. | ++ | +++ | ++ | +++ |
| A4 | Aureobasidium pullulans | ++ | ++ | ++ | + |
| A5 | Aureobasidium pullulans | - | - | ++ | + |
| A6 | Aureobasidium pullulans | - | ND | - | - |
| BA35 | Aureobasidium pullulans | + | + | + | - |
| A3 | Exophiala sp. | - | - | -/+ | - |
| BA32 | Exophiala sp. | ++ | ND | ++ | - |
| L6 | Paecilomyces sp. | - | - | - | - |
| L21 | Paecilomyces sp. | - | ND | -/+ | - |
| BA37 | Rhizomucor sp. | - | ND | - | - |
| Chemical Compound | Detergent Type | Ce sod-4 Fluorescence | Ce cyp-34A9 Fluorescence | Ce Motility | Cell Proliferation | Sperm Motility |
|---|---|---|---|---|---|---|
| DDDAC | cationic | +++ | ++ | +++ | +++ | +++ |
| Genapol X-080 | non-ionic | ++ | ++ | + | + | +++ |
| Triton X-100 | non-ionic | +++ | +++ | ++ | + | +++ |
| Tween 80 | non-ionic | - | - | ++ | - | ++ |
| SDS | anionic | - | - | - | - | ++ |
| Strain Code | Species/Genus | Ce Fluorescence |
|---|---|---|
| a/464 | Aspergillus niger | ++ |
| Asp25 | Aspergillus ochraeus | +++ |
| a/459 | Chaetomium sp. | + |
| b/548 | Geotrichium candidum | - |
| a/462 | Paecilomyces variotii | + |
| a/467 | Stachybotrys sp. | +++ |
| a/465 | Trichoderma sp. | + |
| a/456 | Wallemia sp. | ++ |
| VOCs/Chamber | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| TVOCs | 1500 | 160 | 560 | 550 |
| 1-butanol | 19 | 3 | - | 6 |
| 2-ethyl-1-hexanol | 830 | 240 | 460 | 770 |
| C9/C10 alcohols | 880 | - | 220 | - |
| a-pinene | 62 | - | 15 | - |
| ß-pinene | 13 | - | - | - |
| DMPCS | 6 | - | - | 11 |
| toluene | - | 3 | - | 5 |
| PMH | - | - | 17 | - |
| 2-ethylhexanal | - | - | 8 | 10 |
| 3-heptanone | - | - | 10 | 7 |
| cyclohexanone | - | - | - | 3 |
| butyl ether | - | - | - | 33 |
| Strain Code | Species/Genus | Origin of Strain | Medium |
|---|---|---|---|
| a/466 | Aspergillus fumigatus1 | unknown | MEA |
| a/464 | Aspergillus niger1,2 | unknown | MEA |
| Asp25 | Aspergillus ochraeus1,2 | timber | MEA |
| a/459 | Chaetomium sp. 1,2 | chipboard/plastic carpet | MEA |
| b/548 | Geotrichium candidum1,2 | negative control | MEA |
| a/462 | Paecilomyces variotii1,2 | unknown | MEA |
| a/452 | Stachybotrys sp. 1 | toja board | MEA |
| a/453 | Stachybotrys sp. 1 | toja board | MEA |
| a/467 | Stachybotrys sp. 1,2 | timber | MEA |
| a/468 | Stachybotrys sp. 1 | mineral wool | MEA |
| a/470 | Stachybotrys sp. 1 | mineral wool | MEA |
| a/465 | Trichoderma sp. 1,2 | plastic paste | MEA |
| a/456 | Wallemia sp. 2 | mineral wool | DG18 |
| Strain Code and Reference | Species/Genus | Origin of Strain |
|---|---|---|
| POB8 [45] | Agrostalagmus luteoalbus1 | cork liner, Espoo |
| K20 [45] | Aspergillus versicolor1 | settled dust, Espoo |
| SL3 [45] | Aspergillus versicolor2 | fall out plate, Helsinki |
| MH1 [45] | Chaetomium 1 | settled dust, Helsinki |
| OT7 = SzMC 24764 [61] | Chaetomium cochliodes1 | settled dust, Helsinki |
| ABCD [61] | Chaetomium globosum1 | settled dust, Helsinki |
| MH5 = SzMC 24456 [61] | Chaetomium globosum1 | settled dust, Espoo |
| TR [42] | Penicillium sp. 1 | indoor air |
| RcP61 [45] | Penicillium expansum2 | cork liner, Espoo |
| SJ40 [42] | Trichoderma citrinoviride1 | settled dust, Espoo |
| THG = SzMC Thg [60] | Trichoderma longibrachiatum1 | insulation, Oulu |
| NJ14 [42] | Trichoderma trixiae2 | settled dust, Nivala |
| Strain Code | Species/Genus | Origin of Strain | Medium |
|---|---|---|---|
| A21 | Acremonium sp. | surface dust, Vantaa | MEA |
| BA36 | Aspergillus section Aspergillus | filter from exhaust air channel, Oulu | DG18 |
| A4 | Aureobasidium pullulans | surface dust, Vantaa | DG18 |
| A5 | Aureobasidium pullulans | surface dust, Vantaa | DG18 |
| A6 | Aureobasidium pullulans | surface dust, Vantaa | DG18 |
| BA35 | Aureobasidium pullulans | filter from exhaust air channel, Oulu | DG18 |
| A3 | Exophiala sp. | surface dust, Vantaa | MEA |
| BA32 | Exophiala sp. | filter from exhaust air channel, Oulu | MEA |
| L6 | Paecilomyces sp. | insulation, Jyväskylä | MEA |
| L21 | Paecilomyces sp. | insulation, Jyväskylä | MEA |
| BA37 | Rhizomucor sp. | filter from exhaust air channel, Oulu | MEA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paavanen-Huhtala, S.; Kalichamy, K.; Pessi, A.-M.; Häkkilä, S.; Saarto, A.; Tuomela, M.; Andersson, M.A.; Koskinen, P.J. Biomonitoring of Indoor Air Fungal or Chemical Toxins with Caenorhabditis elegans nematodes. Pathogens 2023, 12, 161. https://doi.org/10.3390/pathogens12020161
Paavanen-Huhtala S, Kalichamy K, Pessi A-M, Häkkilä S, Saarto A, Tuomela M, Andersson MA, Koskinen PJ. Biomonitoring of Indoor Air Fungal or Chemical Toxins with Caenorhabditis elegans nematodes. Pathogens. 2023; 12(2):161. https://doi.org/10.3390/pathogens12020161
Chicago/Turabian StylePaavanen-Huhtala, Sari, Karunambigai Kalichamy, Anna-Mari Pessi, Sirkku Häkkilä, Annika Saarto, Marja Tuomela, Maria A. Andersson, and Päivi J. Koskinen. 2023. "Biomonitoring of Indoor Air Fungal or Chemical Toxins with Caenorhabditis elegans nematodes" Pathogens 12, no. 2: 161. https://doi.org/10.3390/pathogens12020161