
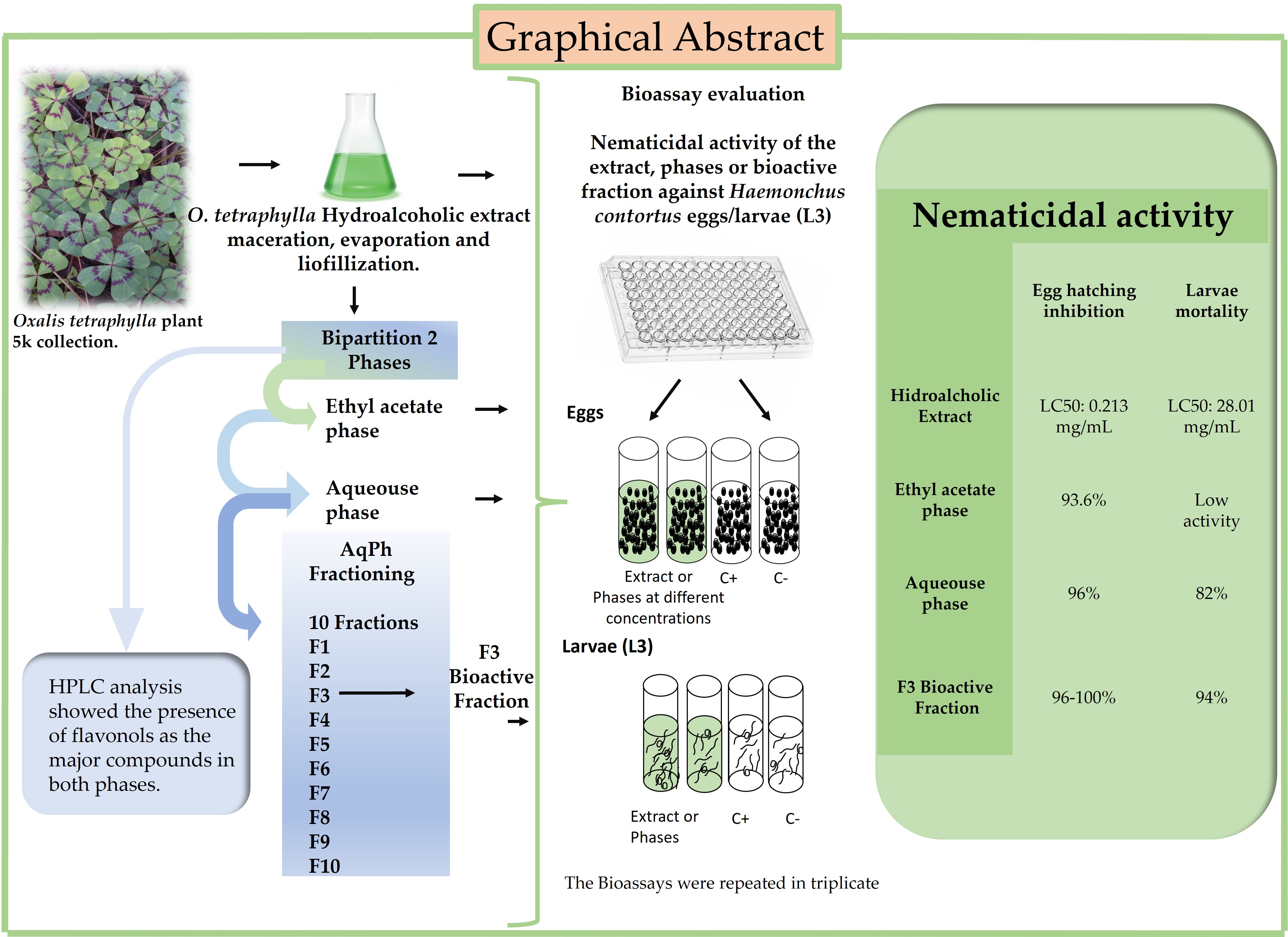
Oxalis tetraphylla (Class: Magnoliopsidae) Possess Flavonoid Phytoconstituents with Nematocidal Activity against Haemonchus contortus
 , , , , , ,
, , , , , ,
Abstract
:
1. Introduction
2. Results
2.1. Assessment and Identification SD = Standard Deviation of the Active Nematicidal Fractions from the Aqueous Phase of Oxalis tetraphylla against Haemonchus contortus Eggs and Larvae (L3)
2.2. Thin-Layer Chromatography Results
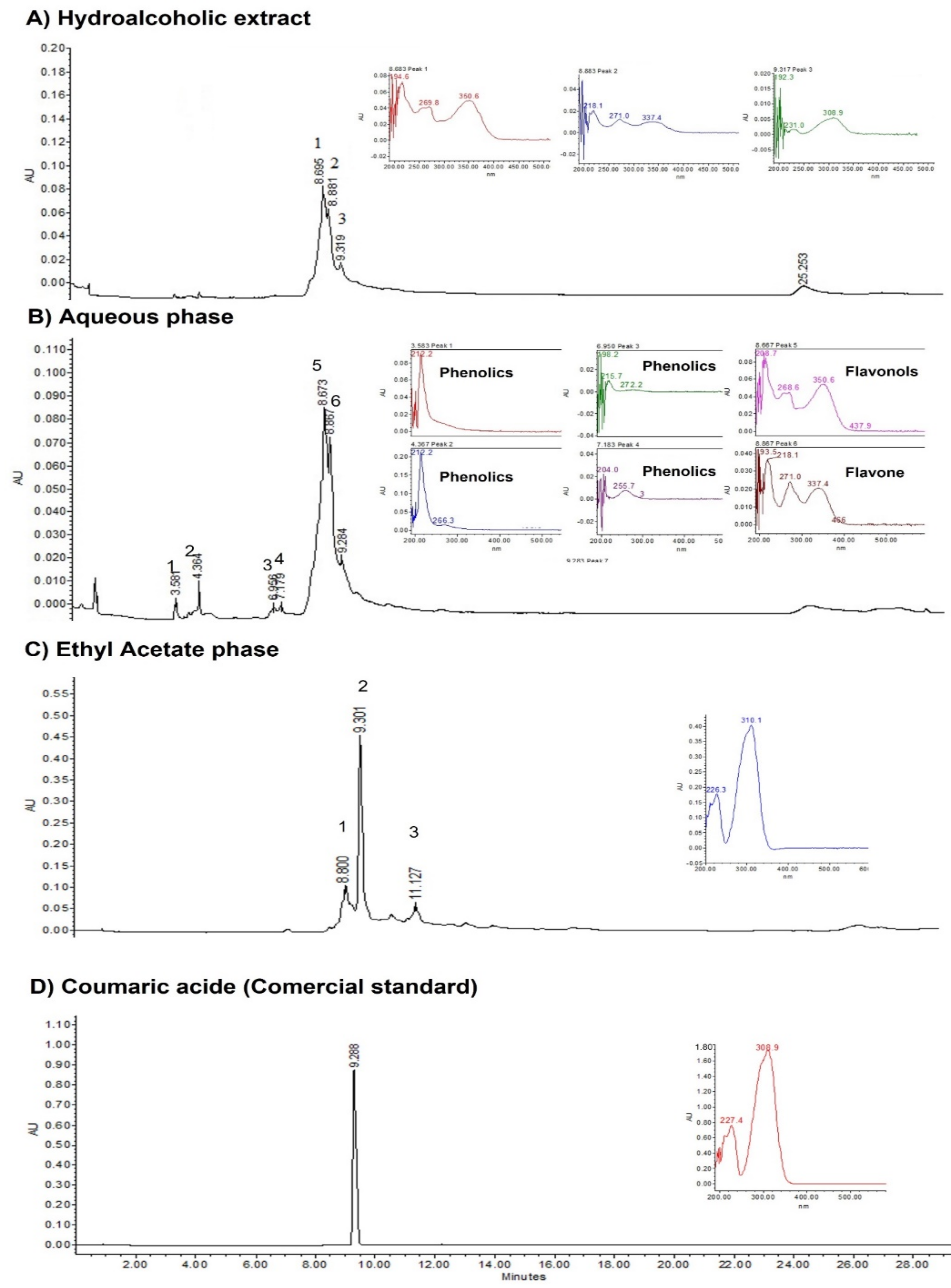
2.3. HPLC Results
2.4. Microscopic Analysis
3. Discussion
3.1. Ovicidal and Larvicidal Activity
3.2. Analysis of Phytoconstituents
3.3. Microscopic Findings
4. Materials and Methods
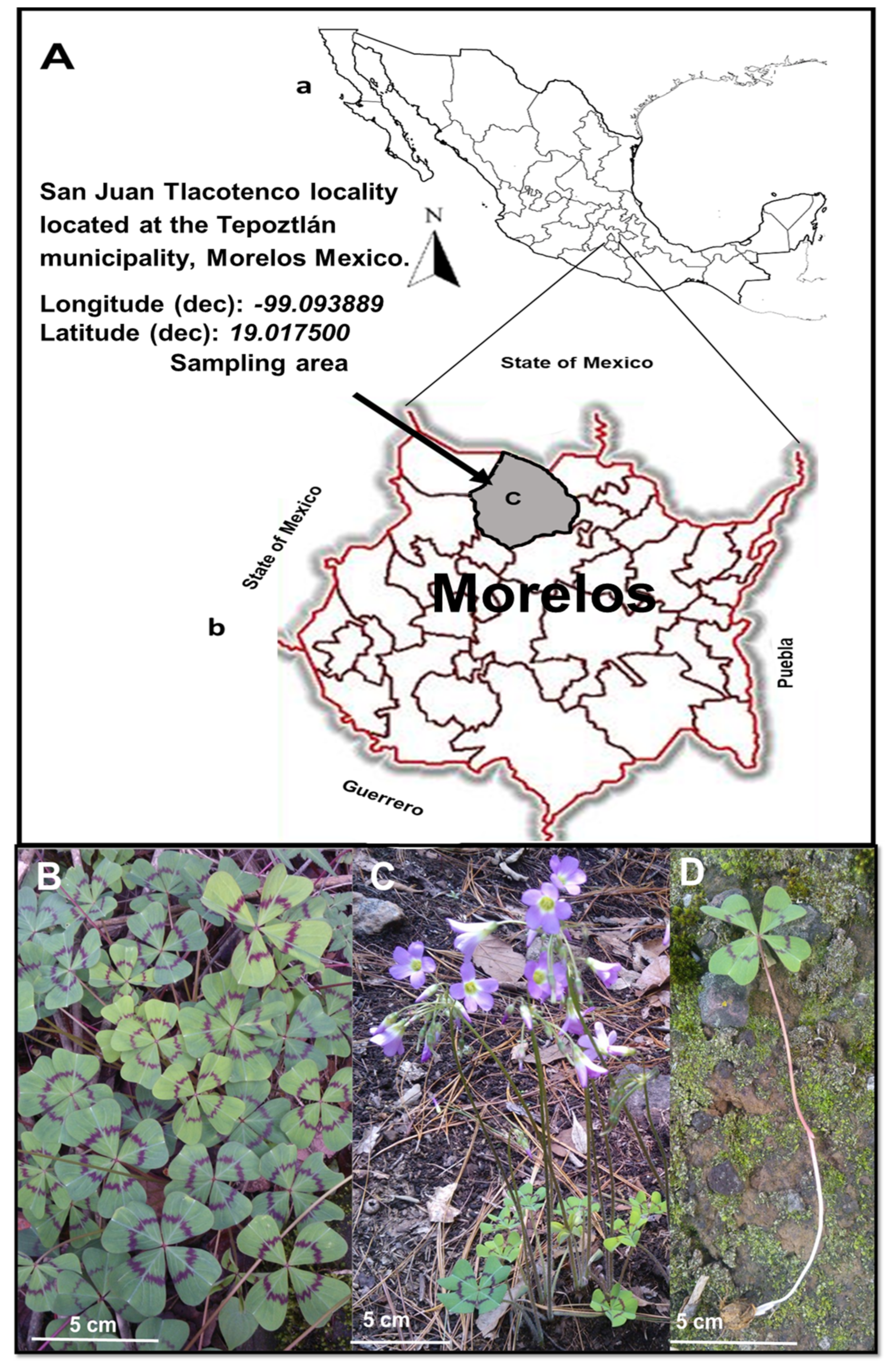
4.1. Location
4.2. Plant Material
4.3. Acquisition of Hydroalcoholic Extracts
4.4. Partition of a Hydroalcoholic Extract
4.5. Chromatographic Fractionation of the Aqueous Phase of Oxalis tetraphylla
4.6. Thin-Layer Chromatography (TLC)
4.7. High-Performance Liquid Chromatography (HPLC) Analysis
4.8. Acquisition of Haemonchus contortus Eggs and Infective Larvae (L3)
4.9. Concentrations Used in Each Hydroalcoholic Extract or Phase of Bipartition from Oxalis tetraphylla Stems and Leaves against Haemonchus contortus
4.10. Experimental Design
4.10.1. Assessing the Ovicidal Activity of Oxalis tetraphylla Extract and Phases against Haemonchus contortus
4.10.2. Assessing the Effect of Oxalis tetraphylla Extract and Phases against Haemonchus contortus Larval Activity
4.11. Statistical Analysis
4.12. Lethal Concentrations (LC50 and LC90)
4.13. Microscopic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halvarsson, P.; Gustafsson, K.; Höglund, J. Farmers’ perception on the control of gastrointestinal parasites in organic and conventional sheep production in Sweden. Vet. Parasitol. Reg. Stud. Rep. 2022, 30, 100713. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.G.; Brophy, P.M.; Williams, H.W.; Davies, N.; Jones, R.A. Gastrointestinal nematode control practices in ewes: Identification of factors associated with application of control methods known to influence anthelmintic resistance development. Vet. Parasitol. Reg. Stud. Rep. 2021, 24, 100562. [Google Scholar] [CrossRef] [PubMed]
- Iliev, T.P.; Prelezov, P.; Ivanov, A.; Kirkova, Z.; Tonev, A. Clinical study of acute haemonchosis in lambs. Trakia J. Sci. 2017, 1, 74–78. [Google Scholar] [CrossRef]
- Flay, K.J.; Hill, F.I.; Muguiro, D.H. A Review: Haemonchus contortus Infection in Pasture-Based Sheep Production Systems, with a Focus on the Pathogenesis of Anaemia and Changes in Haematological Parameters. Animals 2022, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Niciura, S.C.M.; Cruvinel, G.G.; Moraes, C.V.; Chagas, A.C.S.; Esteves, S.N.; Benavides, M.V.; Amarante, A.F.T. In vivo selection for Haemonchus contortus resistance to monepantel. J. Helminthol. 2019, 94, 46. [Google Scholar] [CrossRef]
- Hou, B.; Yong, R.; Wuen, J.; Zhang, Y.; Buyin, B.; Subu, D.; Zha, H.; Li, H.; Hasi, S. Positivity Rate Investigation and Anthelmintic Resistance Analysis of Gastrointestinal Nematodes in Sheep and Cattle in Ordos, China. Animals 2022, 12, 891. [Google Scholar] [CrossRef]
- Han, T.; Wang, M.; Zhang, G.; Xinwei, L.; Guowen, L.; Xiaobing, L.; Zhe, W. Gastrointestinal nematodes infections and anthelmintic resistance in grazing sheep in the Eastern Inner Mongolia in China. Acta Parasitol. 2017, 62, 815–822. [Google Scholar] [CrossRef]
- Mondragón-Ancelmo, J.; Olmedo-Juárez, A.; Reyes-Guerrero, D.E.; Ramírez-Vargas, G.; Ariza-Román, A.E.; López-Arellano, M.E.; Mendoza-de Gives, P.; Napolitano, F. Detection of Gastrointestinal Nematode Populations Resistant to Albendazole and Ivermectin in Sheep. Animals 2019, 9, 775. [Google Scholar] [CrossRef]
- Santiago-Figueroa, I.; Lara-Bueno, A.; González-Garduño, R.; López-Arellano, M.E.; de la Rosa-Arana, J.L.; Maldonado-Simán, E.J. Anthelmintic resistance in hair sheep farms in a sub-humid tropical climate, in the Huasteca Potosina, Mexico. Vet. Parasitol. Reg. Stud. Rep. 2019, 17, 100292. [Google Scholar] [CrossRef]
- Ocegueda, S.; Moreno, E.; Koleff, P. Plantas utilizadas en la medicina tradicional y su identificación científica. Biodiversitas 2005, 62, 12–15. [Google Scholar]
- Sánchez-Pedraja, Ó. Oxalis. In Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares (Rhamnaceae-Polygalaceae); Muñoz, G.F., Navarro, C., Quintanar, A., Buira, A., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 2015; Volume 9, pp. 383–405. [Google Scholar]
- Sarkar, T.; Ghosh, P.; Poddar, S.; Choudhury, S.; Sarkar, A.; Chatterjee, S. Oxalis corniculata Linn. (Oxalidaceae): A brief review. J. Pharmacogn. Phytochem. 2020, 9, 651–655. [Google Scholar]
- Vineer, H.R.; Morgan, E.R.; Hertzberg, H.; Bartley, D.J.; Bosco, A.; Charlier, J.; Chartier, C.; Claerebout, E.; de Waal, T.; Hendrickx, G.; et al. Increasing importance of anthelmintic resistance in European livestock: Creation and meta-analysis of an open database. Parasite 2020, 27, 69. [Google Scholar] [CrossRef] [PubMed]
- Wondimu, A.; Bayu, Y. Anthelmintic Drug Resistance of Gastrointestinal Nematodes of Naturally Infected Goats in Haramaya, Ethiopia. J. Parasitol. Res. 2022, 2022, 4025902. [Google Scholar] [CrossRef] [PubMed]
- González-Cruz, B.J.; Rodríguez-Labastida, M.; González-Cortázar, M.; Zamilpa, A.; López-Arellano, M.E.; Aguilar-Marcelino, L.; González-Garduño, R.; Torres-Acosta, J.F.J.; Olmedo-Juárez, A.; Mendoza-de Gives, P. In vitro larvicidal and in vivo anthelmintic effects of Oxalis tetraphylla (Oxalidaceae) hydroalcoholic extract against Haemonchus contortus in lambs. J. Helminthol. 2018, 92, 309–316. [Google Scholar] [CrossRef]
- von Son-de Fernex, E.; Alonso, D.M.A.; Mendoza de Gives, P.; Valles de la Mora, B.; Zamilpa, A.; Gonzalez, C.M. Actividad ovicida de cuatro especies de plantas contra el nematodo gastrointestinal Cooperia punctata. Vet. Mex. 2016, 3, 10–25. [Google Scholar]
- von Son-de Fernex, E.; Alonso-Díaz, M.A.; Mendoza-de Gives, P.; Valles-de la Mora, B.; González-Cortazar, M.; Zamilpa, A.; Castillo, G.E. Elucidation of Leucaena leucocephala anthelmintic-like phytochemicals and the ultrastructural damage generated to eggs of Cooperia spp. Vet. Parasitol. 2015, 30, 89–95. [Google Scholar] [CrossRef]
- Muhammad, A.; Ahmed, H.; Iqbal, M.N.; Qayyum, M. Detection of multiple anthelmintic resistance of Haemonchus contortus and Teladorsagia circumcincta in sheep and goats of Northern Punjab, Pakistan. Kafkas Univ. Vet. Fak. Derg. 2015, 21, 389–395. [Google Scholar]
- Mancilla-Montelongo, G.; Castañeda-Ramírez, G.S.; Juan Felipe de Jesús Torres-Acosta, J.F.J.; Sandoval-Castro, C.A.; Borges-Argáez, R. Evaluation of cinnamic acid and six analogues against eggs and larvae of Haemonchus contortus. Vet. Parasitol. 2019, 270, 25–30. [Google Scholar] [CrossRef]
- Castillo-Mitre, G.F.; Olmedo-Juárez, A.; Rojo-Rubio, R.; González-Cortázar, M.; Mendoza-de Gives, P.; Hernández-Beteta, E.E.; Reyes-Guerrero, D.E.; López-Arellano, M.E.; Vázquez-Armijo, J.F.; Ramírez-Vargas, G.; et al. Caffeoyl and coumaroyl derivatives from Acacia cochliacantha exhibit ovicidal activity against Haemonchus contortus. J. Ethnopharmacol. 2017, 23, 125–131. [Google Scholar] [CrossRef]
- Rastija, V.; Vrandečić, K.; Ćosić, J.; Šarić, G.K.; Majić, I.; Agić, D.; Šubarić, D.; Karnaš, M.; Bešlo, D.; Komar, M.; et al. Effects of Coumarinyl Schiff Bases against Phytopathogenic Fungi, the Soil-Beneficial Bacteria and Entomopathogenic Nematodes: Deeper Insight into the Mechanism of Action. Molecules 2022, 27, 2196. [Google Scholar] [CrossRef]
- El Mihyaoui, A.; Esteves-da Silva, J.C.G.; Charfi, S.; Candela, C.M.E.; Lamarti, A.; Arnao, M.B. Chamomile (Matricaria chamomilla L.): A Review of Ethnomedicinal Use, Phytochemistry and Pharmacological Uses. Life 2022, 12, 479. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, R.; Kaur, D.J.; Bajaj, K.L. Nematicidal activity of phenolic compounds against Meloigogyne incognita. Nematol. Mediterr. 1992, 20, 217–219. [Google Scholar]
- González-Cortazar, M.; Zamilpa, A.; López-Arellano, M.E.; Aguilar-Marcelino, L.; Reyes-Guerrero, D.E.; Olazarán-Jenkins, S.; Ramírez-Vargas, G.; Olmedo-Juárez, A.; Mendoza-de-Gives, P. Lysiloma acapulcensis leaves contain anthelmintic metabolites that reduce the gastrointestinal nematode egg population in sheep faeces. Comp. Clin. Pathol. 2018, 27, 189–197. [Google Scholar] [CrossRef]
- Spiegler, V.; Sendker, J.; Petereit, F.; Liebau, E.; Hensel, A. Bioassay-Guided Fractionation of a Leaf Extract from Combretum mucronatum with Anthelmintic Activity: Oligomeric Procyanidins as the Active Principle. Molecules 2015, 20, 14810–14832. [Google Scholar] [CrossRef]
- Barrau, E.; Fabre, N.; Fouraste, I.; Hoste, H. Effect of bioactive compounds from sainfoin (Onobrychis viciifolia) on the in vitro larval migration of Haemonchus contortus: Role of tannins and flavonol glycosides. Parasitology 2005, 131, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Akkari, H.; Rtibi, K.; B’chir, F.; Rekik, M.; Darghouth, M.A.; Gharbi, M. In vitro evidence that the pastoral Artemisia campestris species exerts an anthelmintic effect on Haemonchus contortus from sheep. Vet. Res. Commun. 2014, 38, 249–255. [Google Scholar] [CrossRef]
- Patil, S.A.; Prabhakara, C.T.; Halasangi, B.M.; Toragalmath, S.S.; Badami, P.S. DNA cleavage, antibacterial, antifungal and anthelmintic studies of Co(II), Ni(II) and Cu(II) complexes of coumarin Schiff bases: Synthesis and spectral approach. Spectrochim Acta A Mol. Biomol. Spectrosc. 2015, 137, 641–651. [Google Scholar] [CrossRef]
- von Son-de Fernex, E.; Alonso-Díaz, M.A.; Valles-de la Mora, B.; Mendoza-de Gives, P.; González-Cortazar, M.; Zamilpa, A. Anthelmintic effect of 2H-chromen-2-one isolated from Gliricidia sepium against Cooperia punctata. Exp. Parasitol. 2017, 178, 1–6. [Google Scholar] [CrossRef]
- Lacy, A.; O’Kennedy, R. Studies on coumarins and coumarin-related compounds to determine their therapeutic role in the treatment of cancer. Curr. Pharm. Des. 2004, 10, 3797–3811. [Google Scholar] [CrossRef]
- Olmedo-Juárez, A.; Zarza-Albarran, M.A.; Rojo-Rubio, R.; Zamilpa, A.; González-Cortazar, M.; Mondragón-Ancelmo, J.; Rivero-Pérez, J.N.; Mendoza-de Gives, P. Acacia farnesiana pods (plant: Fabaceae) possesses anti-parasitic compounds against Haemonchus contortus in female lambs. Exp. Parasitol. 2020, 218, 107980. [Google Scholar] [CrossRef]
- Delgado-Núñez, E.J.; Zamilpa, A.; González-Cortazar, M.; Olmedo-Juárez, A.; Cardoso-Taketa, A.; Sánchez-Mendoza, E.; Tapia-Maruri, D.; Salinas-Sánchez, D.O.; Mendoza-de Gives, P. Isorhamnetin: A Nematocidal Flavonoid from Prosopis laevigata Leaves Against Haemonchus contortus Eggs and Larvae. Biomolecules 2020, 10, 773. [Google Scholar] [CrossRef] [PubMed]
- Brunet, S.; Fourquaux, I.; Hoste, H. Ultrastructural changes in the third-stage, infective larvae of ruminant nematodes treated with sainfoin (Onobrychis vicifolia) extract. Parasitol. Int. 2011, 60, 419–424. [Google Scholar] [CrossRef] [PubMed]
- López, A.U.; Salinas-Sánchez, O.D.; Mendoza, G.P.; López, A.M.E.; Liébano, H.E.; Valladares, C.G.; Arias, D.M.; Hernández, V.V.M. In vitro nematicidal effect of medicinal plants from “Sierra de Huautla”, Biosphere Reserve, Morelos, Mexico against Haemonchus contortus infective larvae. J. Helminthol. 2008, 82, 25–31. [Google Scholar] [CrossRef]
- Olmedo-Juarez, A.; Rojo-Rubio, R.; Zamilpa, A.; Mendoza de Gives, P.; Arece-García, J.; López-Arellano, M.E.; von Son-de Fernex, E. In vitro larvicidal effect of a hydroalcoholic extract from Acacia cochliacantha leaf against 6 ruminant parasitic nematodes. Vet. Res. Commun. 2017, 41, 227–232. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (mg/mL) | Mean of Unhatched Eggs/ Mean of Recovered Larvae (*) | EHI % ± SD |
|---|---|---|
| 0.065 | 22/242 | 9 ± 2.7 e |
| 0.1 | 54/197 | 27.4 ± 1.0 de |
| 0.15 | 96/238 | 40.3 ±12.8 cd |
| 0.2 | 155/266 | 58.2 ±10.3 bc |
| 0.3 | 166/241 | 68.9 ± 8.58 b |
| Ivermectin (5 mg/mL) | 237/237 | 100 a |
| Distilled water | 18/250 | 7.2 ± 2.7 e |
| Concentration (mg/mL) | Mean of Dead Larvae/Mean of Recovered Larvae (*) | Larval Mortality % ± SD |
|---|---|---|
| 15 | 154/417 | 37 ± 3.7 d |
| 20 | 196/389 | 50.3 ± 1.9 c |
| 30 | 170/308 | 55.2 ± 2 c |
| 50 | 335/405 | 83 ± 2.8 b |
| 60 | 380/434 | 88 ± 3.2 b |
| Ivermectin (5 mg/mL) | 380/380 | 100 a |
| Distilled water | 14/322 | 4.5 ± 2.75 e |
| Parasitic Stage | Lethal Concentrations and Confidence Interval (Lower–Upper) | |
|---|---|---|
| LC50 (mg/mL) | LC90 (mg/mL) | |
| Eggs | 0.21 (0.18–0.24) | 0.71 (0.62–0.97) |
| Larvae | 28 (25.9–29.7) | 69.3 (63–78.7) |
| Treatment | Mean of Unhatched Eggs/Mean of Recovered Larvae (*) | Percentages of Egg-Hatching Inhibition | Mean of Unhatched Eggs/Mean of Recovered Larvae * | Percentages of Egg-Hatching Inhibition ± SD |
|---|---|---|---|---|
| 0.25 mg/mL | 1 mg/mL | |||
| Aqueous phase (Aq-Ph) | 391/14.3 | 92.5 ± 7 a | 450/2.3 | 96.6 ± 0.5 a |
| Ethyl Acetate Phase (EtOAc-Ph) | 519/0 | 93 a | 603/0 | 93.6 a |
| Ivermectin (5 mg/mL) | 17/506.6 | 3.4 ± 3.08 b | 442/64.3 | 87.2 ± 12.8 b |
| DMSO 0.5% | 30/488 | 6.3 ± 4.41 b | 30/488 | 6.3 ± 4.41 c |
| Control water | 8/392 | 2.8 ± 3.44 b | 8/392 | 2.8 ± 3.44 c |
| Treatment | Mean of Dead Larvae/Mean of Live Larvae | % Larval Mortality ±SD | Mean of Dead Larvae/Mean of Live Larvae | % Larval Mortality ±SD | Mean of Dead Larvae/Mean of Live Larvae | % Larval Mortality ±SD |
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||||
| Aqueous Phase (Aq-Ph) | 618/152 | 82 ± 9.1 b | 725/127 | 82 ± 9 b | 747/144 | 81.6 ± 14 b |
| Ethyl Acetate Phase (EtOAc-Ph) | 9/656 | 1.2 ± 0.5 c | 11/690 | 1.5 ± 0.1 c | 8/648 | 1.7 ± 1.1 c |
| Ivermectin (5 mg/mL) | 211/0 | 100 a | 308/0 | 100 a | 630/0 | 100 a |
| DMSO 1% | 0/728 | 0 c | 0/760 | 0 c | 760/12 | 0 c |
| Distilled water | 13/540 | 3.7 ± 2 c | 3/594 | 0.44 ± 0.4 c | 0/620 | 0 c |
| Treatment | Mean of Dead Larvae/Mean of Live Larvae | % Larval Mortality ±SD | Mean of Dead Larvae/Mean of Live Larvae | % Larval Mortality ±SD | Mean of Dead Larvae/Mean of Live Larvae | % Larval Mortality ±SD |
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||||
| Fractions 10 mg/mL: | ||||||
| F1 | 25/283 | 8.1 ± 5.6 b | 41/264 | 13.4 ± 7 c | 39/245 | 13.7 ± 2.3 c |
| F2 | 5/280 | 1.7 ± 0.7 c | 26/261 | 9 ± 3.7 cd | 10/293 | 3.3 ± 2.7 de |
| F3 | 11/263 | 4 ± 2 bc | 89/202 | 30.6 ± 3.4 b | 142/134 | 51.4 ± 5 b |
| F4 | 5/302 | 1.6 ± 1.2 c | 13/336 | 3.7 ± 2.4 c | 31/292 | 9.6 ± 3.2 cd |
| F5 | 7/302 | 2.2 ± 0.6 bc | 9/336 | 2.6 ± 2.2 c | 10/306 | 3.1 ± 2.3 e |
| Ivermectin (5 mg/mL) | 338/0 | 100 a | 336/0 | 100 a | 372/0 | 100 a |
| Water | 0/286 | 0 c | 1/325 | 0.3 ± 0.6 d | 4/297 | 1.3 ± 1.1 e |
| Fractions 30 mg/mL: | ||||||
| F1 | 135/134 | 50.2 ± 2.5 c | 206/46 | 81.7 ± 12.6 b | 242/44 | 84.6 ± 3.1 b |
| F2 | 21/188 | 10 ± 2.4 e | 105/108 | 49.3 ± 14.4 c | 192/68 | 73.8 ± 5.6 bc |
| F3 | 285/45 | 86.3 ± 4.2 b | 328/21 | 94 ± 2.5 b | 280/34 | 89.2 ± 2.7 b |
| F4 | 92/272 | 25.3 ± 3.3 d | 184/174 | 51.4 ± 5.6 c | 194/142 | 57.7 ± 7.2 c |
| F5 | 4/335 | 1.1 ± 0.9 f | 63/274 | 18.7 ± 6 d | 117/295 | 28.4 ± 9.9 d |
| Ivermectin (5 mg/mL) | 338/0 | 100 a | 336/0 | 100 a | 372/0 | 100 a |
| Distilled Water | 0/296 | 0 g | 1/325 | 0.3 ± 0.6 e | 4/352 | 1.1 ± 0.9 e |
| Plant | Compound | Parasite | Concentration | Effect | Authors |
|---|---|---|---|---|---|
| Onobrychis viciifolia | Condensed tannin <2000 Da, 3 flavonol glycosides as: rutin, nicotiflorin and narcissin | Haemonchus contortus (L3) | 1200 µg/mL each compound | Inhibition of larval migration | Barrau et al., 2005 [26] |
| Artemisia campestris | Ethanolic extract: Quercetin, 3 methyl-quercetin, 7 methyl-taxifolin 7,3 methyl-kaempferol. Aqueous extract: Apigenin-6,8-di C-glucoside Eupafolin-glucoside Acethyl-luteolin-glucuronide Apigenin-7-glucoronide Hesperidin Luteolin | H. contortus (L3) | Ethanolic extract: 0.5 1 and 2 mg/mL Aqueous extract. 0.5 1 and 2 mg/mL | 70–74% Mortality at 24 h. 100% mortality at 24 h | Akkari et al., 2014 [27] |
| Acacia cochliacantha | Caffeoyl and coumaroyl derivatives: caffeic acid, p-coumaric acid, ferulic acid, methyl caffeate, methyl-p-coumarate, methyl ferulate and quercetin | H. contortus eggs | 1 mg/mL | Ovicidal activity 71–98% | Castillo-Mitre et al., 2017 [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ocampo-Gutiérrez, A.Y.; Hernández-Velázquez, V.M.; Zamilpa, A.; López-Arellano, M.E.; Olmedo-Juárez, A.; Higuera-Piedrahita, R.I.; Delgado-Núñez, E.J.; González-Cortázar, M.; Mendoza-de Gives, P. Oxalis tetraphylla (Class: Magnoliopsidae) Possess Flavonoid Phytoconstituents with Nematocidal Activity against Haemonchus contortus. Pathogens 2022, 11, 1024. https://doi.org/10.3390/pathogens11091024
Ocampo-Gutiérrez AY, Hernández-Velázquez VM, Zamilpa A, López-Arellano ME, Olmedo-Juárez A, Higuera-Piedrahita RI, Delgado-Núñez EJ, González-Cortázar M, Mendoza-de Gives P. Oxalis tetraphylla (Class: Magnoliopsidae) Possess Flavonoid Phytoconstituents with Nematocidal Activity against Haemonchus contortus. Pathogens. 2022; 11(9):1024. https://doi.org/10.3390/pathogens11091024
Chicago/Turabian StyleOcampo-Gutiérrez, Ana Yuridia, Víctor Manuel Hernández-Velázquez, Alejandro Zamilpa, María Eugenia López-Arellano, Agustín Olmedo-Juárez, Rosa Isabel Higuera-Piedrahita, Edgar Jesús Delgado-Núñez, Manasés González-Cortázar, and Pedro Mendoza-de Gives. 2022. "Oxalis tetraphylla (Class: Magnoliopsidae) Possess Flavonoid Phytoconstituents with Nematocidal Activity against Haemonchus contortus" Pathogens 11, no. 9: 1024. https://doi.org/10.3390/pathogens11091024