

Oreoch-1: A Peptide from Oreochromis niloticus as a Potential Tool against Staphylococci

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Peptide Synthesis and Characterization
2.2. In Vitro Cytotoxicity Evaluation
2.3. Microorganisms: Isolation, Drug Susceptibility, and Culture Conditions
2.4. Antibacterial Assay
2.5. Biofilm Maturation Test
2.6. Biofilm Evaluation by Fluorescence Microscopy
2.7. Statistical Analysis
3. Results
3.1. Evaluation of Cytotoxicity Profile
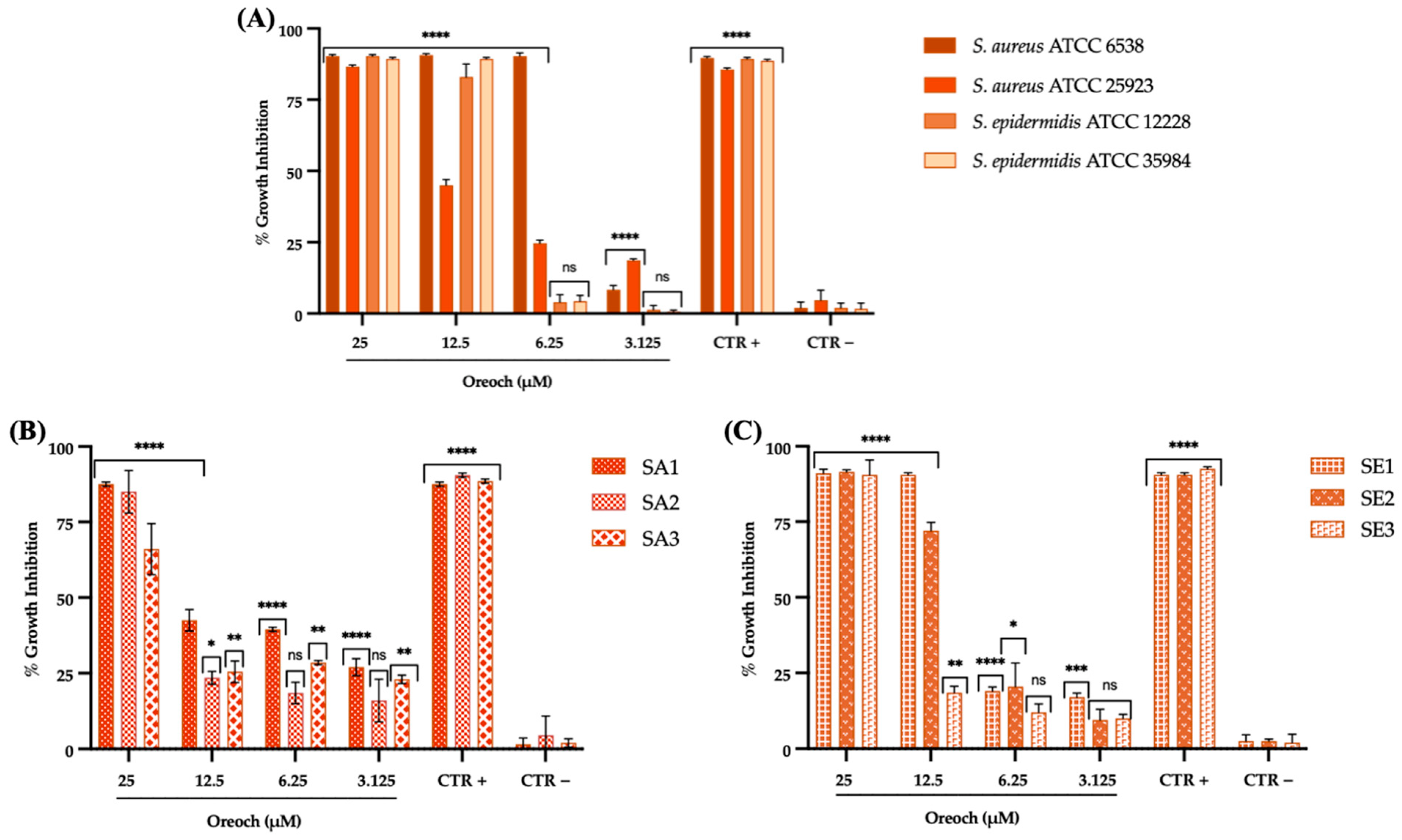
3.2. Evaluation of Antibacterial Effect
3.3. Antibiofilm Action Analyzed via Fluorescence Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bowler, P.; Murphy, C.; Wolcott, R. Biofilm Exacerbates Antibiotic Resistance: Is This a Current Oversight in Antimicrobial Stewardship? Antimicrob. Resist. Infect. Control 2020, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus Epidermidis—The “accidental” Pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-Negative Staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.W.; Dowd, S.E.; Brackee, G.; Fralick, J.A.; Vediyappan, G. Inhibition of Staphylococcus Aureus Biofilm Formation by Gurmarin, a Plant-Derived Cyclic Peptide. Front. Cell. Infect. Microbiol. 2022, 12, 1017545. [Google Scholar] [CrossRef] [PubMed]
- Kırmusaoğlu, S.; Kaşıkçı, H. Identification of Ica -Dependent Biofilm Production by Staphylococcus Aureus Clinical Isolates and Antibiofilm Effects of Ascorbic Acid against Biofilm Production. J. Clin. Pathol. 2020, 73, 261–266. [Google Scholar] [CrossRef]
- Khaleghi, M.; Khorrami, S. Down-Regulation of Biofilm-Associated Genes in MecA-Positive Methicillin-Resistant S. Aureus Treated with M. Communis Extract and Its Antibacterial Activity. AMB Express 2021, 11, 85. [Google Scholar] [CrossRef]
- ECDC: Antimicrobial Resistance Surveillance Data in Europe 2022–2020. Available online: Https://Www.Ecdc.Europa.Eu/En/Publications-Data/Antimicrobial-Resistance-Surveillance-Europe-2022-2020-Data (accessed on 31 July 2023).
- Lisowska-Łysiak, K.; Lauterbach, R.; Międzobrodzki, J.; Kosecka-Strojek, M. Epidemiology and Pathogenesis of Staphylococcus Bloodstream Infections in Humans: A Review. Pol. J. Microbiol. 2021, 70, 13–23. [Google Scholar] [CrossRef]
- Stewart, E.J.; Payne, D.E.; Ma, T.M.; VanEpps, J.S.; Boles, B.R.; Younger, J.G.; Solomon, M.J. Effect of Antimicrobial and Physical Treatments on Growth of Multispecies Staphylococcal Biofilms. Appl. Environ. Microbiol. 2017, 83, e03483-16. [Google Scholar] [CrossRef]
- Delpech, G.; Ceci, M.; Lissarrague, S.; García Allende, L.; Baldaccini, B.; Sparo, M. In Vitro Activity of the Antimicrobial Peptide AP7121 against the Human Methicillin-Resistant Biofilm Producers Staphylococcus Aureus and Staphylococcus Epidermidis. Biofouling 2020, 36, 266–275. [Google Scholar] [CrossRef]
- Deplano, A.; Vandendriessche, S.; Nonhoff, C.; Dodémont, M.; Roisin, S.; Denis, O. National Surveillance of Staphylococcus Epidermidis Recovered from Bloodstream Infections in Belgian Hospitals. J. Antimicrob. Chemother. 2016, 71, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Memariani, H.; Memariani, M.; Robati, R.M.; Nasiri, S.; Abdollahimajd, F.; Baseri, Z.; Moravvej, H. Anti-Staphylococcal and Cytotoxic Activities of the Short Anti-Microbial Peptide PVP. World J. Microbiol. Biotechnol. 2020, 36, 174. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Sharma, G.; Dang, S.; Gupta, S.; Gabrani, R. Antimicrobial Peptides as Anti-Infectives against Staphylococcus Epidermidis. Med. Princ. Pract. 2016, 25, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Chianese, A.; Zannella, C.; Monti, A.; De Filippis, A.; Doti, N.; Franci, G.; Galdiero, M. The Broad-Spectrum Antiviral Potential of the Amphibian Peptide AR-23. Int. J. Mol. Sci. 2022, 23, 883. [Google Scholar] [CrossRef]
- Chianese, A.; Zannella, C.; Foglia, F.; Nastri, B.M.; Monti, A.; Doti, N.; Franci, G.; De Filippis, A.; Galdiero, M. Hylin-A1: A Host Defense Peptide with Antibacterial Potential against Staphylococcus Aureus Multi-Resistant Strains. Pharmaceuticals 2023, 16, 509. [Google Scholar] [CrossRef]
- Zhu, S.; Li, W.; O’Brien-Simpson, N.; Separovic, F.; Sani, M.-A. C-Terminus Amidation Influences Biological Activity and Membrane Interaction of Maculatin 1.1. Amino Acids 2021, 53, 769–777. [Google Scholar] [CrossRef]
- Caporale, A.; Doti, N.; Monti, A.; Sandomenico, A.; Ruvo, M. Automatic Procedures for the Synthesis of Difficult Peptides Using Oxyma as Activating Reagent: A Comparative Study on the Use of Bases and on Different Deprotection and Agitation Conditions. Peptides 2018, 102, 38–46. [Google Scholar] [CrossRef]
- Pignataro, D.; Foglia, F.; Della Rocca, M.T.; Melardo, C.; Santella, B.; Folliero, V.; Shinde, S.; Pafundi, P.C.; Sasso, F.C.; Iovene, M.R.; et al. Methicillin-Resistant Staphylococcus Aureus: Epidemiology and Antimicrobial Susceptibility Experiences from the University Hospital ‘Luigi Vanvitelli’ of Naples. Pathog. Glob. Health 2020, 114, 451–456. [Google Scholar] [CrossRef]
- Dell’Annunziata, F.; Folliero, V.; Palma, F.; Crudele, V.; Finamore, E.; Sanna, G.; Manzin, A.; De Filippis, A.; Galdiero, M.; Franci, G. Anthraquinone Rhein Exhibits Antibacterial Activity against Staphylococcus Aureus. Appl. Sci. 2022, 12, 8691. [Google Scholar] [CrossRef]
- Yadav, M.K.; Chae, S.-W.; Im, G.J.; Chung, J.-W.; Song, J.-J. Eugenol: A Phyto-Compound Effective against Methicillin-Resistant and Methicillin-Sensitive Staphylococcus Aureus Clinical Strain Biofilms. PLoS ONE 2015, 10, e0119564. [Google Scholar] [CrossRef]
- Tuon, F.F.; Suss, P.H.; Telles, J.P.; Dantas, L.R.; Borges, N.H.; Ribeiro, V.S.T. Antimicrobial Treatment of Staphylococcus Aureus Biofilms. Antibiotics 2023, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Scheper, H.; Wubbolts, J.M.; Verhagen, J.A.M.; De Visser, A.W.; Van Der Wal, R.J.P.; Visser, L.G.; De Boer, M.G.J.; Nibbering, P.H. SAAP-148 Eradicates MRSA Persisters Within Mature Biofilm Models Simulating Prosthetic Joint Infection. Front. Microbiol. 2021, 12, 625952. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Lu, T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, Y.; Dong, M.; Hang, B.; Sun, Y.; Wang, L.; Wang, Y.; Hu, J.; Zhang, W. HJH-1, a Broad-Spectrum Antimicrobial Activity and Low Cytotoxicity Antimicrobial Peptide. Molecules 2018, 23, 2026. [Google Scholar] [CrossRef] [PubMed]
- Dijksteel, G.S.; Ulrich, M.M.W.; Middelkoop, E.; Boekema, B.K.H.L. Review: Lessons Learned From Clinical Trials Using Antimicrobial Peptides (AMPs). Front. Microbiol. 2021, 12, 616979. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Wang, Y.; Yang, Y.; Li, L.; Yuan, K.; Cao, H.; Qiu, Z.; Guo, G.; Wu, J.; Peng, J. Cec4-Derived Peptide Inhibits Planktonic and Biofilm-Associated Methicillin Resistant Staphylococcus Epidermidis. Microbiol. Spectr. 2022, 10, e02409-22. [Google Scholar] [CrossRef]
- Zhu, C.; Zhao, Y.; Zhao, X.; Liu, S.; Xia, X.; Zhang, S.; Wang, Y.; Zhang, H.; Xu, Y.; Chen, S.; et al. The Antimicrobial Peptide MPX Can Kill Staphylococcus Aureus, Reduce Biofilm Formation, and Effectively Treat Bacterial Skin Infections in Mice. Front. Vet. Sci. 2022, 9, 819921. [Google Scholar] [CrossRef]
- Erdem Büyükkiraz, M.; Kesmen, Z. Antimicrobial Peptides (AMPs): A Promising Class of Antimicrobial Compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef]
- Pan, C.-Y.; Chen, J.-C.; Chen, T.-L.; Wu, J.-L.; Hui, C.-F.; Chen, J.-Y. Piscidin Is Highly Active against Carbapenem-Resistant Acinetobacter Baumannii and NDM-1-Producing Klebsiella Pneumonia in a Systemic Septicaemia Infection Mouse Model. Mar. Drugs 2015, 13, 2287–2305. [Google Scholar] [CrossRef]
- Nibbering, P.H.; Göblyös, A.; Adriaans, A.E.; Cordfunke, R.A.; Ravensbergen, B.; Rietveld, M.H.; Zwart, S.; Commandeur, S.; Van Leeuwen, R.; Haisma, E.M.; et al. Eradication of Meticillin-Resistant Staphylococcus Aureus from Human Skin by the Novel LL-37-Derived Peptide P10 in Four Pharmaceutical Ointments. Int. J. Antimicrob. Agents 2019, 54, 610–618. [Google Scholar] [CrossRef]
- Fernández-Cuenca, F.; López-Hernández, I.; Cercenado, E.; Conejo, C.; Tormo, N.; Gimeno, C.; Pascual, A. Reporting Antimicrobial Susceptibilities and Resistance Phenotypes in Staphylococcus Spp.: A Nationwide Proficiency Study. J. Antimicrob. Chemother. 2021, 76, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, T.; Lönn, J.; Khalaf, H.; Palm, E. The Lantibiotic Gallidermin Acts Bactericidal against Staphylococcus Epidermidis and Staphylococcus Aureus and Antagonizes the Bacteria-induced Proinflammatory Responses in Dermal Fibroblasts. Microbiologyopen 2018, 7, e00606. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.; Montero, V.; Carpio, Y.; Velázquez, J.; Garay, H.E.; Reyes, O.; Cabrales, A.; Masforrol, Y.; Morales, A.; Estrada, M.P. Cloning and Functional Characterization of Three Novel Antimicrobial Peptides from Tilapia (Oreochromis niloticus). Aquaculture 2013, 372–375, 9–18. [Google Scholar] [CrossRef]
- Kuo, H.-M.; Tseng, C.-C.; Chen, N.-F.; Tai, M.-H.; Hung, H.-C.; Feng, C.-W.; Cheng, S.-Y.; Huang, S.-Y.; Jean, Y.-H.; Wen, Z.-H. MSP-4, an Antimicrobial Peptide, Induces Apoptosis via Activation of Extrinsic Fas/FasL- and Intrinsic Mitochondria-Mediated Pathways in One Osteosarcoma Cell Line. Mar. Drugs 2018, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-N.; Chan, Y.-L.; Hui, C.-F.; Wu, J.-L.; Wu, C.-J.; Chen, J.-Y. Use of Tilapia Piscidin 3 (TP3) to Protect against MRSA Infection in Mice with Skin Injuries. Oncotarget 2015, 6, 12955–12969. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.-C.; Lee, S.-H.; Hour, A.-L.; Pan, C.-Y.; Lee, L.-H.; Chen, J.-Y. Five Different Piscidins from Nile Tilapia, Oreochromis Niloticus: Analysis of Their Expressions and Biological Functions. PLoS ONE 2012, 7, e50263. [Google Scholar] [CrossRef]
- Mihailescu, M.; Sorci, M.; Seckute, J.; Silin, V.I.; Hammer, J.; Perrin, B.S.; Hernandez, J.I.; Smajic, N.; Shrestha, A.; Bogardus, K.A.; et al. Structure and Function in Antimicrobial Piscidins: Histidine Position, Directionality of Membrane Insertion, and PH-Dependent Permeabilization. J. Am. Chem. Soc. 2019, 141, 9837–9853. [Google Scholar] [CrossRef]
- Lin, W.-C.; Chang, H.-Y.; Chen, J.-Y. Electrotransfer of the Tilapia Piscidin 3 and Tilapia Piscidin 4 Genes into Skeletal Muscle Enhances the Antibacterial and Immunomodulatory Functions of Oreochromis Niloticus. Fish Shellfish Immunol. 2016, 50, 200–209. [Google Scholar] [CrossRef]
- Pan, C.-Y.; Tsai, T.-Y.; Su, B.-C.; Hui, C.-F.; Chen, J.-Y. Study of the Antimicrobial Activity of Tilapia Piscidin 3 (TP3) and TP4 and Their Effects on Immune Functions in Hybrid Tilapia (Oreochromis Spp.). PLoS ONE 2017, 12, e0169678. [Google Scholar] [CrossRef]
- Le, M.N.-T.; Kawada-Matsuo, M.; Komatsuzawa, H. Efficiency of Antimicrobial Peptides Against Multidrug-Resistant Staphylococcal Pathogens. Front. Microbiol. 2022, 13, 930629. [Google Scholar] [CrossRef]
- Hou, Z.; Zhang, B.; She, P.; Wu, Y.; Zhang, R. Anti-Planktonic and Anti-Biofilm Effects of Two Synthetic Anti-Microbial Peptides against Staphylococcus Epidermidis. Zhong Nan Da Xue Xue Bao. Yi Xue Ban = J. Cent. South University. Med. Sci. 2021, 46, 481–487. [Google Scholar] [CrossRef]
- Choudhury, A.; Islam, S.M.A.; Ghidey, M.R.; Kearney, C.M. Repurposing a Drug Targeting Peptide for Targeting Antimicrobial Peptides against Staphylococcus. Biotechnol. Lett. 2020, 42, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, W.F.; Silva, P.M.S.; Silva, R.C.S.; Silva, G.M.M.; Machado, G.; Coelho, L.C.B.B.; Correia, M.T.S. Staphylococcus Aureus and Staphylococcus Epidermidis Infections on Implants. J. Hosp. Infect. 2018, 98, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-W.; Wei, S.-Y.; Wang, S.-H.; Wei, H.-M.; Wang, Y.-J.; Wang, C.-F.; Chen, C.; Liao, Y.-D. Hydrophobic Residues Are Critical for the Helix-Forming, Hemolytic and Bactericidal Activities of Amphipathic Antimicrobial Peptide TP4. PLoS ONE 2017, 12, e0186442. [Google Scholar] [CrossRef]
- Mazurkiewicz-Pisarek, A.; Baran, J.; Ciach, T. Antimicrobial Peptides: Challenging Journey to the Pharmaceutical, Biomedical, and Cosmeceutical Use. Int. J. Mol. Sci. 2023, 24, 9031. [Google Scholar] [CrossRef]
- Zarghami, V.; Ghorbani, M.; Pooshang Bagheri, K.; Shokrgozar, M.A. Melittin Antimicrobial Peptide Thin Layer on Bone Implant Chitosan-Antibiotic Coatings and Their Bactericidal Properties. Mater. Chem. Phys. 2021, 263, 124432. [Google Scholar] [CrossRef]
- Haisma, E.M.; De Breij, A.; Chan, H.; Van Dissel, J.T.; Drijfhout, J.W.; Hiemstra, P.S.; El Ghalbzouri, A.; Nibbering, P.H. LL-37-Derived Peptides Eradicate Multidrug-Resistant Staphylococcus Aureus from Thermally Wounded Human Skin Equivalents. Antimicrob. Agents Chemother. 2014, 58, 4411–4419. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Staphylococci | Strain Code | Resistance Profile | Clinical Specimen |
|---|---|---|---|
| S. aureus | ATCC 6538 | Standard strain | |
| S. aureus | ATCC 25923 | Standard strain | |
| S. epidermidis | ATCC 12228 | Standard strain | |
| S. epidermidis | ATCC 35984 | Standard strain | |
| S. aureus | SA1 | Methicillin-, macrolides-, fluoroquinolones-resistant | Nasal swab |
| S. aureus | SA2 | Methicillin-susceptible, aminoglycoside-resistant, inducible MLSb phenotype | Pharyngeal swab |
| S. aureus | SA3 | Methicillin-susceptible, tetracycline-resistant, inducible MLSb phenotype | Blood culture |
| S. epidermidis | SE1 | Multi-sensitive | Venous Catheter |
| S. epidermidis | SE2 | Methicillin-, fluoroquinolones-, aminoglycosides-, rifamicins-, tetracycline-resistant, inducible MLSb phenotype | Blood culture |
| S. epidermidis | SE3 | Fluoroquinolones, lincosamides-, macrolides-, aminoglycosides-, rifamicins-, oxazolidinones-resistant | Blood culture |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palma, F.; Chianese, A.; Panico, E.; Greco, G.; Fusco, A.; Savio, V.; Ruocco, E.; Monti, A.; Doti, N.; Zannella, C.; et al. Oreoch-1: A Peptide from Oreochromis niloticus as a Potential Tool against Staphylococci. Pathogens 2023, 12, 1188. https://doi.org/10.3390/pathogens12101188
Palma F, Chianese A, Panico E, Greco G, Fusco A, Savio V, Ruocco E, Monti A, Doti N, Zannella C, et al. Oreoch-1: A Peptide from Oreochromis niloticus as a Potential Tool against Staphylococci. Pathogens. 2023; 12(10):1188. https://doi.org/10.3390/pathogens12101188
Chicago/Turabian StylePalma, Francesca, Annalisa Chianese, Erica Panico, Giuseppe Greco, Alessandra Fusco, Vittoria Savio, Eleonora Ruocco, Alessandra Monti, Nunzianna Doti, Carla Zannella, and et al. 2023. "Oreoch-1: A Peptide from Oreochromis niloticus as a Potential Tool against Staphylococci" Pathogens 12, no. 10: 1188. https://doi.org/10.3390/pathogens12101188