
Dirofilaria immitis: Genotyping Randomly Selected European Clinical Samples and USA Laboratory Isolates with Molecular Markers Associated with Macrocyclic Lactone Susceptibility and Resistance

 ,
,
Abstract
:1. Introduction
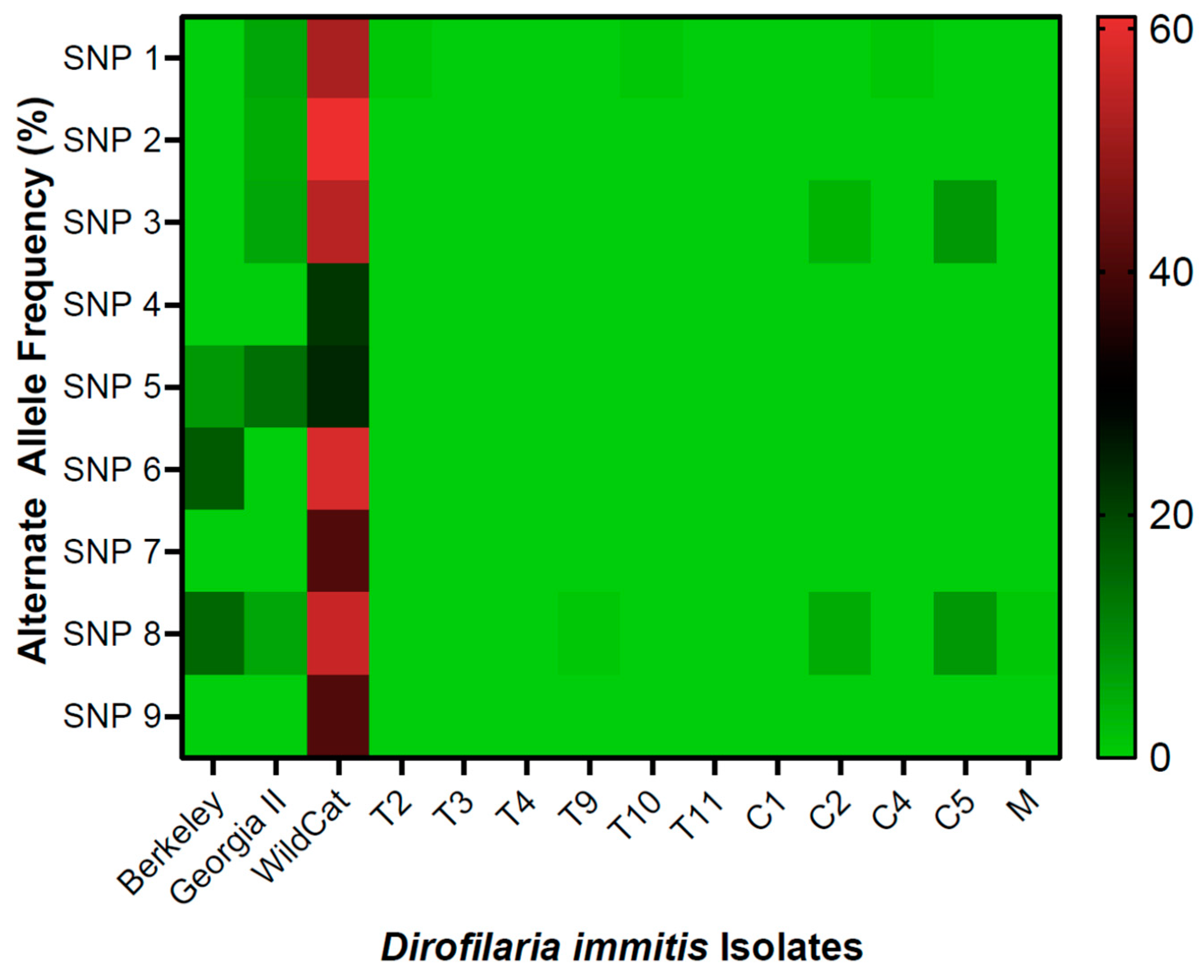
2. Results
3. Discussion
4. Materials and Methods
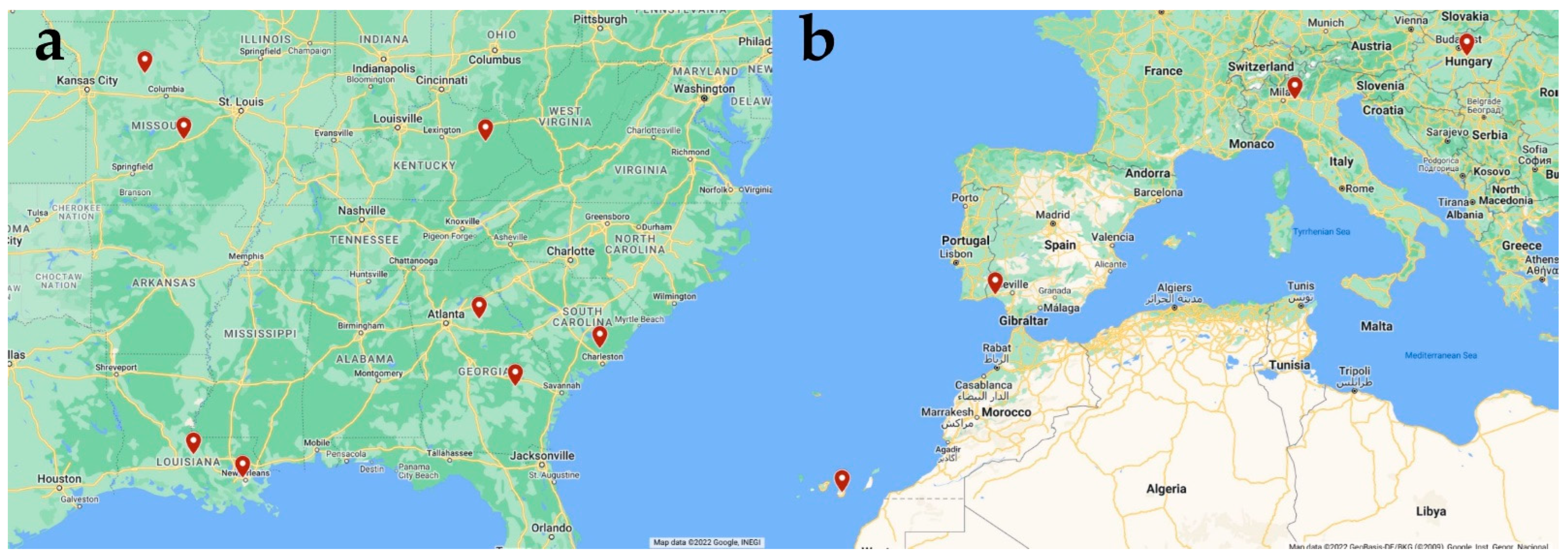
4.1. USA Samples
4.2. European Sample Details
4.3. Sample Processing and DNA Extraction
4.4. SNP Markers
4.5. Sequencing
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chabala, J.C.; Mrozik, H.; Tolman, R.L.; Eskola, P.; Lusi, A.; Peterson, L.H.; Woods, M.F.; Fisher, M.H.; Campbell, W.C. Ivermectin, a new broad-spectrum antiparasitic agent. J. Med. Chem. 1980, 23, 1134–1136. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.C. An introduction to the avermectins. N. Z. Vet. J. 1981, 29, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.C.; Fisher, M.H.; Stapley, E.O.; Albers-Schönberg, G.; Jacob, T.A. Ivermectin: A potent new antiparasitic agent. Science 1983, 221, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, I.; Katae, H.; Hayasaki, M.; Nakagaki, K.; Tada, Y. Prophylactic Activity of Ivermectin against Dirofilaria immitis Infection in Dogs: Establishment of Effective Dose and Administration Schedule. Jpn. J. Vet. 1987, 49, 439–445. [Google Scholar] [CrossRef]
- Ohishi, I.; Katae, H.; Hayasaki, M.; Tada, Y. Prophylactic Activity of Ivermectin against Dirofilaria immitis Infection in Dogs: Larvicidal Activity of Ivermectin against D. immitis Larvae 30 Days after Infection. Jpn. J. Vet. 1987, 49, 115–120. [Google Scholar] [CrossRef]
- Hampshire, V.A. Evaluation of efficacy of heartworm preventive products at the FDA. Vet. Parasitol. 2005, 133, 191–195. [Google Scholar] [CrossRef]
- Pulaski, C.N.; Malone, J.B.; Bourguinat, C.; Prichard, R.; Geary, T.; Ward, D.; Klei, T.R.; Guidry, T.; Smith, G.B.; Delcambre, B.; et al. Establishment of macrocyclic lactone resistant Dirofilaria immitis isolates in experimentally infected laboratory dogs. Parasites Vectors 2014, 7, 494. [Google Scholar] [CrossRef]
- Bourguinat, C.; Lee, A.C.Y.; Lizundia, R.; Blagburn, B.L.; Liotta, J.L.; Kraus, M.S.; Keller, K.; Epe, C.; Letourneau, L.; Kleinman, C.L.; et al. Macrocyclic lactone resistance in Dirofilaria immitis: Failure of heartworm preventives and investigation of genetic markers for resistance. Vet. Parasitol. 2015, 210, 167–178. [Google Scholar] [CrossRef]
- Bourguinat, C.; Keller, K.; Xia, J.; Lepage, P.; McTier, T.L.; Woods, D.J.; Prichard, R.K. Genetic profiles of ten Dirofilaria immitis isolates susceptible or resistant to macrocyclic lactone heartworm preventives. Parasites Vectors 2017, 10, 504. [Google Scholar] [CrossRef]
- Bourguinat, C.; Lefebvre, F.; Sandoval, J.; Bondesen, B.; Moreno, Y.; Prichard, R.K. Dirofilaria immitis JYD-34 isolate: Whole genome analysis. Parasites Vectors 2017, 10, 494. [Google Scholar] [CrossRef]
- Ballesteros, C.; Pulaski, C.N.; Bourguinat, C.; Keller, K.; Prichard, R.K.; Geary, T.G. Clinical validation of molecular markers of macrocyclic lactone resistance in Dirofilaria immitis. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Genchi, C.; Kramer, L.H. The prevalence of Dirofilaria immitis and D. repens in the Old World. Vet. Parasitol. 2020, 280, 108995. [Google Scholar] [CrossRef] [PubMed]
- Genchi, C.; Rinaldi, L.; Cascone, C.; Mortarino, M.; Cringoli, G. Is heartworm disease really spreading in Europe? Vet. Parasitol. 2005, 133, 137–148. [Google Scholar] [CrossRef]
- Genchi, C.; Rinaldi, L.; Mortarino, M.; Genchi, M.; Cringoli, G. Climate and Dirofilaria infection in Europe. Vet. Parasitol. 2009, 163, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Traversa, D.; Di Cesare, A.; Conboy, G. Canine and feline cardiopulmonary parasitic nematodes in Europe: Emerging and underestimated. Parasites Vectors 2010, 3, 62. [Google Scholar] [CrossRef] [PubMed]
- Norman, C.; Stavisky, J.; Westgarth, C. Importing rescue dogs into the UK: Reasons, methods and welfare considerations. Vet. Rec. 2020, 186, 248. [Google Scholar] [CrossRef]
- Jacobson, L.S.; Ward, K.A.; Lacaden, A.B.; Hornak, T.A. Prevalence of heartworm in relocated, local and outreach clinic dogs: A Canadian sheltering perspective. Vet. Parasitol. 2020, 283, 109081. [Google Scholar] [CrossRef]
- Sonnberger, K.; Duscher, G.G.; Fuehrer, H.-P.; Leschnik, M. Current trends in canine dirofilariosis in Austria—Do we face a pre-endemic status? Parasitol. Res. 2020, 119, 1001–1009. [Google Scholar] [CrossRef]
- Montoya-Alonso, J.A.; Carretón, E.; Morchón, R.; Silveira-Viera, L.; Falcón, Y.; Simón, F. The impact of the climate on the epidemiology of Dirofilaria immitis in the pet population of the Canary Islands. Vet. Parasitol. 2016, 216, 66–71. [Google Scholar] [CrossRef]
- Bacsadi, Á.; Papp, A.; Szeredi, L.; Tóth, G.; Nemes, C.; Imre, V.; Tolnai, Z.; Széll, Z.; Sréter, T. Retrospective study on the distribution of Dirofilaria immitis in dogs in Hungary. Vet. Parasitol. 2016, 220, 83–86. [Google Scholar] [CrossRef]
- Ionică, A.M.; Matei, I.A.; D’Amico, G.; Ababii, J.; Daskalaki, A.A.; Sándor, A.D.; Enache, D.V.; Gherman, C.M.; Mihalca, A.D. Filarioid infections in wild carnivores: A multispecies survey in Romania. Parasites Vectors 2017, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Farkas, R.; Mag, V.; Gyurkovszky, M.; Takács, N.; Vörös, K.; Solymosi, N. The current situation of canine dirofilariosis in Hungary. Parasitol. Res. 2020, 119, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Savić, S.; Stosic, M.Z.; Marcic, D.; Hernández, I.; Potkonjak, A.; Otasevic, S.; Ruzic, M.; Morchón, R. Seroepidemiological Study of Canine and Human Dirofilariasis in the Endemic Region of Northern Serbia. Front. Vet. Sci. 2020, 7, 571. [Google Scholar] [CrossRef] [PubMed]
- Laidoudi, Y.; Bedjaoui, S.; Latrofa, M.S.; Fanelli, A.; Dantas-Torres, F.; Otranto, D. Genetic and geographical delineation of zoonotic vector-borne helminths of canids. Sci. Rep. 2022, 12, 6699. [Google Scholar] [CrossRef] [PubMed]
- AHS. Current Canine Guidelines for the Prevention, Diagnosis, and Management of Heartworm (Dirofilaria immitis) Infection in Dogs; AHS: Holly Springs, NC, USA, 2014. [Google Scholar]
- Drake, J.; Wiseman, S. Increasing incidence of Dirofilaria immitis in dogs in USA with focus on the southeast region 2013–2016. Parasites Vectors 2018, 11, 39. [Google Scholar] [CrossRef]
- Sanchez, J.; Dharmarajan, G.; George, M.M.; Pulaski, C.; Wolstenholme, A.J.; Gilleard, J.S.; Kaplan, R.M. Using population genetics to examine relationships of Dirofilaria immitis based on both macrocyclic lactone-resistance status and geography. Vet. Parasitol. 2020, 283, 109125. [Google Scholar] [CrossRef]
- Curry, E.; Prichard, R. Development of rapid in vitro colorimetric enzymatic activity assay to differentiate macrocyclic lactone susceptible and resistant Dirofilaria immitis isolates. Vet. Parasitol 2022, 304, 109696. [Google Scholar] [CrossRef]
- Prichard, R.K. Macrocyclic lactone resistance in Dirofilaria immitis: Risks for prevention of heartworm disease. Int. J. Parasitol. 2021, 51, 1121–1132. [Google Scholar] [CrossRef]
- Montoya-Alonso, J.A.; Carretón, E.; Juste, M.; Mellado, I.; Morchón, R.; Simón, F. Epidemiological survey of canine heartworm disease on the island of Gran Canaria (Canary Islands–Spain) between 2000 and 2008. Vet. Parasitol 2010, 173, 165–168. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample | Life Stage | Isolate | Dog Type | ML Treatment | Origin |
|---|---|---|---|---|---|
| USA Laboratory-Maintained Isolates | |||||
| WildCat | Blood mf | WildCat | Unknown | Non treated | West Liberty, KY, USA |
| Berkeley | Blood mf | Berkeley | Unknown | Non treated | Berkeley County, SC, USA |
| Georgia II | Blood mf | Georgia II | Unknown | Non treated | Vidalia, GA, USA |
| European Clinical Samples | |||||
| T2 | Adult ♀ | Unknown | Unknown | Unknown | Lombardy Region, Italy |
| T3 | Adult ♀ | Unknown | Unknown | Unknown | Lombardy Region, Italy |
| T4 | Adult ♀ | Unknown | Unknown | Unknown | Lombardy Region, Italy |
| T9 | Adult ♀ | Unknown | Unknown | Unknown | Hungary |
| T10 | Adult ♀ | Unknown | Unknown | Unknown | Hungary |
| T11 | Adult ♀ | Unknown | Unknown | Unknown | Hungary |
| C1 | Blood mf | Unknown | Canary Mastiff | Non treated | Canary Island, Spain |
| C2 | Blood mf | Unknown | Canary Mastiff | Non treated | Canary Island, Spain |
| C4 | Blood mf | Unknown | Canary Hound | Non treated | Canary Island, Spain |
| C5 | Blood mf | Unknown | Canary Mastiff | Non treated | Canary Island, Spain |
| M | Blood mf | Unknown | Spanish Greyhound | Ivermectin | † Huelva, Andalusia Spain |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curry, E.; Traversa, D.; Carretón, E.; Kramer, L.; Sager, H.; Young, L.; Prichard, R. Dirofilaria immitis: Genotyping Randomly Selected European Clinical Samples and USA Laboratory Isolates with Molecular Markers Associated with Macrocyclic Lactone Susceptibility and Resistance. Pathogens 2022, 11, 934. https://doi.org/10.3390/pathogens11080934
Curry E, Traversa D, Carretón E, Kramer L, Sager H, Young L, Prichard R. Dirofilaria immitis: Genotyping Randomly Selected European Clinical Samples and USA Laboratory Isolates with Molecular Markers Associated with Macrocyclic Lactone Susceptibility and Resistance. Pathogens. 2022; 11(8):934. https://doi.org/10.3390/pathogens11080934
Chicago/Turabian StyleCurry, Emily, Donato Traversa, Elena Carretón, Laura Kramer, Heinz Sager, Lisa Young, and Roger Prichard. 2022. "Dirofilaria immitis: Genotyping Randomly Selected European Clinical Samples and USA Laboratory Isolates with Molecular Markers Associated with Macrocyclic Lactone Susceptibility and Resistance" Pathogens 11, no. 8: 934. https://doi.org/10.3390/pathogens11080934