
Anisakid Presence in the European Conger, Conger conger, from Spanish Mediterranean Waters †
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Host Sampling and Parasite Collection
2.2. Host Diet
2.3. Epidemiological Concepts and Statistical Analysis
3. Results
3.1. Host Parasitic Infection
3.2. Host Prey Items
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tortonese, E. Fauna d’Italia. Osteichthyes (Pesci Ossei); Calderini: Bologna, Italy, 1970. [Google Scholar]
- Santos, R.V.S.; Silva, W.M.M.L.; Novoa-Pabon, A.M.; Silva, H.M.; Pinho, M.R. Long-Term Changes in the Diversity, Abundance and Size Composition of Deep Sea Demersal Teleosts from the Azores Assessed through Surveys and Commercial Landings. Aquat. Living Resour. 2019, 32, 25. [Google Scholar] [CrossRef]
- Casadevall, M.; Sarrà-Alarcón, L.; Delgado, E.; Matallanas, J. The Sexual Segregation of the European Eel, Conger conger (Linnaeus, 1758) (Anguilliformes, Congridae) and Female Semelparity in the North-West Mediterranean. J. Fish. Res. 2017, 1, 5–14. [Google Scholar]
- Xavier, J.C.; Cherel, Y.; Assis, C.A.; Sendão, J.; Borges, T.C. Feeding Ecology of Conger Eels (Conger conger) in North-East Atlantic Waters. J. Mar. Biol. Assoc. UK 2010, 90, 493–501. [Google Scholar] [CrossRef]
- Cau, A.; Manconi, P. Relationship of Feeding, Reproductive Cycle and Bathymetric Distribution in Conger conger. Mar. Biol. 1984, 81, 147–151. [Google Scholar] [CrossRef]
- Bauchot, M.L.; Saldanha, L. Congridae. In Fishes of the Northeastern Atlantic and Mediterranean; Whitehead, J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; Volume 2, pp. 567–574. [Google Scholar]
- Santos, R.; Medeiros-Leal, W.; Pinho, M. Synopsis of Biological, Ecological and Fisheries-Related Information on Priority Marine Species in the Azores Region. Arquipelago 2020, 1, 1–138. [Google Scholar] [CrossRef]
- FAO Fisheries and Aquaculture. Available online: https://www.fao.org/fishery/en/aqspecies/2994/en (accessed on 2 October 2023).
- Medeiros-Leal, W.; Santos, R.; Novoa-Pabon, A.; Silva, H.; Pinho, M. Population Structure of the European Conger Conger conger from the Mid-North Atlantic Ocean Inferred from Bathymetric Distribution, Length Composition and Movement Patterns Analyses. Fish. Manag. Ecol. 2021, 28, 468–477. [Google Scholar] [CrossRef]
- Piñeiro, C.G.; Casas, M.; Bañón, R. The Deep-Water Fisheries Exploited by Spanish Fleets in the Northeast Atlantic: A Review of the Current Status. Fish. Res. 2001, 51, 311–320. [Google Scholar] [CrossRef]
- Saraiva, A.; Cruz, C.; Santos, M.J.; Ferreira, S. Nematode Parasites from Digestive Tract of Conger Conger conger L. from the Northwest Coast of the Iberian Peninsula. Bull. Eur. Fish. Pathol. 2000, 20, 163–166. [Google Scholar]
- Costa, G.; Santos, M.J.; Costa, L.; Biscoito, M.; Pinheiro De Carvalho, M.A.A.; Melo-Moreira, E. Helminth Parasites from the Stomach of Conger Eel, Conger conger, from Madeira Island, Atlantic Ocean. J. Parasitol. 2009, 95, 1013–1015. [Google Scholar] [CrossRef]
- Bartoli, P.; Bray, R.A.; Gibson, D.I. Opecoelidae (Digenea) from Western Mediterranean Fishes: Three Rare Species. Syst. Parasitol. 2003, 55, 81–95. [Google Scholar] [CrossRef]
- Culurgioni, J.; D’Amico, V.; Coluccia, E.; Mulas, A.; Figus, V. Metazoan Parasite Fauna of Conger Eel Conger conger L. from Sardinian Waters (Italy). Ittiopatologia 2006, 3, 253–261. [Google Scholar]
- Sanmartin Duran, M.; Quinteiro, P.; Ubeira, F. Nematode Parasites of Commercially Important Fish in NW Spain. Dis. Aquat. Organ. 1989, 7, 75–77. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards. Scientific Opinion on Risk Assessment of Parasites in Fishery Products. EFSA J. 2010, 8, 1–91. [Google Scholar] [CrossRef]
- Audícana, M.T.; Ansotegui, I.J.; Fernández de Corres, L.; Kennedy, M.W. Anisakis simplex: Dangerous Dead and Alive? Trends Parasitol. 2002, 18, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; D’Amelio, S. Anisakiasis. In Helminth Infections and Their Impact on Global Public Health; Bruschi, F., Ed.; Springer-Verlag: Viena, Austria, 2014; pp. 325–365. [Google Scholar]
- Berland, B. Nematodes from Some Norwegian Marine Fishes. Sarsia 1961, 2, 1–50. [Google Scholar] [CrossRef]
- Hartwich, G. Keys to Genera of the Ascaridoidea. In CIH Keys to the Nematode Parasites of Vertebrates; Anderson, R.C., Chabaud, A.G., Willmott, S., Eds.; Commonwealth Agricultural Bureaux: Slough, UK, 1974; Volume 2, pp. 1–15. [Google Scholar]
- Petter, A.J. Quelques Spirurides de Poissons de La Région Nantaise. Ann. Parasitol. Hum. Comp. 1970, 45, 31–46. [Google Scholar] [CrossRef]
- Quinteiro, P.; Fernández, J.A.; Sanmartín, M.L. First Description of the Male and New Data on the Female of Cristitectus congeri Petter, 1970 (Nematoda: Cystidicolidae). Syst. Parasitol. 1989, 14, 37–41. [Google Scholar] [CrossRef]
- Zariquiey Alvarez, R. Crustáceos Decápodos Ibéricos. Invest. Pesq. 1968, 32, 1–510. [Google Scholar]
- Fischer, W.; Schneider, M.; Bauchot, M.-L. Fiches FAO d’Identification des Espèces pour Les Besoins de La Pêche: Méditerranée et Mer Noir Zone de Pêche 37; Organisation des Nations Unies pour l’Alimentation et l’Agriculture: Rome, Italy, 1987. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying Parasites in Samples of Hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Alburqueque, R.A.; Palomba, M.; Santoro, M.; Mattiucci, S. Molecular Identification of Zoonotic Parasites of the Genus Anisakis (Nematoda: Anisakidae) from Fish of the Southeastern Pacific Ocean (off Peru Coast). Pathogens 2020, 9, 910. [Google Scholar] [CrossRef] [PubMed]
- Palomba, M.; Mattiucci, S.; Crocetta, F.; Osca, D.; Santoro, M. Insights into the Role of Deep-Sea Squids of the Genus Histioteuthis (Histioteuthidae) in the Life Cycle of Ascaridoid Parasites in the Central Mediterranean Sea Waters. Sci. Rep. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Nascetti, G. Advances and Trends in the Molecular Systematics of Anisakid Nematodes, with Implications for Their Evolutionary Ecology and Host-Parasite Co-Evolutionary Processes. Adv. Parasitol. 2008, 66, 47–148. [Google Scholar] [CrossRef] [PubMed]
- Roca-Geronès, X.; Montoliu, I.; Godínez-González, C.; Fisa, R.; Shamsi, S. Morphological and Genetic Characterization of Hysterothylacium Ward & Magath, 1917 (Nematoda: Raphidascarididae) Larvae in Horse Mackerel, Blue Whiting and Anchovy from Spanish Atlantic and Mediterranean Waters. J. Fish Dis. 2018, 41, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Moravec, F.; Justine, J. Lou New Records of Cucullanid Nematodes from Marine Fishes off New Caledonia, with Descriptions of Five New Species of Cucullanus (Nematoda, Cucullanidae). Parasite 2020, 27, 1–23. [Google Scholar] [CrossRef]
- Roca-Geronès, X.; Segovia, M.; Godínez-González, C.; Fisa, R.; Montoliu, I. Anisakis and Hysterothylacium Species in Mediterranean and North-East Atlantic Fishes Commonly Consumed in Spain: Epidemiological, Molecular and Morphometric Discriminant Analysis. Int. J. Food Microbiol. 2020, 325, 108642. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular Epidemiology of Anisakis and Anisakiasis: An Ecological and Evolutionary Road Map. Adv. Parasito.l 2018, 99, 93–263. [Google Scholar] [CrossRef]
- Piras, M.C.; Tedde, T.; Garippa, G.; Virgilio, S.; Sanna, D.; Farjallah, S.; Merella, P. Molecular and Epidemiological Data on Anisakis Spp. (Nematoda: Anisakidae) in Commercial Fish Caught off Northern Sardinia (Western Mediterranean Sea). Vet. Parasitol. 2014, 203, 237–240. [Google Scholar] [CrossRef]
- Muñoz, M.V.; Fernández, J.P.; Orts, M.E.; Carbonell, E. Sobre Algunos Nematodos Cucullanidos Parásitos de Conger conger (L.) En Aguas Del Litoral Valenciano. Rev. Iber. Parasitol. 1988, 48, 275–281. [Google Scholar]
- Cuéllar, M.C.; Fontanilla, J.C.; Pérez-Fuentes, J.; Pérez-Tauler, M.P. Biología y Epidemiología de La Anisakidosis Larvaria. Enfermedad Arenque. Cienc. Vet. 1991, 4, 57–63. [Google Scholar]
- Barcala, E.; Ramilo, A.; Ortega, N.; Picó, G.; Abollo, E.; Pascual, S.; Muñoz, P. Occurrence of Anisakis and Hysterothylacium Larvae in Commercial Fish from Balearic Sea (Western Mediterranean Sea). Parasitol. Res. 2018, 117, 4003–4012. [Google Scholar] [CrossRef] [PubMed]
- Molina-Fernández, D.; Rubio-Calvo, D.; Adroher, F.J.; Benítez, R. Molecular Epidemiology of Anisakis Spp. in Blue Whiting Micromesistius poutassou in Eastern Waters of Spain, Western Mediterranean Sea. Int. J. Food. Microbiol. 2018, 282, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Akmirza, A. Metazoan Parasite Fauna of Conger Eel (Conger conger L.) near Gökçeada, Northeasten Aegean Sea, Turkey. Kafkas Univ. Vet. Fak. Derg. 2012, 18, 845–848. [Google Scholar] [CrossRef]
- Sanmartín, M.L.; Alvarez, M.F.; Peris, D.; Iglesias, R.; Leiro, J. Helminth Parasite Communities of the Conger Eel in the Estuaries of Arousa and Muros (Galicia, North-West Spain). J. Fish Biol. 2000, 57, 1122–1133. [Google Scholar] [CrossRef]
- Santos, M.J.; Castro, R.; Cavaleiro, F.; Rangel, L.; Palm, H.W. Comparison of Anisakid Infection Levels between Two of Atlantic Mackerel (Scomber colias and S. scombrus) off the Atlantic Portuguese Coast. Sci. Mar. 2017, 81, 179–185. [Google Scholar] [CrossRef]
- Saldanha, L.; Almeida, A.J.; Andrade, F.; Guerreiro, J. Observations on the Diet of Some Slope Dwelling Fishes of Southern Portugal. Int. Rev. Gesamten. Hydrobiol. 1995, 80, 217–234. [Google Scholar] [CrossRef]
- Morato, T.; Solà, E.; Grós, M.P.; Menezes, G. Diets of Forkbeard (Phycis phycis) and Conger Eel (Conger conger) off the Azores during Spring of 1996 and 1997. Arquipelago 1999, 17, 51–64. [Google Scholar]
- Matić-Skoko, S.; Ferri, J.; Tutman, P.; Skaramuca, D.; Dikić, D.; Lisičić, D.; Franić, Z.; Skaramuca, B. The Age, Growth and Feeding Habits of the European Conger Eel, Conger conger (L.) in the Adriatic Sea. Mar. Biol. Res. 2012, 8, 1012–1018. [Google Scholar] [CrossRef]
- Madrid, E.; Galán-Puchades, M.T.; Fuentes, M.V. Risk Analysis of Human Anisakidosis Through the Consumption of the Blue Whiting, Micromesistius poutassou, Sold at Spanish Supermarkets. Foodborne Pathog. Dis. 2012, 9, 934–938. [Google Scholar] [CrossRef]
- Osanz-Mur, A.C. Presencia de Larvas de Anisákidos (Nematoda: Ascaridoidea) en Pescado de Consumo Capturado en la Zona Pesquera de Tarragona. Ph.D. Thesis, Universitat Autonoma de Barcelona, Bellaterra, Spain, 2001. [Google Scholar]
- Rello, F.J.; Adroher, F.J.; Valero, A. Hysterothylacium aduncum, the Only Anisakid Parasite of Sardines (Sardina pilchardus) from the Southern and Eastern Coasts of Spain. Parasitol. Res. 2008, 104, 117–121. [Google Scholar] [CrossRef]
- Gutiérrez-Galindo, J.F.; Osanz-Mur, A.C.; Mora-Ventura, M.T. Occurrence and Infection Dynamics of Anisakid Larvae in Scomber scombrus, Trachurus trachurus, Sardina pilchardus, and Engraulis encrasicolus from Tarragona (NE Spain). Food Control 2010, 21, 1550–1555. [Google Scholar] [CrossRef]
- Herrador, Z.; Daschner, Á.; Perteguer, M.J.; Benito, A. Epidemiological Scenario of Anisakidosis in Spain Based on Associated Hospitalizations: The Tip of the Iceberg. Clin. Inf. Dis. 2019, 69, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Roca-Geronès, X.; Alcover, M.M.; Godínez-González, C.; González-Moreno, O.; Masachs, M.; Fisa, R.; Montoliu, I. First Molecular Diagnosis of Clinical Cases of Gastric in Spain. Genes 2020, 11, 452. [Google Scholar] [CrossRef] [PubMed]
- López-Serrano, M.C.; Gomez, A.A.; Daschner, Á.; Moreno-Ancillo, Á.; Suarez de Parga, J.M.; Caballero, M.T.; Barranco, P.; Cabañas, R. Gastroallergic Anisakiasis: Findings in 22 Patients. J. Gastroenterol. Hepatol. 2000, 15, 503–506. [Google Scholar] [CrossRef] [PubMed]
- González Quijada, S.; González Escudero, R.; Arias García, L.; Gil Martín, A.R.; Vicente Serrano, J.; Corral Fernández, E. Manifestaciones Digestivas de La Anisakiasis: Descripción de 42 Casos. Rev. Clin. Esp. 2005, 205, 311–315. [Google Scholar] [CrossRef]
- Klapper, R.; Kuhn, T.; Münster, J.; Levsen, A.; Karl, H.; Klimpel, S. Anisakid Nematodes in Beaked Redfish (Sebastes mentella) from Three Fishing Grounds in the North Atlantic, with Special Notes on Distribution in the Fish Musculature. Vet. Parasitol. 2015, 207, 72–80. [Google Scholar] [CrossRef]
- Levsen, A.; Karl, H. Anisakis simplex (s.l.) in Grey Gurnard (Eutrigla gurnardus) from the North Sea: Food Safety Considerations in Relation to Fishing Ground and Distribution in the Flesh. Food Control 2014, 36, 15–19. [Google Scholar] [CrossRef]
- Choi, S.J.; Lee, J.C.; Kim, M.J.; Hur, G.Y.; Shin, S.Y.; Park, H.S. The Clinical Characteristics of Anisakis Allergy in Korea. Korean J. Intern. Med. 2009, 24, 160–163. [Google Scholar] [CrossRef]
- Sohn, W.M.; Na, B.K.; Kim, T.H.; Park, T.J. Anisakiasis: Report of 15 Gastric Cases Caused by Anisakis Type I Larvae and a Brief Review of Korean Anisakiasis Cases. Korean J. Parasitol. 2015, 53, 465–470. [Google Scholar] [CrossRef]
- Chen, H.X.; Zhang, L.P.; Gibson, D.I.; Lü, L.; Xu, Z.; Li, H.T.; Ju, H.D.; Li, L. Detection of Ascaridoid Nematode Parasites in the Important Marine Food-Fish Conger myriaster (Brevoort) (Anguilliformes: Congridae) from the Zhoushan Fishery, China. Parasit Vectors 2018, 11, 274. [Google Scholar] [CrossRef]
- Cipriani, P.; Sbaraglia, G.L.; Paoletti, M.; Giulietti, L.; Bellisario, B.; Palomba, M.; Bušelić, I.; Mladineo, I.; Nascetti, G.; Mattiucci, S. The Mediterranean European Hake, Merluccius merluccius: Detecting Drivers Influencing the Anisakis Spp. Larvae Distribution. Fish Res. 2018, 202, 79–89. [Google Scholar] [CrossRef]
- Audicana, M.T.; Kennedy, M.W. Anisakis simplex: From Obscure Infectious Worm to Inducer of Immune Hypersensitivity. Clin. Microbiol. Rev. 2008, 21, 360–379. [Google Scholar] [CrossRef]
- Mattiucci, S.; Palomba, M.; Nascetti, G. Anisakis. In Encyclopedia of Infection and Immunity; Elsevier: Amsterdam, The Netherlands, 2022; Volume 2, pp. 408–423. [Google Scholar]


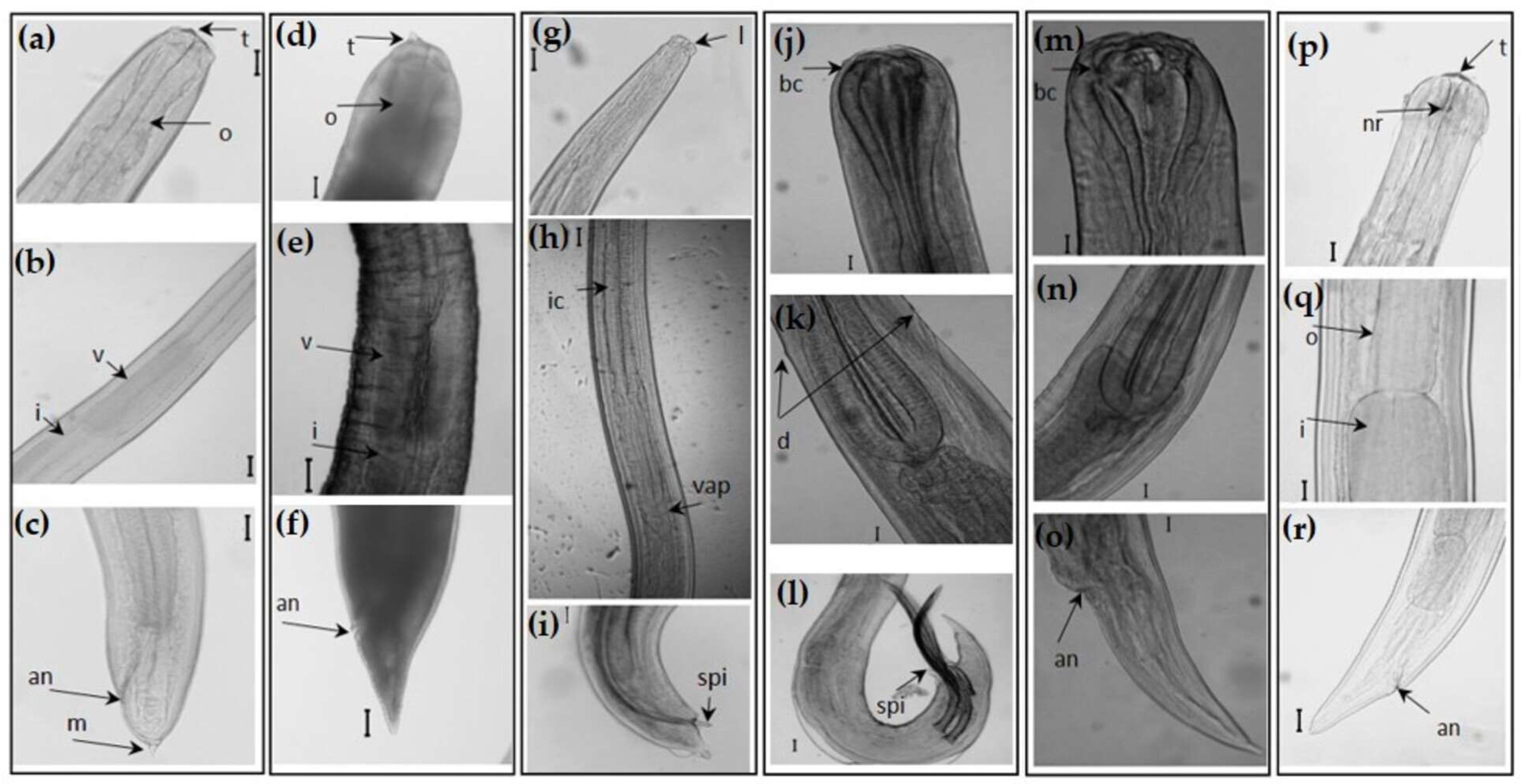{kind=link}
| Parameter | Mean ± SD | Range |
|---|---|---|
| Total length (cm) | 75.3 ± 26.7 | 180.0–29.8 |
| Total weight 1 (g) | 1252.1 ± 1972.1 | 15,000.0–31.9 |
| Eviscerated weight 2 (g) | 976.3 ± 1293.7 | 8200.0–27.4 |
| Parasite Specie | N | Prevalence % (95% CI) | Mean Intensity (95% CI) | Mean Abundance (95% CI) |
|---|---|---|---|---|
| Anisakis type I (sensu Berland, 1961) | 131 | 33.3 (25.1–42.7) | 3.6 (2.7–4.6) | 1.2 (0.7–1.8) |
| Anisakis type II (sensu Berland, 1961) | 18 | 10.2 (5.6–17.5) | 1.6 (1.2–2.1) | 0.2 (0.05–0.29) |
| Cucullanus sp. | 391 | 79.6 (71.0–86.2) | 4.5 (3.6–5.5) | 3.6 (2.8–4.5) |
| Cristitectus congeri | 69 | 13.0 (7.8–20.7) | 4.9 (3.2–6.7) | 0.6 (0.06–1.2) |
| Hysterothylacium spp. | 48 | 26.9 (19.4–36.0) | 1.7 (1.3–2.0) | 0.4 (0.3–0.6) |
| Species | N. of Preys (%) | Range |
|---|---|---|
| Fish-Osteichthyes | ||
| Dalophis imberbis (Delaroche, 1809) | 1 (1.3) | - |
| Species of Serranus Cuvier, 1816 | 1 (1.3) | - |
| Serranus cabrilla (Linnaeus, 1758) | 2 (2.5) | 1–1 |
| Merluccius merluccius (Linnaeus, 1758) | 1 (1.3) | - |
| Mullus surmuletus Linnaeus, 1758 | 1 (1.3) | - |
| Mullus barbatus Linnaeus, 1758 | 1 (1.3) | - |
| Gnathophis mystax (Delaroche, 1809) | 1 (1.3) | - |
| Family Scombridae Rafinesque, 1815 | 1 (1.3) | - |
| Micromesistius poutassou (Risso, 1827) | 15 (19.0) | 1–4 |
| Sardina pilchardus (Walbaum, 1792) | 8 (10.1) | 1–3 |
| Phycis blennoides (Brünnich, 1768) | 1 (1.3) | - |
| Conger conger (Linnaeus, 1758) | 2 (2.5) | 1–1 |
| Family Argentinidae Bonaparte, 1846 | 1 (1.3) | - |
| Argentina sphyraena Linnaeus, 1758 | 2 (2.5) | - 1 |
| Family Gonostomatidae Cocco, 1838 | 1 (1.3) | - |
| Boops boops (Linnaeus, 1758) | 1 (1.3) | - |
| Lepidorhombus boscii (Risso, 1810) | 1 (1.3) | - |
| Peristedion cataphractum (Linnaeus, 1758) | 1 (1.3) | - |
| Scyliorhinus canicula (Linnaeus, 1758) | 1 (1.3) | - |
| Trachyrhynchus trachyrhynchus (Risso, 1810) | 1 (1.3) | - |
| Species of Apterichtus Duméril, 1806 | 1 (1.3) | - |
| Pagrus pagrus (Linnaeus, 1758) | 1 (1.3) | - |
| Diplodus sargus sargus (Linnaeus, 1758) | 1 (1.3) | - |
| Crustacea-Decapod | ||
| Scyllarus pygmaeus (Spence Bate, 1888) | 1 (1.5) | - |
| Species of Processa Leach, 1815 | 2 (3.1) | 1–1 |
| Processa canaliculata Leach, 1815 | 1 (1.5) | - |
| Processa edulis (Risso, 1816) | 1 (1.5) | - |
| Species of Macropipus Prestandrea, 1833 | 11 (16.9) | 1–3 |
| Macropipus puber (Linnaeus, 1767) | 11 (16.9) | 1–4 |
| Liocarcinus depurator (Linnaeus, 1758) | 2 (3.1) | 1–1 |
| Goneplax rhomboides (Linnaeus, 1758) | 8 (12.3) | 1–4 |
| Solenocera membranacea (Risso, 1816) | 1 (1.5) | - |
| Species of Alpheus Fabricius, 1798 | 1 (1.5) | - |
| Alpheus glaber (Olivi, 1792) | 1 (1.5) | - |
| Alpheus macrocheles (Hailstone, 1835) | 1 (1.5) | - |
| Galathea strigosa Linnaeus, 1761) | 1 (1.5) | - |
| Munida intermedia A. Milne-Edwards & Bouvier, 1899 | 2 (3.1) | - 1 |
| Aristeus antennatus (Risso, 1816) | 1 (1.5) | - |
| Calappa granulata (Linnaeus, 1758) | 1 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roca-Geronès, X.; Sarrà-Alarcón, L.; Delgado, E.; Alcover, M.M.; Casadevall, M.; Fisa, R. Anisakid Presence in the European Conger, Conger conger, from Spanish Mediterranean Waters. Pathogens 2023, 12, 1326. https://doi.org/10.3390/pathogens12111326
Roca-Geronès X, Sarrà-Alarcón L, Delgado E, Alcover MM, Casadevall M, Fisa R. Anisakid Presence in the European Conger, Conger conger, from Spanish Mediterranean Waters. Pathogens. 2023; 12(11):1326. https://doi.org/10.3390/pathogens12111326
Chicago/Turabian StyleRoca-Geronès, Xavier, Lídia Sarrà-Alarcón, Eulàlia Delgado, Maria Magdalena Alcover, Margarida Casadevall, and Roser Fisa. 2023. "Anisakid Presence in the European Conger, Conger conger, from Spanish Mediterranean Waters" Pathogens 12, no. 11: 1326. https://doi.org/10.3390/pathogens12111326