
In Silico Evaluation of CRISPR-Based Assays for Effective Detection of SARS-CoV-2

 ,
,
Abstract
:1. Introduction
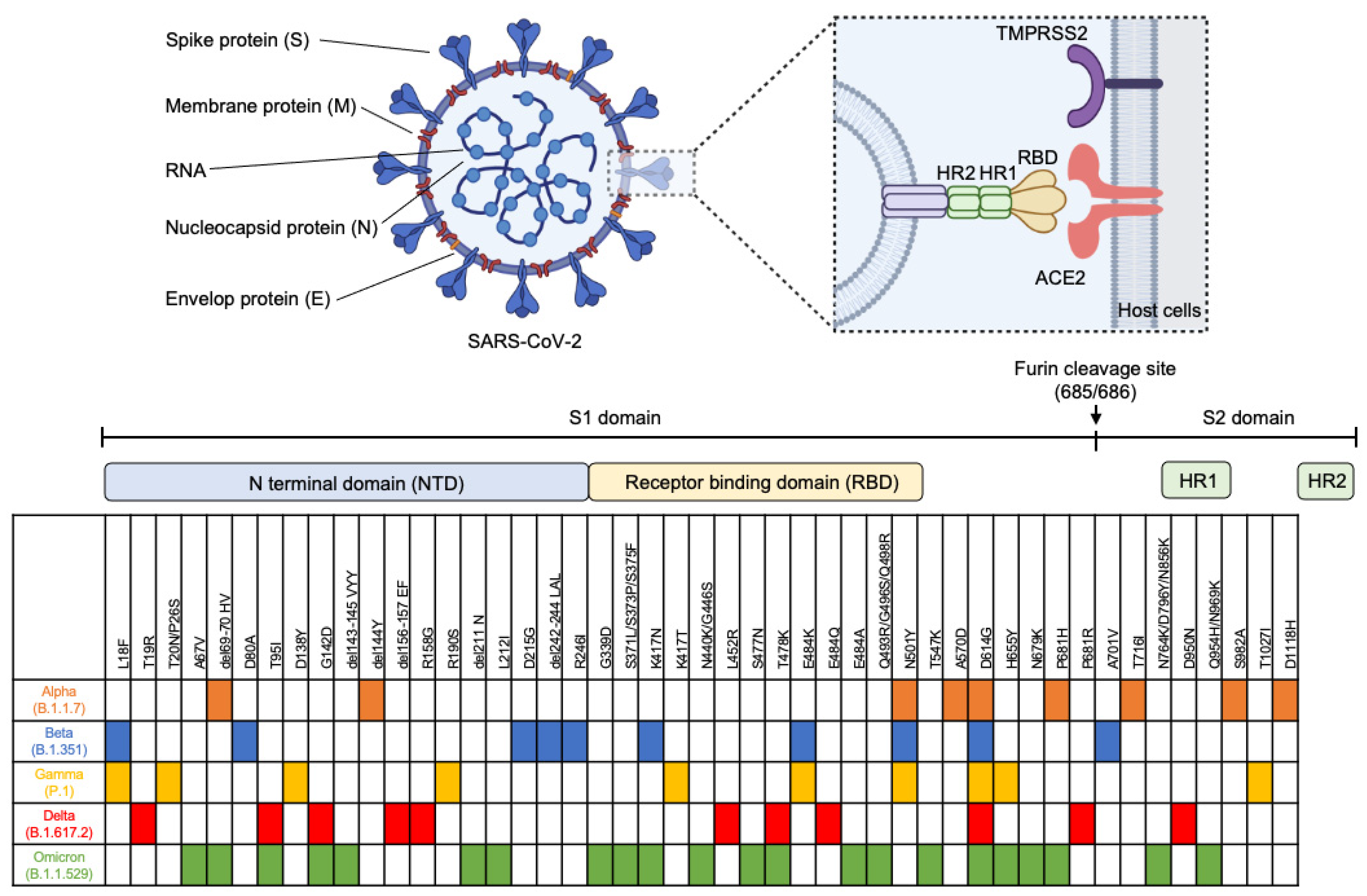
2. Mutations in SARS-CoV-2 Variants
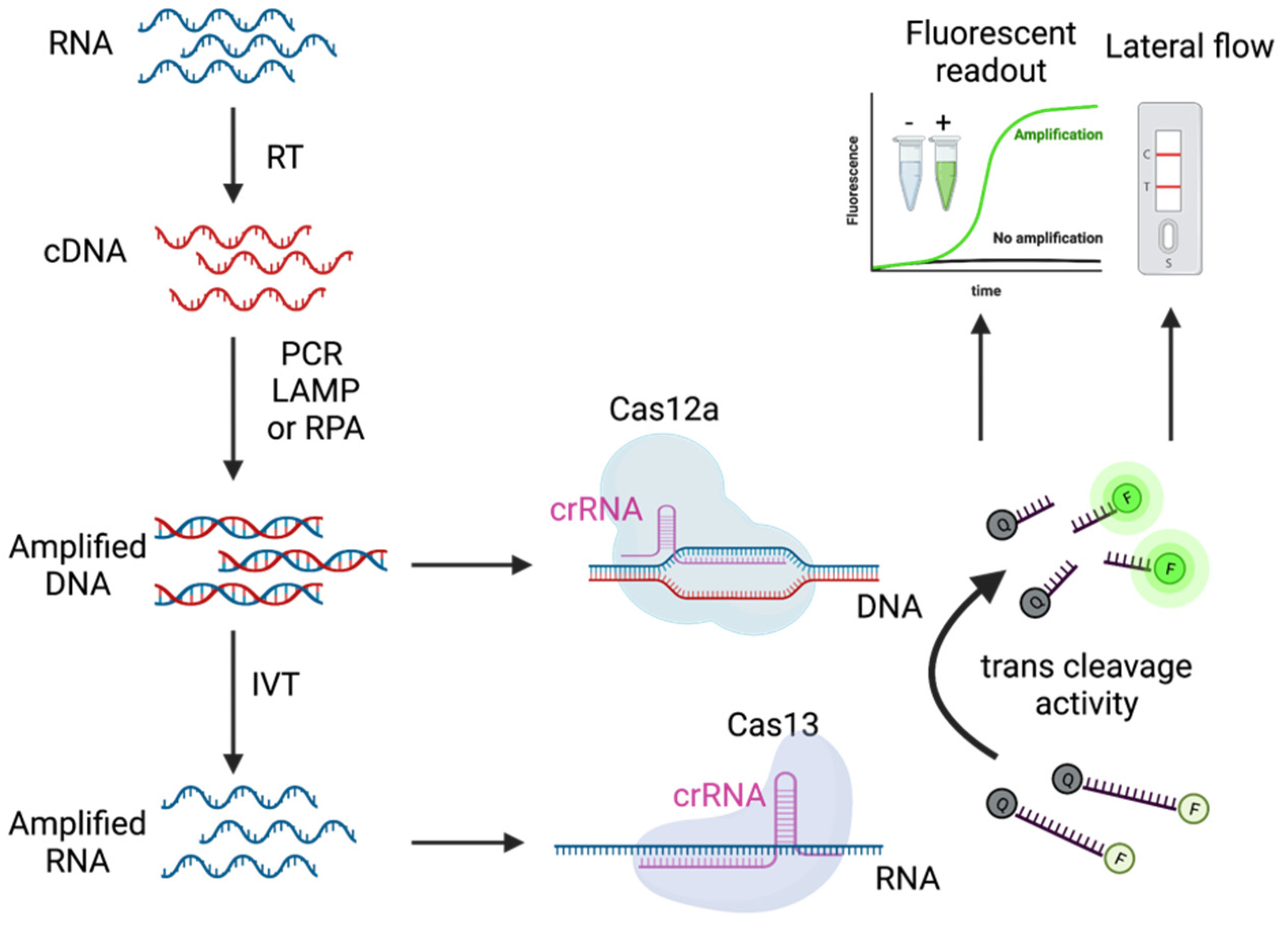
3. CRISPR-Cas Detection System
4. Mismatches in the Amplification Step and CRISPR-Cas Detection Step
5. Mismatches in Published CRISPR-Cas Detection System
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 18 July 2022).
- World Health Organization. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 18 July 2022).
- Plante, J.A.; Liu, Y.; Liu, J.; Xia, H.; Johnson, B.A.; Lokugamage, K.G.; Zhang, X.; Muruato, A.E.; Zou, J.; Fontes-Garfias, C.R.; et al. Spike Mutation D614G Alters SARS-CoV-2 Fitness. Nature 2021, 592, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic Acid Detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef]
- Chen, J.S.; Ma, E.; Harrington, L.B.; da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a Target Binding Unleashes Indiscriminate Single-Stranded DNase Activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Letko, M.; Marzi, A.; Munster, V. Functional Assessment of Cell Entry and Receptor Usage for SARS-CoV-2 and Other Lineage B Betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.D.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C.; et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 2020, 182, 1295–1310.e20. [Google Scholar] [CrossRef]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.-Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y.; et al. Adaptation of SARS-CoV-2 in BALB/c Mice for Testing Vaccine Efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef]
- Greaney, A.J.; Loes, A.N.; Crawford, K.H.D.; Starr, T.N.; Malone, K.D.; Chu, H.Y.; Bloom, J.D. Comprehensive Mapping of Mutations in the SARS-CoV-2 Receptor-Binding Domain That Affect Recognition by Polyclonal Human Plasma Antibodies. Cell Host Microbe 2021, 29, 463–476.e6. [Google Scholar] [CrossRef]
- Motozono, C.; Toyoda, M.; Zahradnik, J.; Saito, A.; Nasser, H.; Tan, T.S.; Ngare, I.; Kimura, I.; Uriu, K.; Kosugi, Y.; et al. SARS-CoV-2 Spike L452R Variant Evades Cellular Immunity and Increases Infectivity. Cell Host Microbe 2021, 29, 1124–1136.e11. [Google Scholar] [CrossRef]
- Khairnar, P.; Soni, M.; Handa, M.; Riadi, Y.; Kesharwani, P.; Shukla, R. Recent Highlights on Omicron as a New SARS-COVID-19 Variant: Evolution, Genetic Mutation, and Future Perspectives. J. Drug Target 2022, 30, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.; Wu, X.; Jiang, W.; Marraffini, L.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Y.; Cheng, Q.-X.; Wang, J.-M.; Li, X.-Y.; Zhang, Z.-L.; Gao, S.; Cao, R.-B.; Zhao, G.-P.; Wang, J. CRISPR-Cas12a-Assisted Nucleic Acid Detection. Cell Discov. 2018, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Guo, L.; Cui, T.; Wang, X.-G.; Xu, K.; Gao, Q.; Zhou, Q.; Li, W. CDetection: CRISPR-Cas12b-Based DNA Detection with Sub-Attomolar Sensitivity and Single-Base Specificity. Genome Biol. 2019, 20, 132. [Google Scholar] [CrossRef]
- Huang, J.; Brieba, L.G.; Sousa, R. Misincorporation by Wild-Type and Mutant T7 RNA Polymerases: Identification of Interactions That Reduce Misincorporation Rates by Stabilizing the Catalytically Incompetent Open Conformation. Biochemistry 2000, 39, 11571–11580. [Google Scholar] [CrossRef]
- Ramachandran, A.; Santiago, J.G. CRISPR Enzyme Kinetics for Molecular Diagnostics. Anal. Chem. 2021, 93, 7456–7464. [Google Scholar] [CrossRef]
- Santiago-Frangos, A.; Hall, L.N.; Nemudraia, A.; Nemudryi, A.; Krishna, P.; Wiegand, T.; Wilkinson, R.A.; Snyder, D.T.; Hedges, J.F.; Cicha, C.; et al. Intrinsic Signal Amplification by Type III CRISPR-Cas Systems Provides a Sequence-Specific SARS-CoV-2 Diagnostic. Cell Rep. Med. 2021, 2, 100319. [Google Scholar] [CrossRef]
- Zou, Y.; Mason, M.G.; Botella, J.R. Evaluation and Improvement of Isothermal Amplification Methods for Point-of-Need Plant Disease Diagnostics. PLoS ONE 2020, 15, e0235216. [Google Scholar] [CrossRef]
- Ayyadevara, S.; Thaden, J.J.; Shmookler Reis, R.J. Discrimination of Primer 3’-Nucleotide Mismatch by Taq DNA Polymerase during Polymerase Chain Reaction. Anal. Biochem. 2000, 284, 11–18. [Google Scholar] [CrossRef]
- Kwok, S.; Kellogg, D.E.; McKinney, N.; Spasic, D.; Goda, L.; Levenson, C.; Sninsky, J.J. Effects of Primer-Template Mismatches on the Polymerase Chain Reaction: Human Immunodeficiency Virus Type 1 Model Studies. Nucleic Acids Res. 1990, 18, 999–1005. [Google Scholar] [CrossRef]
- Ooi, K.H.; Liu, M.M.; Tay, J.W.D.; Teo, S.Y.; Kaewsapsak, P.; Jin, S.; Lee, C.K.; Hou, J.; Maurer-Stroh, S.; Lin, W.; et al. An Engineered CRISPR-Cas12a Variant and DNA-RNA Hybrid Guides Enable Robust and Rapid COVID-19 Testing. Nat. Commun. 2021, 12, 1739. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 Is a Single-Component Programmable RNA-Guided RNA-Targeting CRISPR Effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [PubMed]
- Wessels, H.-H.; Méndez-Mancilla, A.; Guo, X.; Legut, M.; Daniloski, Z.; Sanjana, N.E. Massively Parallel Cas13 Screens Reveal Principles for Guide RNA Design. Nat. Biotechnol. 2020, 38, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, J.; Zhou, X. A CRISPR-Based and Post-Amplification Coupled SARS-CoV-2 Detection with a Portable Evanescent Wave Biosensor. Biosens. Bioelectron. 2021, 190, 113418. [Google Scholar] [CrossRef]
- Mahas, A.; Wang, Q.; Marsic, T.; Mahfouz, M.M. A Novel Miniature CRISPR-Cas13 System for SARS-CoV-2 Diagnostics. ACS Synth. Biol. 2021, 10, 2541–2551. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, J.; Ren, L.; Jiang, W.; Wang, M.; Zhuang, L.; Zheng, Q.; Yang, R.; Zeng, Y.; Luu, L.D.W.; et al. A One-Step, One-Pot CRISPR Nucleic Acid Detection Platform (CRISPR-Top): Application for the Diagnosis of COVID-19. Talanta 2021, 233, 122591. [Google Scholar] [CrossRef] [PubMed]
- Azmi, I.; Faizan, M.I.; Kumar, R.; Raj Yadav, S.; Chaudhary, N.; Kumar Singh, D.; Butola, R.; Ganotra, A.; Datt Joshi, G.; Deep Jhingan, G.; et al. A Saliva-Based RNA Extraction-Free Workflow Integrated with Cas13a for SARS-CoV-2 Detection. Front. Cell Infect. Microbiol. 2021, 11, 632646. [Google Scholar] [CrossRef]
- Rauch, J.N.; Valois, E.; Solley, S.C.; Braig, F.; Lach, R.S.; Audouard, M.; Ponce-Rojas, J.C.; Costello, M.S.; Baxter, N.J.; Kosik, K.S.; et al. A Scalable, Easy-to-Deploy Protocol for Cas13-Based Detection of SARS-CoV-2 Genetic Material. J. Clin. Microbiol. 2021, 59, e02402-20. [Google Scholar] [CrossRef]
- Ma, L.; Yin, L.; Li, X.; Chen, S.; Peng, L.; Liu, G.; Ye, S.; Zhang, W.; Man, S. A Smartphone-Based Visual Biosensor for CRISPR-Cas Powered SARS-CoV-2 Diagnostics. Biosens. Bioelectron. 2022, 195, 113646. [Google Scholar] [CrossRef]
- Ning, B.; Yu, T.; Zhang, S.; Huang, Z.; Tian, D.; Lin, Z.; Niu, A.; Golden, N.; Hensley, K.; Threeton, B.; et al. A Smartphone-Read Ultrasensitive and Quantitative Saliva Test for COVID-19. Sci. Adv. 2021, 7, eabe3703. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Macaluso, N.C.; Pizzano, B.L.M.; Cash, M.N.; Spacek, J.; Karasek, J.; Miller, M.R.; Lednicky, J.A.; Dinglasan, R.R.; Salemi, M.; et al. A Thermostable Cas12b from Brevibacillus Leverages One-Pot Discrimination of SARS-CoV-2 Variants of Concern. EBioMedicine 2022, 77, 103926. [Google Scholar] [CrossRef] [PubMed]
- Fozouni, P.; Son, S.; Díaz de León Derby, M.; Knott, G.J.; Gray, C.N.; D’Ambrosio, M.V.; Zhao, C.; Switz, N.A.; Kumar, G.R.; Stephens, S.I.; et al. Amplification-Free Detection of SARS-CoV-2 with CRISPR-Cas13a and Mobile Phone Microscopy. Cell 2021, 184, 323–333.e9. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Teng, P.; Xiao, W.; He, G.; Song, Q.; Zhang, Y.; Peng, B.; Li, G.; Hu, L.; Cao, D.; et al. Application of the Amplification-Free SERS-Based CRISPR/Cas12a Platform in the Identification of SARS-CoV-2 from Clinical Samples. J. Nanobiotechnol. 2021, 19, 273. [Google Scholar] [CrossRef] [PubMed]
- Patchsung, M.; Jantarug, K.; Pattama, A.; Aphicho, K.; Suraritdechachai, S.; Meesawat, P.; Sappakhaw, K.; Leelahakorn, N.; Ruenkam, T.; Wongsatit, T.; et al. Clinical Validation of a Cas13-Based Assay for the Detection of SARS-CoV-2 RNA. Nat. Biomed. Eng. 2020, 4, 1140–1149. [Google Scholar] [CrossRef]
- Chen, Y.; Shi, Y.; Chen, Y.; Yang, Z.; Wu, H.; Zhou, Z.; Li, J.; Ping, J.; He, L.; Shen, H.; et al. Contamination-Free Visual Detection of SARS-CoV-2 with CRISPR/Cas12a: A Promising Method in the Point-of-Care Detection. Biosens. Bioelectron. 2020, 169, 112642. [Google Scholar] [CrossRef]
- Liu, S.; Huang, M.; Xu, Y.; Kang, J.; Ye, S.; Liu, S.; Wang, Z.; Liu, H.; Yu, J.; Hu, K.; et al. CRISPR/Cas12a Technology Combined with RT-ERA for Rapid and Portable SARS-CoV-2 Detection. Virol. Sin. 2021, 36, 1083–1087. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, J.; Pang, B.; Zhang, H.; Le, X.C. CRISPR/Cas12a-Mediated Gold Nanoparticle Aggregation for Colorimetric Detection of SARS-CoV-2. Chem. Commun. 2021, 57, 6871–6874. [Google Scholar] [CrossRef]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR-Cas12-Based Detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef]
- Talwar, C.S.; Park, K.-H.; Ahn, W.-C.; Kim, Y.-S.; Kwon, O.S.; Yong, D.; Kang, T.; Woo, E. Detection of Infectious Viruses Using CRISPR-Cas12-Based Assay. Biosensors 2021, 11, 301. [Google Scholar] [CrossRef]
- Jiang, Y.; Hu, M.; Liu, A.-A.; Lin, Y.; Liu, L.; Yu, B.; Zhou, X.; Pang, D.-W. Detection of SARS-CoV-2 by CRISPR/Cas12a-Enhanced Colorimetry. ACS Sens. 2021, 6, 1086–1093. [Google Scholar] [CrossRef]
- Mayuramart, O.; Nimsamer, P.; Rattanaburi, S.; Chantaravisoot, N.; Khongnomnan, K.; Chansaenroj, J.; Puenpa, J.; Suntronwong, N.; Vichaiwattana, P.; Poovorawan, Y.; et al. Detection of Severe Acute Respiratory Syndrome Coronavirus 2 and Influenza Viruses Based on CRISPR-Cas12a. Exp. Biol. Med. 2021, 246, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Wang, X.; Wang, Y.; Dang, L.; Liu, X.; Ma, X.; Chi, T.; Wang, X.; Zhao, Q.; Yang, G.; et al. Detection of the SARS-CoV-2 D614G Mutation Using Engineered Cas12a Guide RNA. Biotechnol. J. 2021, 16, e2100040. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Zeng, W.; Yang, M.; Chen, W.; Ren, L.; Ai, J.; Wu, J.; Liao, Y.; Gou, X.; Li, Y.; et al. Development and Evaluation of a Rapid CRISPR-Based Diagnostic for COVID-19. PLoS Pathog. 2020, 16, e1008705. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, D.; Lin, H.; Chen, D.; Sun, J.; Xie, Y.; Wang, X.; Ma, P.; Nie, Y.; Mei, H.; et al. Development of a Broadly Applicable Cas12a-Linked Beam Unlocking Reaction for Sensitive and Specific Detection of Respiratory Pathogens Including SARS-CoV-2. ACS Chem. Biol. 2021, 16, 491–500. [Google Scholar] [CrossRef]
- Brogan, D.J.; Chaverra-Rodriguez, D.; Lin, C.P.; Smidler, A.L.; Yang, T.; Alcantara, L.M.; Antoshechkin, I.; Liu, J.; Raban, R.R.; Belda-Ferre, P.; et al. Development of a Rapid and Sensitive CasRx-Based Diagnostic Assay for SARS-CoV-2. ACS Sens. 2021, 6, 3957–3966. [Google Scholar] [CrossRef]
- Park, J.S.; Hsieh, K.; Chen, L.; Kaushik, A.; Trick, A.Y.; Wang, T.-H. Digital CRISPR/Cas-Assisted Assay for Rapid and Sensitive Detection of SARS-CoV-2. Adv. Sci. 2021, 8, 2003564. [Google Scholar] [CrossRef]
- Ramachandran, A.; Huyke, D.A.; Sharma, E.; Sahoo, M.K.; Huang, C.; Banaei, N.; Pinsky, B.A.; Santiago, J.G. Electric Field-Driven Microfluidics for Rapid CRISPR-Based Diagnostics and Its Application to Detection of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 29518–29525. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Smith, B.M.; Jain, P.K. Enhancement of Trans-Cleavage Activity of Cas12a with Engineered CrRNA Enables Amplified Nucleic Acid Detection. Nat. Commun. 2020, 11, 4906. [Google Scholar] [CrossRef]
- Lee, C.Y.; Degani, I.; Cheong, J.; Lee, J.-H.; Choi, H.-J.; Cheon, J.; Lee, H. Fluorescence Polarization System for Rapid COVID-19 Diagnosis. Biosens. Bioelectron. 2021, 178, 113049. [Google Scholar] [CrossRef]
- Li, Z.; Ding, X.; Yin, K.; Avery, L.; Ballesteros, E.; Liu, C. Instrument-Free, CRISPR-Based Diagnostics of SARS-CoV-2 Using Self-Contained Microfluidic System. Biosens. Bioelectron. 2022, 199, 113865. [Google Scholar] [CrossRef]
- Ali, Z.; Aman, R.; Mahas, A.; Rao, G.S.; Tehseen, M.; Marsic, T.; Salunke, R.; Subudhi, A.K.; Hala, S.M.; Hamdan, S.M.; et al. ISCAN: An RT-LAMP-Coupled CRISPR-Cas12 Module for Rapid, Sensitive Detection of SARS-CoV-2. Virus Res. 2020, 288, 198129. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Xu, J.; Liu, Y.; Peng, H.; Feng, W.; Cao, Y.; Wu, J.; Xiao, H.; Pabbaraju, K.; Tipples, G.; et al. Isothermal Amplification and Ambient Visualization in a Single Tube for the Detection of SARS-CoV-2 Using Loop-Mediated Amplification and CRISPR Technology. Anal. Chem. 2020, 92, 16204–16212. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Meng, Q.; Sun, B.; Zhao, B.; Dang, L.; Zhong, M.; Liu, S.; Xu, H.; Mei, H.; Liu, J.; et al. MeCas12a, a Highly Sensitive and Specific System for COVID-19 Detection. Adv. Sci. 2020, 7, 2001300. [Google Scholar] [CrossRef] [PubMed]
- de Puig, H.; Lee, R.A.; Najjar, D.; Tan, X.; Soeknsen, L.R.; Angenent-Mari, N.M.; Donghia, N.M.; Weckman, N.E.; Ory, A.; Ng, C.F.; et al. Minimally Instrumented SHERLOCK (MiSHERLOCK) for CRISPR-Based Point-of-Care Diagnosis of SARS-CoV-2 and Emerging Variants. Sci. Adv. 2021, 7, abh2944. [Google Scholar] [CrossRef]
- Sun, Y.; Yu, L.; Liu, C.; Ye, S.; Chen, W.; Li, D.; Huang, W. One-Tube SARS-CoV-2 Detection Platform Based on RT-RPA and CRISPR/Cas12a. J. Transl. Med. 2021, 19, 74. [Google Scholar] [CrossRef]
- Chen, F.-E.; Lee, P.-W.; Trick, A.Y.; Park, J.S.; Chen, L.; Shah, K.; Mostafa, H.; Carroll, K.C.; Hsieh, K.; Wang, T.-H. Point-of-Care CRISPR-Cas-Assisted SARS-CoV-2 Detection in an Automated and Portable Droplet Magnetofluidic Device. Biosens. Bioelectron. 2021, 190, 113390. [Google Scholar] [CrossRef]
- Joung, J.; Ladha, A.; Saito, M.; Segel, M.; Bruneau, R.; Huang, M.W.; Kim, N.G.; Yu, X.; Li, J.; Walker, B.D.; et al. Point-of-Care Testing for COVID-19 Using SHERLOCK Diagnostics. MedRxiv, 2020; Preprint. [Google Scholar] [CrossRef]
- Wang, X.; Zhong, M.; Liu, Y.; Ma, P.; Dang, L.; Meng, Q.; Wan, W.; Ma, X.; Liu, J.; Yang, G.; et al. Rapid and Sensitive Detection of COVID-19 Using CRISPR/Cas12a-Based Detection with Naked Eye Readout, CRISPR/Cas12a-NER. Sci. Bull. 2020, 65, 1436–1439. [Google Scholar] [CrossRef]
- Tsou, J.-H.; Liu, H.; Stass, S.A.; Jiang, F. Rapid and Sensitive Detection of SARS-CoV-2 Using Clustered Regularly Interspaced Short Palindromic Repeats. Biomedicines 2021, 9, 239. [Google Scholar] [CrossRef]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Singh, J.; Streithorst, J.; Granados, A.; Sotomayor-Gonzalez, A.; Zorn, K.; Gopez, A.; et al. Rapid Detection of 2019 Novel Coronavirus SARS-CoV-2 Using a CRISPR-Based DETECTR Lateral Flow Assay. MedRxiv, 2020; Preprint. [Google Scholar] [CrossRef]
- Xiong, D.; Dai, W.; Gong, J.; Li, G.; Liu, N.; Wu, W.; Pan, J.; Chen, C.; Jiao, Y.; Deng, H.; et al. Rapid Detection of SARS-CoV-2 with CRISPR-Cas12a. PLoS Biol. 2020, 18, e3000978. [Google Scholar] [CrossRef]
- Schermer, B.; Fabretti, F.; Damagnez, M.; di Cristanziano, V.; Heger, E.; Arjune, S.; Tanner, N.A.; Imhof, T.; Koch, M.; Ladha, A.; et al. Rapid SARS-CoV-2 Testing in Primary Material Based on a Novel Multiplex RT-LAMP Assay. PLoS ONE 2020, 15, e0238612. [Google Scholar] [CrossRef]
- Zhang, W.S.; Pan, J.; Li, F.; Zhu, M.; Xu, M.; Zhu, H.; Yu, Y.; Su, G. Reverse Transcription Recombinase Polymerase Amplification Coupled with CRISPR-Cas12a for Facile and Highly Sensitive Colorimetric SARS-CoV-2 Detection. Anal. Chem. 2021, 93, 4126–4133. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Sun, X.; Wang, X.; Liang, C.; Jiang, H.; Gao, Q.; Dai, M.; Qu, B.; Fang, S.; Mao, Y.; et al. SARS-CoV-2 Detection with CRISPR Diagnostics. Cell Discov. 2020, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Venzor, A.; Rueda-Zarazua, B.; Marquez-Garcia, E.; Maldonado, V.; Moncada-Morales, A.; Olivera, H.; Lopez, I.; Zuñiga, J.; Melendez-Zajgla, J. SARS-CoV-2 Direct Detection Without RNA Isolation with Loop-Mediated Isothermal Amplification (LAMP) and CRISPR-Cas12. Front. Med. 2021, 8, 627679. [Google Scholar] [CrossRef]
- Silva, F.S.R.; Erdogmus, E.; Shokr, A.; Kandula, H.; Thirumalaraju, P.; Kanakasabapathy, M.K.; Hardie, J.M.; Pacheco, L.G.C.; Li, J.Z.; Kuritzkes, D.R.; et al. SARS-CoV-2 RNA Detection by a Cellphone-Based Amplification-Free System with CRISPR/CAS-Dependent Enzymatic (CASCADE) Assay. Adv. Mater. Technol. 2021, 6, 2100602. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dong, X.; Wang, Y.; Yang, L.; Cai, K.; Zhang, X.; Kou, Z.; He, L.; Sun, S.; Li, T.; et al. Sensitive and Easy-Read CRISPR Strip for COVID-19 Rapid Point-of-Care Testing. CRISPR J. 2021, 4, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, M.; Liu, C.; Chen, J.; Luo, X.; Xue, Y.; Liang, Q.; Zhou, L.; Tao, Y.; Li, M.; et al. Sensitive and Rapid On-Site Detection of SARS-CoV-2 Using a Gold Nanoparticle-Based High-Throughput Platform Coupled with CRISPR/Cas12-Assisted RT-LAMP. Sens. Actuators Chem. 2021, 345, 130411. [Google Scholar] [CrossRef]
- Huang, Z.; Ning, B.; Yang, H.S.; Youngquist, B.M.; Niu, A.; Lyon, C.J.; Beddingfield, B.J.; Fears, A.C.; Monk, C.H.; Murrell, A.E.; et al. Sensitive Tracking of Circulating Viral RNA through All Stages of SARS-CoV-2 Infection. J. Clin. Investig. 2021, 131, e146031. [Google Scholar] [CrossRef]
- Arizti-Sanz, J.; Freije, C.A.; Stanton, A.C.; Petros, B.A.; Boehm, C.K.; Siddiqui, S.; Shaw, B.M.; Adams, G.; Kosoko-Thoroddsen, T.-S.F.; Kemball, M.E.; et al. Streamlined Inactivation, Amplification, and Cas13-Based Detection of SARS-CoV-2. Nat. Commun. 2020, 11, 5921. [Google Scholar] [CrossRef]
- Huang, Z.; Tian, D.; Liu, Y.; Lin, Z.; Lyon, C.J.; Lai, W.; Fusco, D.; Drouin, A.; Yin, X.; Hu, T.; et al. Ultra-Sensitive and High-Throughput CRISPR-Powered COVID-19 Diagnosis. Biosens. Bioelectron. 2020, 164, 112316. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, X.; Li, S.; Luo, W.; Zhang, X.; Wang, C.; Chen, Q.; Yu, S.; Tai, J.; Wang, Y. Rapid, Ultrasensitive, and Highly Specific Diagnosis of COVID-19 by CRISPR-Based Detection. ACS Sens. 2021, 6, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, R.; Peñaranda, K.; Mendoza-Rojas, G.; Nakamoto, J.A.; Dueñas, E.; Alvarez, D.; Adaui, V.; Milón, P. UnCovid: A Versatile, Low-Cost, and Open-Source Protocol for SARS-CoV-2 RNA Detection. STAR Protoc. 2021, 2, 100878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, W.; Zhao, W.; Si, Y.; Chen, N.; Chen, X.; Zhang, X.; Fan, L.; Sui, G. Universally Stable and Precise CRISPR-LAMP Detection Platform for Precise Multiple Respiratory Tract Virus Diagnosis Including Mutant SARS-CoV-2 Spike N501Y. Anal. Chem. 2021, 93, 16184–16193. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, R.; Peñaranda, K.; Mendoza-Rojas, G.; Nakamoto, J.A.; Martins-Luna, J.; del Valle-Mendoza, J.; Adaui, V.; Milón, P. Unlocking SARS-CoV-2 Detection in Low- and Middle-Income Countries. Cell Rep. Methods 2021, 1, 100093. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Title | Target | Amp | Primers Match for Variants of Concern | Cas | crRNA | Remark | Ref | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| α | β | γ | δ | O | |||||||
| A CRISPR-based and post-amplification coupled SARS-CoV-2 detection with a portable evanescent wave biosensor | S | - | - | - | - | - | - | Cas13a | ✓ | [26] | |
| N | - | - | - | - | - | - | Cas13a | ✓ | |||
| Orf1ab | - | - | - | - | - | - | Cas13a | MM1/MM2 | MM1: | ||
| β, γ, δ, O | |||||||||||
| MM2: α | |||||||||||
| A Novel Miniature CRISPR-Cas13 System for SARS-CoV-2 Diagnostics | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | mCas13 | ✓ | [27] | |
| A one-step, one-pot CRISPR nucleic acid detection platform (CRISPR-top): Application for the diagnosis of COVID-19 | Orf1ab | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12b | ✓ | [28] | |
| A Saliva-Based RNA Extraction-Free Workflow Integrated With Cas13a for SARS-CoV-2 Detection | N | LAMP | ✓ | ✓ | ✓ | MM1 | ✓ | Cas12b | ✓ | [29] | |
| S | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| Orf1ab | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| A Scalable, Easy-to-Deploy Protocol for Cas13-Based Detection of SARS-CoV-2 Genetic Material | N1 | PCR | ✓ | MM1 | ✓ | ✓ | ✓ | Cas13a | MM1 | Omicron | [30] |
| N2 | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| N3 | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| A smartphone-based visual biosensor for CRISPR-Cas powered SARS-CoV-2 diagnostics | N | PCR | MM4 | MM1 | MM3 | MM1 | MM3 | Cas12a | ✓ | [31] | |
| A smartphone-read ultrasensitive and quantitative saliva test for COVID-19 | Orf1ab | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [32] | |
| N | PCR | MM4 | MM1 | MM3 | MM1 | MM3 | Cas12a | ✓ | |||
| A Thermostable Cas12b from Brevibacillus Leverages One-pot Detection of SARS-CoV-2 Variants of Concern | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12b | ✓ | [33] | |
| Amplification-free detection of SARS-CoV-2 with CRISPR-Cas13a and mobile phone microscopy | N | - | - | - | - | - | - | Cas13a | ✓ | All three crRNAs are perfect match | [34] |
| An engineered CRISPR-Cas12a variant and DNA-RNA hybrid guides enable robust and rapid COVID-19 testing | S | LAMP | ✓ | MM1 | ✓ | ✓ | Del3, Ins9 | Cas12a | ✓ | [23] | |
| Application of the amplification-free SERS-based CRISPR/Cas12a platform in the identification of SARS-CoV-2 from clinical samples | N | - | - | - | - | - | - | Cas12a | ✓ | [35] | |
| Clinical validation of a Cas13-based assay for the detection of SARS-CoV-2 RNA | S | RPA | ✓ | DEL9 | ✓ | ✓ | ✓ | Cas13a | ✓ | [36] | |
| Orf1ab | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | DEL3 | Omicron | ||
| Orf1b | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| N | RPA | MM1 | ✓ | ✓ | ✓ | ✓ | Cas13a | MM1 | Delta | ||
| Contamination-free visual detection of SARS-CoV-2 with CRISPR/Cas12a: A promising method in the point-of-care detection | E | LAMP | ✓ | ✓ | ✓ | ✓ | MM1 | Cas12a | ✓ | [37] | |
| Orf1ab | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| N | LAMP | MM1 | ✓ | ✓ | ✓ | ✓ | Cas12a | MM1 | Alpha | ||
| CRISPR/Cas12a Technology Combined with RT-ERA for Rapid and Portable SARS-CoV-2 Detection | N | ERA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [38] | |
| Orf1ab | ERA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| CRISPR/Cas12a-mediated gold nanoparticle aggregation for colorimetric detection of SARS-CoV-2 | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [39] | |
| E | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| CRISPR-Cas12-based detection of SARS-CoV-2 | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [40] | |
| E | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| Detection of Infectious Viruses Using CRISPR-Cas12-Based Assay | S | RPA | ✓ | MM1 | MM1 | MM1 | MM3 | Cas12a | ✓ | [41] | |
| Detection of SARS-CoV-2 by CRISPR/Cas12a-Enhanced Colorimetry | E | RPA | ✓ | MM1 | ✓ | ✓ | MM1 | Cas12a | ✓ | [42] | |
| Orf1ab | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| N1 | RPA | MM1 | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| N2 | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| Detection of severe acute respiratory syndrome coronavirus 2 and influenza viruses based on CRISPR-Cas12a | S1 | RPA | ✓ | ✓ | ✓ | MM1, DEL6 | MM1 | Cas12a | ✓ | [43] | |
| S2 | RPA | ✓ | ✓ | ✓ | MM1 | MM2 | Cas12a | MM3 | Omicron | ||
| Detection of the SARS-CoV-2 D614G mutation using engineered Cas12a guide RNA | E | RPA | ✓ | MM1 | ✓ | ✓ | MM1 | Cas12a | ✓ | [44] | |
| Development and evaluation of a rapid CRISPR-based diagnostic for COVID-19 | Orf1ab | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | [45] | |
| Development of a Broadly Applicable Cas12a-Linked Beam Unlocking Reaction for Sensitive and Specific Detection of Respiratory Pathogens Including SARS-CoV-2 | S | RPA | DEL6 | ✓ | ✓ | ✓ | MM1, DEL6 | Cas12a | ✓ | [46] | |
| Development of a Rapid and Sensitive CasRx-Based Diagnostic Assay for SARS-CoV-2 | S3 | RPA | MM1 | ✓ | ✓ | ✓ | ✓ | Cas13d | ✓ | [47] | |
| N1 | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13d | ✓ | |||
| Digital CRISPR/Cas-Assisted Assay for Rapid and Sensitive Detection of SARS-CoV-2 | N | RPA | MM4 | MM1 | MM5 | MM1 | MM3 | Cas12a | MM1 | Delta: One crRNA has MM. The other has PM. | [48] |
| Electric field-driven microfluidics for rapid CRISPR-based diagnostics and its application to detection of SARS-CoV-2 | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [49] | |
| E | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| Enhancement of trans-cleavage activity of Cas12a with engineered crRNA enables amplified nucleic acid detection | N1 | LAMP | MM1 | MM2 | ✓ | DEL6 | ✓ | Cas12a | ✓ | [50] | |
| N2 | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| Fluorescence polarization system for rapid COVID-19 diagnosis | N1 | RPA | MM4 | MM1 | MM5 | MM1 | MM3 | Cas12a | MM1 | Delta | [51] |
| N2 | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| Instrument-free, CRISPR-based diagnostics of SARS-CoV-2 using self-contained microfluidic system | N | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [52] | |
| iSCAN: An RT-LAMP-coupled CRISPR-Cas12 module for rapid, sensitive detection of SARS-CoV-2 | E | LAMP | ✓ | MM1 | ✓ | ✓ | ✓ | Cas12a | ✓ | [53] | |
| Cas12b | ✓ | ||||||||||
| Isothermal Amplification and Ambient Visualization in a Single Tube for the Detection of SARS-CoV-2 Using Loop-Mediated Amplification and CRISPR Technology | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [54] | |
| E | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| MeCas12a, a Highly Sensitive and Specific System for COVID-19 Detection | E | RPA | ✓ | MM1 | ✓ | ✓ | MM1 | Cas12a | ✓ | [55] | |
| Minimally instrumented SHERLOCK (miSHERLOCK) for CRISPR-based point-of-care diagnosis of SARS-CoV-2 and emerging variants | N | RPA | MM1 | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [56] | |
| One-tube SARS-CoV-2 detection platform based on RT-RPA and CRISPR/Cas12a | RdRp | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | MM1 | Delta | [57] |
| N | RPA | MM4 | MM1 | MM5 | MM1 | MM3 | Cas12a | ✓ | |||
| Point-of-care CRISPR-Cas-assisted SARS-CoV-2 detection in an automated and portable droplet magnetofluidic device | N | RPA | MM4 | MM1 | MM5 | MM1 | MM3 | Cas12a | MM1 | Delta | [58] |
| Point-of-care testing for COVID-19 using SHERLOCK diagnostics | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12b | ✓ | [59] | |
| Rapid and sensitive detection of COVID-19 using CRISPR/Cas12a-based detection with naked eye readout, CRISPR/Cas12a-NER | Orf1a | RPA | ✓ | ✓ | ✓ | ✓ | MM1 | Cas12a | ✓ | [60] | |
| Orf1b | RPA | ✓ | ✓ | ✓ | MM1 | ✓ | Cas12a | ✓ | |||
| E | RPA | ✓ | MM1 | ✓ | ✓ | MM1 | Cas12a | ✓ | |||
| N | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | MM1 | Alpha | ||
| Rapid and Sensitive Detection of SARS-CoV-2 Using Clustered Regularly Interspaced Short Palindromic Repeats | M | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [61] | |
| N2 | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| S2 | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | MM1 | Gamma, Omicron | ||
| Rapid Detection of 2019 Novel Coronavirus SARS-CoV-2 Using a CRISPR-based DETECTR Lateral Flow Assay | N2 | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [62] | |
| E | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| E | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| Rapid detection of SARS-CoV-2 with CRISPR-Cas12a | Orf1ab | LAMP | ✓ | ✓ | ✓ | MM1 | ✓ | Cas12a | ✓ | [63] | |
| N | LAMP | MM3 | MM1 | MM5 | MM1 | MM3 | Cas12a | ✓ | |||
| Rapid SARS-CoV-2 testing in primary material based on a novel multiplex RT-LAMP assay | Orf1a | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | [64] | |
| N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| ORF7a | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| ORF3a | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| Orf1ab | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | DEL3 | Omicron | ||
| S | RPA | ✓ | DEL9 | ✓ | ✓ | ✓ | Cas13a | ✓ | |||
| Reverse Transcription Recombinase Polymerase Amplification Coupled with CRISPR-Cas12a for Facile and Highly Sensitive Colorimetric SARS-CoV-2 Detection | Orf1ab | RPA | ✓ | ✓ | ✓ | ✓ | MM1 | Cas12a | ✓ | [65] | |
| N | RPA | MM4 | MM1 | MM3 | MM1 | MM3 | Cas12a | ✓ | |||
| SARS-CoV-2 detection with CRISPR diagnostics | RdRp | RAA | MM1 | ✓ | ✓ | MM1 | ✓ | Cas12b | ✓ | [66] | |
| SARS-CoV-2 Direct Detection Without RNA Isolation With Loop-Mediated Isothermal Amplification (LAMP) and CRISPR-Cas12 | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [67] | |
| SARS-CoV-2 RNA Detection by a Cellphone-Based Amplification-Free System with CRISPR/CAS-Dependent Enzymatic (CASCADE) Assay | Orf1ab | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [68] | |
| Sensitive and Easy-Read CRISPR Strip for COVID-19 Rapid Point-of-Care Testing | N | RAA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | [69] | |
| Sensitive and rapid on-site detection of SARS-CoV-2 using a gold nanoparticle-based high-throughput platform coupled with CRISPR/Cas12-assisted RT-LAMP | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [70] | |
| Sensitive tracking of circulating viral RNA through all stages of SARS-CoV-2 infection | Orf1ab | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [71] | |
| Streamlined inactivation, amplification, and Cas13-based detection of SARS-CoV-2 | Orf1ab | RPA | ✓ | ✓ | ✓ | ✓ | ✓ | Cas13a | ✓ | [72] | |
| Ultra-sensitive and high-throughput CRISPR-powered COVID-19 diagnosis | Orf1ab | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [73] | |
| N | PCR | MM4 | MM1 | MM3 | MM1 | MM3 | Cas12a | ✓ | |||
| Ultrasensitive and visual detection of SARS-CoV-2 using all-in-one dual CRISPR-Cas12a assay | N | RPA | MM4 | MM1 | MM5 | MM1 | MM3 | Cas12a | MM1 | Delta: use 3 crRNAs and one crRNA has one MM | [74] |
| UnCovid: A versatile, low-cost, and open-source protocol for SARS-CoV-2 RNA detection | Orf1ab | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [75] | |
| N | PCR | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| Universally Stable and Precise CRISPR-LAMP Detection Platform for Precise Multiple Respiratory Tract Virus Diagnosis Including Mutant SARS-CoV-2 Spike N501Y | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [76] | |
| RdRp | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
| S | LAMP | ✓ | ✓ | ✓ | MM1 | ✓ | Cas12a | MM1, MM3 | Alpha and Beta strains have MM1. Omicron strain has MM3. | ||
| Unlocking SARS-CoV-2 detection in low- and middle-income countries | N | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | [77] | |
| Orf1ab | LAMP | ✓ | ✓ | ✓ | ✓ | ✓ | Cas12a | ✓ | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaewsapsak, P.; Chantaravisoot, N.; Nimsamer, P.; Mayuramart, O.; Mankhong, S.; Payungporn, S. In Silico Evaluation of CRISPR-Based Assays for Effective Detection of SARS-CoV-2. Pathogens 2022, 11, 968. https://doi.org/10.3390/pathogens11090968
Kaewsapsak P, Chantaravisoot N, Nimsamer P, Mayuramart O, Mankhong S, Payungporn S. In Silico Evaluation of CRISPR-Based Assays for Effective Detection of SARS-CoV-2. Pathogens. 2022; 11(9):968. https://doi.org/10.3390/pathogens11090968
Chicago/Turabian StyleKaewsapsak, Pornchai, Naphat Chantaravisoot, Pattaraporn Nimsamer, Oraphan Mayuramart, Suwanan Mankhong, and Sunchai Payungporn. 2022. "In Silico Evaluation of CRISPR-Based Assays for Effective Detection of SARS-CoV-2" Pathogens 11, no. 9: 968. https://doi.org/10.3390/pathogens11090968