
An Epidemiological Update on Anisakis Nematode Larvae in Red Mullet (Mullus barbatus) from the Ligurian Sea


 , ,
, ,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Fish Sampling and Parasite Identification
2.2. Molecular Analysis
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sieli, G.; Badalucco, C.; Di Stefano, G.; Rizzo, P.; D’Anna, G.; Fiorentino, F. Biology of red mullet, Mullus barbatus (L. 1758), in the Gulf of Castellammare (NW Sicily, Mediterranean Sea) subject to a trawling ban. J. Appl. Ichthyol. 2011, 27, 1218–1225. [Google Scholar] [CrossRef]
- Esposito, V.; Andaloro, F.; Bianca, D.; Natalotto, A.; Romeo, T.; Scotti, G.; Castriota, L. Diet and prey selectivity of the red mullet, Mullus barbatus (Pisces: Mullidae), from the southern Tyrrhenian Sea: The role of the surf zone as a feeding ground. Mar. Biol. Res. 2014, 10, 167–178. [Google Scholar] [CrossRef]
- Tserpes, G.; Fiorentino, F.; Levi, D.; Cau, A.; Murenu, M.; Zamboni, A.; Papaconstantinou, C. Distribution of Mullus barbatus and M. surmuletus (Osteichthyes: Perciformes) in the Mediterranean continental shelf: Implications for management. Sci. Mar. 2002, 66, 39–54. [Google Scholar] [CrossRef]
- Lombarte, A.; Recasens, L.; Gonzalez, M.; de Sola, L.G. Spatial segregation of two species of Mullidae (Mullus surmuletus and M. barbatus) in relation to habitat. Mar. Ecol. Prog. Ser. 2000, 206, 23949. [Google Scholar] [CrossRef]
- Cherif, M.; Zarrad, R.; Gharbi, H.; Missaoui, H.; Jarboui, O. Some biological parameters of the red mullet, Mullus barbatus L., 1758, from the Gulf of Tunis. Acta Adriat. 2007, 48, 131–144. [Google Scholar]
- Carbonara, P.; Intini, S.; Modugno, E.; Maradonna, F.; Spedicato, M.T.; Lembo, G.; Carnevali, O. Reproductive biology characteristics of red mullet (Mullus barbatus L., 1758) in Southern Adriatic Sea and management implications. Aquat. Living Resour. 2015, 28, 21–31. [Google Scholar] [CrossRef]
- Gismondi, R.; Labanchi, L.; Marzocchi, B.M.; Sermoneta, C. Statistiche Sulla Pesca in Italia, Uso Integrato di Indagini Campionarie e Dati Amministrativi. Available online: https://www.istat.it/it/files/2020/05/IWP-4-2020.pdf (accessed on 10 June 2023).
- Adroher-Auroux, F.J.; Benítez-Rodríguez, R. Anisakiasis and Anisakis: An underdiagnosed emerging disease and its main etiological agents. Res. Vet. Sci. 2020, 132, 535–545. [Google Scholar] [CrossRef]
- Rahmati, A.R.; Kiani, B.; Afshari, A.; Moghaddas, E.; Williams, M.; Shamsi, S. World-Wide Prevalence of Anisakis Larvae in Fish and Its Relationship to Human Allergic Anisakiasis: A Systematic Review. Parasitol. Res. 2020, 119, 3585–3594. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular Epidemiology of Anisakis and Anisakiasis: An Ecological and Evolutionary Road Map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [CrossRef]
- Klimpel, S.; Palm, H.W. Anisakid Nematode (Ascaridoidea) Life Cycles and Distribution: Increasing Zoonotic Potential in the Time of Climate Change? In Progress in Parasitology. Parasitology Research Monographs; Mehlhorn, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 201–222. [Google Scholar] [CrossRef]
- Menconi, V.; Pastorino, P.; Burioli, E.A.V.; Righetti, M.; Scanzio, T.; Favaro, L.; Bona, M.C.; Pavoletti, E.; Dondo, A.; Prearo, M. Occurrence of ascaridoid nematodes in Illex coindetii, a commercially relevant cephalopod species from the Ligurian Sea (Northwest Mediterranean Sea). Food Control 2020, 116, 107311. [Google Scholar] [CrossRef]
- Sakanari, J.A.; McKerrow, J.H. Identification of the secreted neutral proteases from Anisakis simplex. J. Parasitol. 1990, 76, 625–630. [Google Scholar] [CrossRef]
- Baptista-Fernandes, T.; Rodrigues, M.; Castro, I.; Paixão, P.; Pinto-Marques, P.; Roque, L.; Toscano, C. Human gastric hyperinfection by Anisakis simplex: A severe and unusual presentation and a brief review. Int. J. Infect. Dis. 2017, 64, 38–41. [Google Scholar] [CrossRef]
- Takabayashi, T.; Mochizuki, T.; Otani, N.; Nishiyama, K.; Ishimatsu, S. Anisakiasis presenting to the ED: Clinical manifestations, time course, hematologic tests, computed tomographic findings, and treatment. Am. J. Emerg. Med. 2014, 32, 1485–1489. [Google Scholar] [CrossRef]
- Shimamura, Y.; Muwanwella, N.; Chandran, S.; Kandel, G.; Marcon, N. Common symptoms from an uncommon infection: Gastrointestinal anisakiasis. Can. J. Gastroenterol. Hepatol. 2016, 2016, 5176502. [Google Scholar] [CrossRef]
- Daschner, A.; Cuéllar, C.; Rodero, M. The Anisakis allergy debate: Does an evolutionary approach help? Trends Parasitol. 2012, 28, 9–15. [Google Scholar] [CrossRef]
- Daschner, A.; Alonso-Gómez, A.; Cabañas, R.; Suarez-de-Parga, J.M.; López-Serrano, M.C. Gastroallergic anisakiasis: Borderline between food allergy and parasitic disease—Clinical and allergology evaluation of 20 patients with confirmed acute parasitism by Anisakis simplex. J. Allergy Clin. Immunol. 2000, 105, 176–181. [Google Scholar] [CrossRef]
- Manfredi, M.; Crosa, G.; Galli, P.; Ganduglia, S. Distribution of Anisakis simplex in fish caught in the Ligurian Sea. Parasitol. Res. 2000, 86, 551–553. [Google Scholar] [CrossRef]
- Serracca, L.; Cencetti, E.; Battistini, R.; Rossini, I.; Prearo, M.; Pavoletti, E.; Fioravanti, M.L.; Righetti, M.; Di Donfrancesco, B.; Ercolini, C. Survey on the presence of Anisakis and Hysterothylacium larvae in fishes and squids caught in Ligurian Sea. Vet. Parasitol. 2013, 196, 547–551. [Google Scholar] [CrossRef]
- Debenedetti, A.L.; Madrid, E.; Fuentes, M.V. Study of helminth parasites in the red mullet, Mullus barbatus, from the Mediterranean Sea and acquired in greater València, Spain. Rev. Ibero-Latinoam. Parasitol. 2013, 72, 118–123. [Google Scholar]
- Pekmezci, G.Z.; Onuk, E.E.; Bolukbas, C.S.; Yardimci, B.; Gurler, A.T.; Acici, M.; Umur, S. Molecular identification of Anisakis species (Nematoda: Anisakidae) from marine fishes collected in Turkish waters. Vet. Parasitol. 2014, 201, 82–94. [Google Scholar] [CrossRef]
- Barcala, E.; Ramilo, A.; Ortega, N.; Picó, G.; Abollo, E.; Pascual, S.; Muñoz, P. Occurrence of Anisakis and Hysterothylacium larvae in commercial fish from Balearic Sea (Western Mediterranean Sea). Parasitol. Res. 2018, 117, 4003–4012. [Google Scholar] [CrossRef]
- Kassem, H.; Rima, E.M.; Bojwari, G.; Bowashi, S.; Hador, A. Nematode infestation in Red Mullet (Mullus surmuletus, Linnaeus, 1758) from Benghazi Coast, Libya. Alq. J. Med. App. Sci. 2023, 6, 81–88. [Google Scholar] [CrossRef]
- Pulleiro-Potel, L.; Barcala, E.; Mayo-Hernández, E.; Muñoz, P. Survey of anisakids in commercial teleosts from the western Mediterranean Sea: Infection rates and possible effects of environmental and ecological factors. Food Control 2015, 55, 12–17. [Google Scholar] [CrossRef]
- Klimpel, S.; Kleinertz, S.; Palm, H.W. Distribution of parasites from red mullets (Mullus surmuletus L., Mullidae) in the North Sea and the Mediterranean Sea. Bull. Fish Biol. 2008, 10, 25–38. [Google Scholar]
- Karl, H.; Levsen, A. Occurrence, and distribution of anisakid nematodes in grey gurnard (Eutrigla gurnardus L.) from the North Sea. Food Control 2011, 22, 1634–1638. [Google Scholar] [CrossRef]
- Berland, B. Nematodes from some Norwegian marine fishes. Sarsia 1961, 2, 1–50. [Google Scholar]
- D’Amelio, S.; Mathiopoulos, K.D.; Santos, C.P.; Pugachev, O.N.; Webb, S.C.; Picanço, M.; Paggi, L. Genetic markers in ribosomal DNA for the identification of members of the genus Anisakis (Nematoda: Ascaridoidea) defined by polymerase-chain-reaction-based restriction fragment length polymorphism. Int. J. Parasitol. 2000, 30, 223–226. [Google Scholar] [CrossRef]
- Pontes, T.; D’Amelio, S.; Costa, G.; Paggi, L. Molecular characterization of larval anisakid nematodes from marine fishes of Madeira by a PCR-based approach, with evidence for a new species. J. Parasitol. 2005, 91, 1430–1434. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, M.; Shostak, A.W. Parasitology meets ecology. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Moravec, F. Parasitic Nematodes of Freshwater Fishes of Europe; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 1–473. [Google Scholar]
- Piras, M.C.; Tedde, T.; Garippa, G.; Virgilio, S.; Sanna, D.; Farjallah, S.; Merella, P. Molecular and epidemiological data on Anisakis spp. (Nematoda: Anisakidae) in commercial fish caught off northern Sardinia (western Mediterranean Sea). Vet. Parasitol. 2014, 203, 237–240. [Google Scholar] [CrossRef]
- Serracca, L.; Battistini, R.; Rossini, I.; Carducci, A.; Verani, M.; Prearo, M.; Tomei, L.; De Montis, G.; Ercolini, C. Food safety considerations in relation to Anisakis pegreffii in anchovy (Engraulis encrasicolus) and sardines (Sardina pilchardus) fished off the Ligurian coast (Cinque Terre National Park, NW Mediterranean). Int. J. Food Microbiol. 2014, 190, 79–83. [Google Scholar] [CrossRef]
- Menconi, V.; Pastorino, P.; Canola, S.; Pavoletti, E.; Vitale, N.; Scanzio, T.; Righetti, M.; Mugetti, D.; Tomasoni, M.; Bona, M.C.; et al. Occurrence and spatial variation of Anisakis pegreffii in the Atlantic horse mackerel Trachurus trachurus (Carangidae): A three-year monitoring survey in the western Ligurian Sea. Food Control 2021, 131, 108423. [Google Scholar] [CrossRef]
- Abattouy, N.; López, A.V.; Maldonado, J.L.; Benajiba, M.H.; Martín-Sánchez, J. Epidemiology and molecular identification of Anisakis pegreffii (Nematoda: Anisakidae) in the horse mackerel Trachurus trachurus from northern Morocco. J. Helminthol. 2014, 88, 257–263. [Google Scholar] [CrossRef]
- Levsen, A.; Svanevik, C.S.; Cipriani, P.; Mattiucci, S.; Gay, M.; Hastie, L.C.; Bušelić, I.; Mladineo, I.; Karl, H.G.N.D.; Ostermeyer, U.; et al. A survey of zoonotic nematodes of commercial key fish species from major European fishing grounds—Introducing the FP7 PARASITE exposure assessment study. Fish. Res. 2018, 202, 4–21. [Google Scholar] [CrossRef]
- Ferrer-Maza, D.; Lloret, J.; Munoz, M.; Faliex, E.; Vila, S.; Sasal, P. Parasitism, condition, and reproduction of the European hake (Merluccius merluccius) in the north-western Mediterranean Sea. ICES J. Mar. Sci. 2014, 71, 1088–1099. [Google Scholar] [CrossRef]
- Ferrer-Maza, D.; Muñoz, M.; Lloret, J.; Faliex, E.; Vila, S.; Sasal, P. Health and re-production of red mullet, Mullus barbatus, in the western Mediterranean Sea. Hydrobiologia 2015, 753, 189–204. [Google Scholar] [CrossRef]
- Mouritsen, K.N.; Hedeholm, R.; Schack, H.B.; Møller, L.N.; Storr-Paulsen, M.; Dzido, J.; Rokicki, J. Occurrence of anisakid nematodes in Atlantic cod (Gadus morhua) and Greenland cod (Gadus ogac), West Greenland. Acta Parasitol. 2010, 55, 81–89. [Google Scholar] [CrossRef]
- Mattiucci, S.; Abaunza, P.; Ramadori, L.; Nascetti, G. Genetic identification of Anisakis larvae in European hake from Atlantic and Mediterranean waters for stock recognition. J. Fish Biol. 2004, 65, 495–510. [Google Scholar] [CrossRef]
- Notarbartolo di Sciara, G.; Agardy, T.; Hyrenbach, D.; Scovazzi, T.; Van Klaveren, P. The Pelagos Sanctuary for Mediterranean marine mammals. Aquat. Conser. Mar. Freshw. Ecosyst. 2008, 18, 367–391. [Google Scholar] [CrossRef]
- Mattiucci, S.; Nascetti, G. Advances and trends in the molecular systematics of anisakid nematodes, with implications for their evolutionary ecology and host-parasite co-evolutionary processes. Adv. Parasitol. 2008, 66, 47–148. [Google Scholar] [CrossRef]
- Marcogliese, D.J. Distribution and abundance of sealworm (Pseudoterranova decipiens) and other anisakid nematodes in fish and seals in the Gulf of St. Lawrence: Potential importance of climatic conditions. NAMMCO Sci. Publ. 2001, 3, 113–128. [Google Scholar] [CrossRef]
- Corradi, N.; Fanucci, F.; Fierro, G.; Firpo, M.; Piccazzo, M.; Mirabile, L. La Piattaforma Continentale Ligure: Caratteri, Struttura ed Evoluzione; Rapporto Tecnico Finale Progetto “Oceanografia e Fondi Marini” CNR: Roma, Italy, 1984. [Google Scholar]
- Valisano, L.; Notari, F.; Mori, M.; Cerrano, C. Temporal variability of sedimentation rates and mobile fauna inside and outside a gorgonian garden. Mar. Ecol. 2016, 37, 1303–1314. [Google Scholar] [CrossRef]
- EFSA Scientific Opinion on Risk Assessment of Parasites in Fishery Products. EFSA Panel on Biological Hazards. 2010. Available online: https://www.sanipes.gob.pe/archivos/2010-EFSA.pdf (accessed on 10 June 2023).




{kind=link}
{kind=link}
{kind=link}
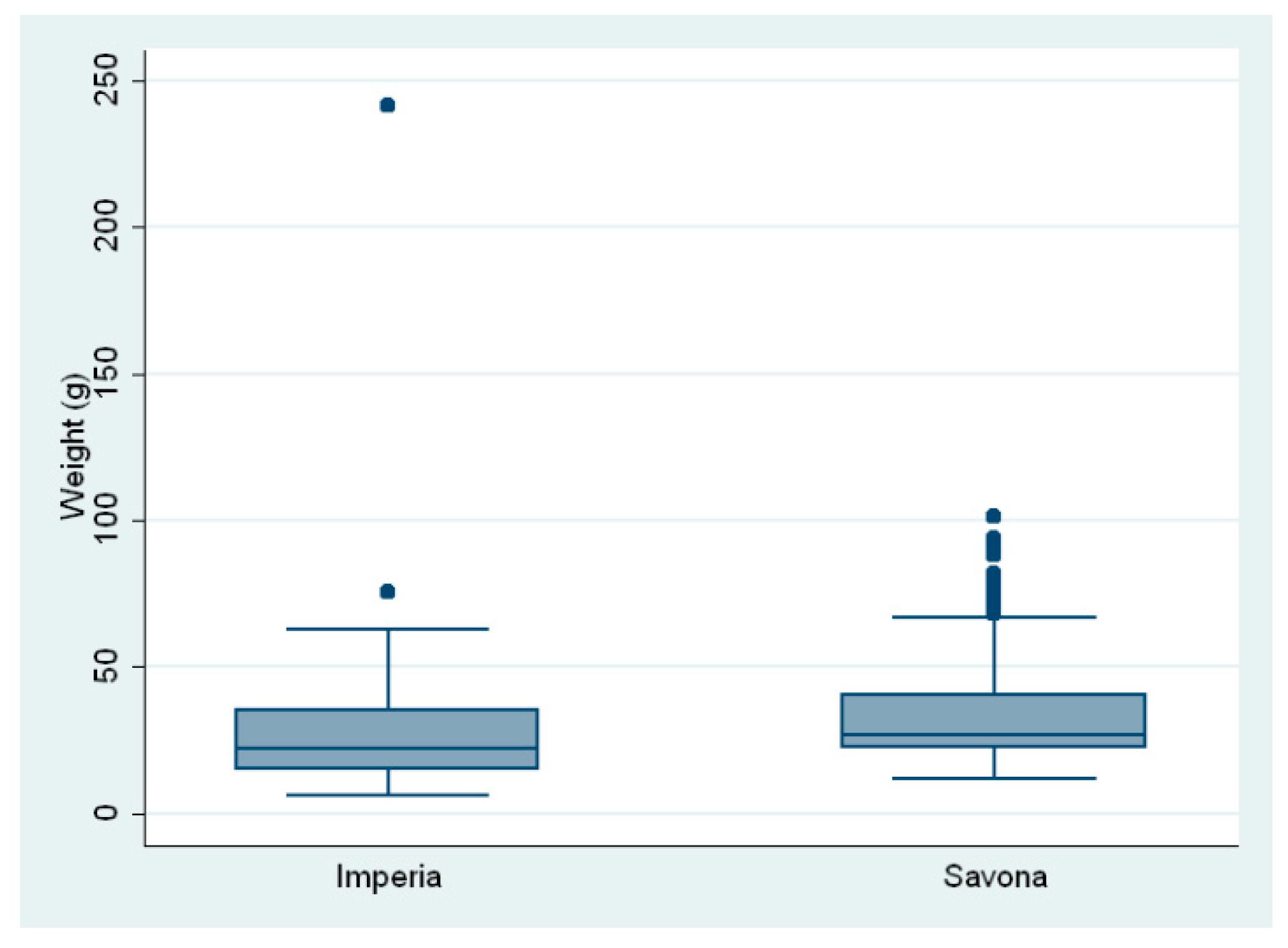{kind=link}
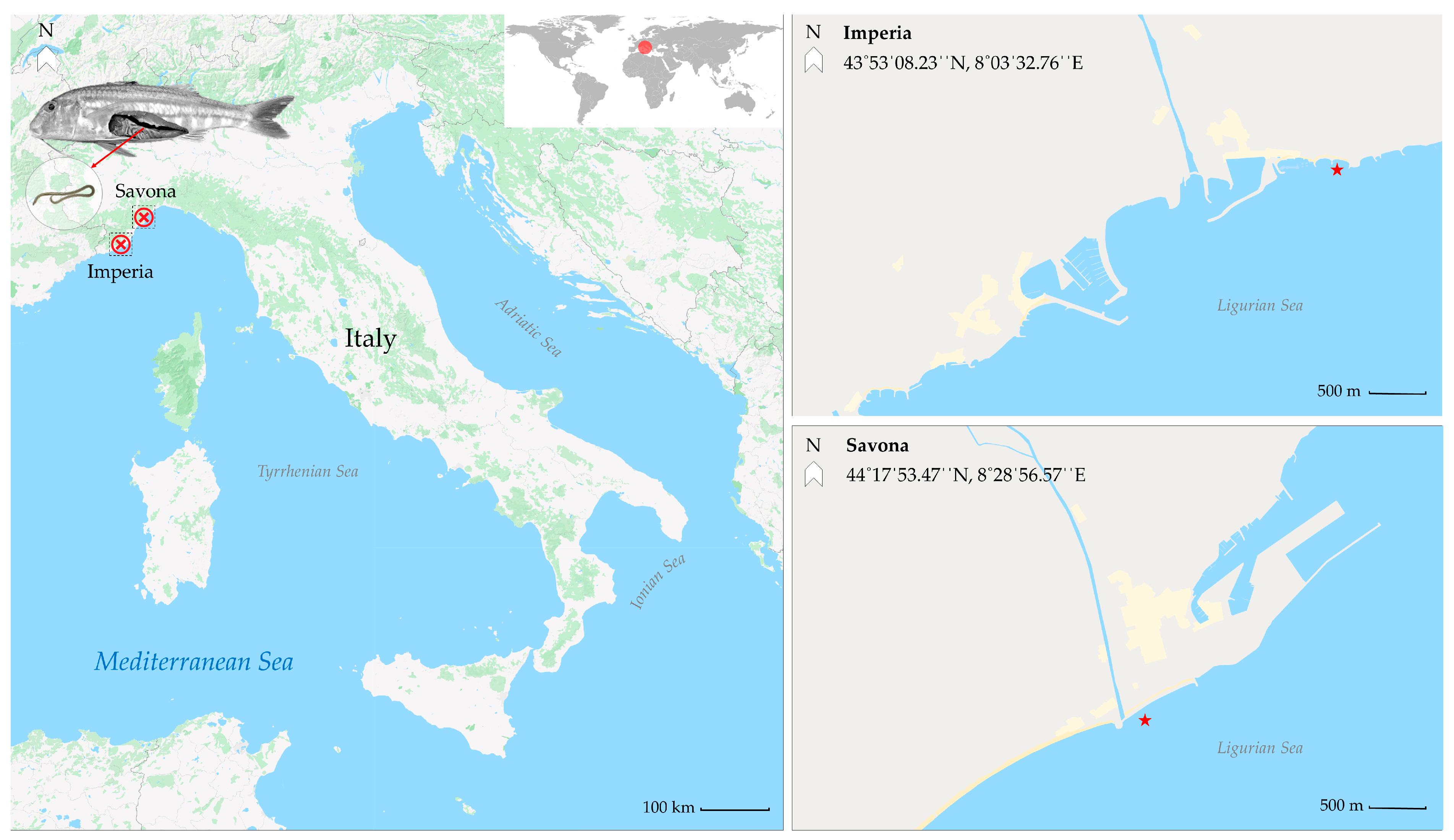
| Sampling Site | Red Mullet Sampled | Parasite | Infected Fish | Larvae | %P (95% CI) | MI (Range) | MA (95% CI) | Coinfected Individuals (%) |
|---|---|---|---|---|---|---|---|---|
| Imperia | 190 | Ascaridoid larvae | 127 | 661 | 66.84 (60.09–73.60) | 5.20 (1–130) | 3.48 (2.98–4.20) | 12 (6.31%) |
| Anisakis pegreffii | 12 | 18 | 6.32 (2.83–9.81) | 1.5 (1–6) | 0.09 (0.02–0.17) | |||
| Hysterothylacium spp. | 127 | 643 | 66.84 (60.09–73.60) | 5.09 (1–124) | 3.38 (1.95–4.82) | |||
| Savona | 648 | Ascaridoid larvae | 530 | 3826 | 81.79 (78.67–84.23) | 7.22 (1–70) | 5.90 (5.11–6.10) | 148 (22.83%) |
| A. pegreffii | 193 | 526 | 29.78 (26.25–33.31) | 2.73 (1–11) | 0.81 (0.68–0.94) | |||
| Hysterothylacium spp. | 485 | 3300 | 74.69 (71.33–78.05) | 6.80 (1–67) | 5.09 (4.46–5.73) | |||
| Total | 838 | Ascaridoid larvae | 657 | 4487 | 78.40 (76.16–80.56) | 6.82 (1–130) | 5.35 (4.21–5.86) | 160 (19.09%) |
| A. pegreffii | 205 | 544 | 24.46 (21.55–27.38) | 2.65 (1–11) | 0.65 (0.54–0.75) | |||
| Hysterothylacium spp. | 612 | 3943 | 73.03 (71.98–75.15) | 6.44 (1–124) | 4.71 (4.12–5.29) |
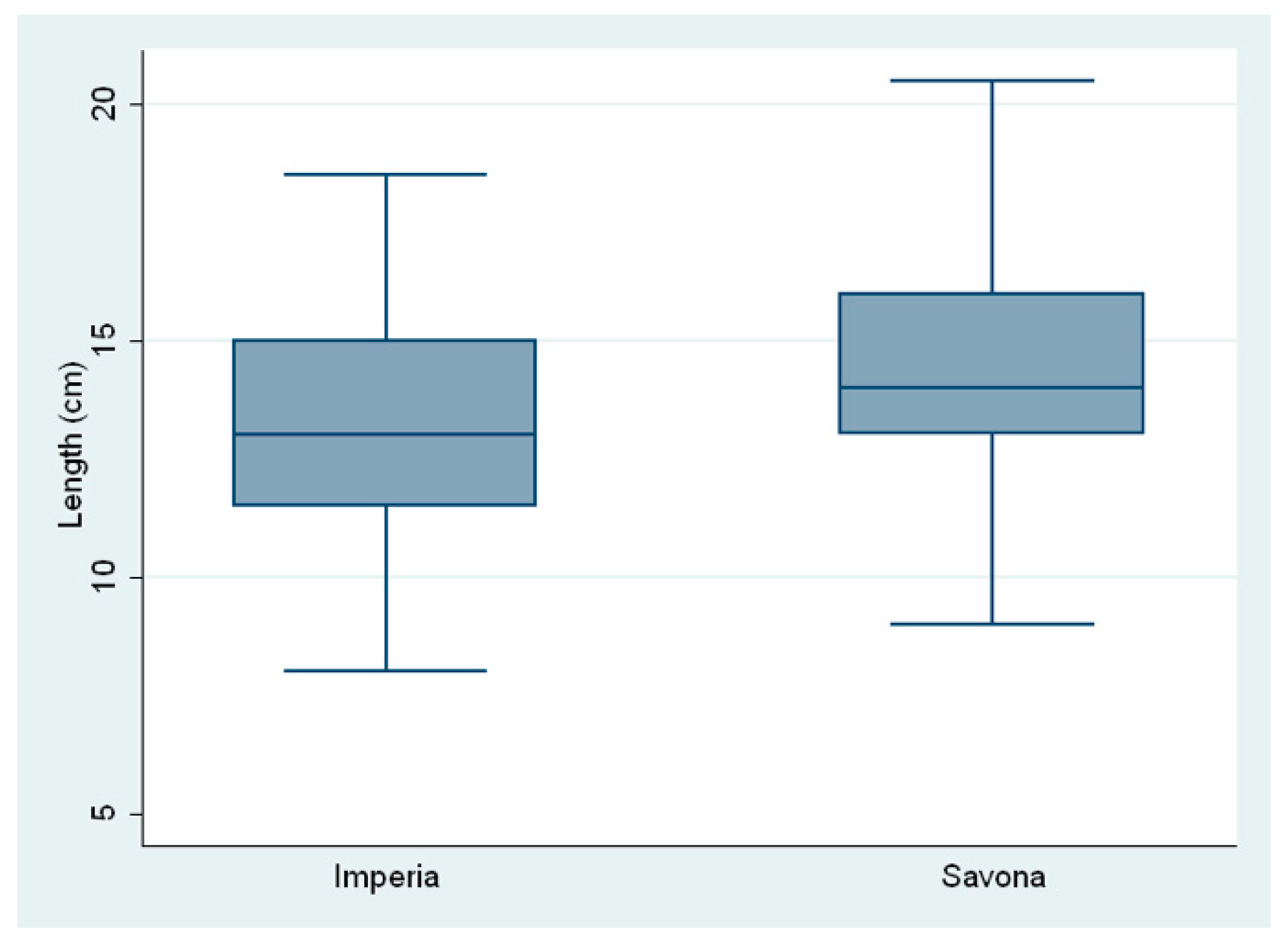
| Larvae (N) | Weight (g) | Length (cm) | |
|---|---|---|---|
| No. of larvae | 1.0000 | ||
| Weight (g) | 0.427 * | 1.0000 | |
| Length (cm) | 0.407 * | 0.947 * | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schleicherová, D.; Menconi, V.; Moroni, B.; Pastorino, P.; Esposito, G.; Canola, S.; Righetti, M.; Dondo, A.; Prearo, M. An Epidemiological Update on Anisakis Nematode Larvae in Red Mullet (Mullus barbatus) from the Ligurian Sea. Pathogens 2023, 12, 1366. https://doi.org/10.3390/pathogens12111366
Schleicherová D, Menconi V, Moroni B, Pastorino P, Esposito G, Canola S, Righetti M, Dondo A, Prearo M. An Epidemiological Update on Anisakis Nematode Larvae in Red Mullet (Mullus barbatus) from the Ligurian Sea. Pathogens. 2023; 12(11):1366. https://doi.org/10.3390/pathogens12111366
Chicago/Turabian StyleSchleicherová, Dáša, Vasco Menconi, Barbara Moroni, Paolo Pastorino, Giuseppe Esposito, Serena Canola, Marzia Righetti, Alessandro Dondo, and Marino Prearo. 2023. "An Epidemiological Update on Anisakis Nematode Larvae in Red Mullet (Mullus barbatus) from the Ligurian Sea" Pathogens 12, no. 11: 1366. https://doi.org/10.3390/pathogens12111366