
Molecular Confirmation of Taenia crassiceps Cysticercosis in a Captive Ring-Tailed Lemur (Lemur catta) in Poland

 , , , and
, , , and
Abstract
:1. Introduction
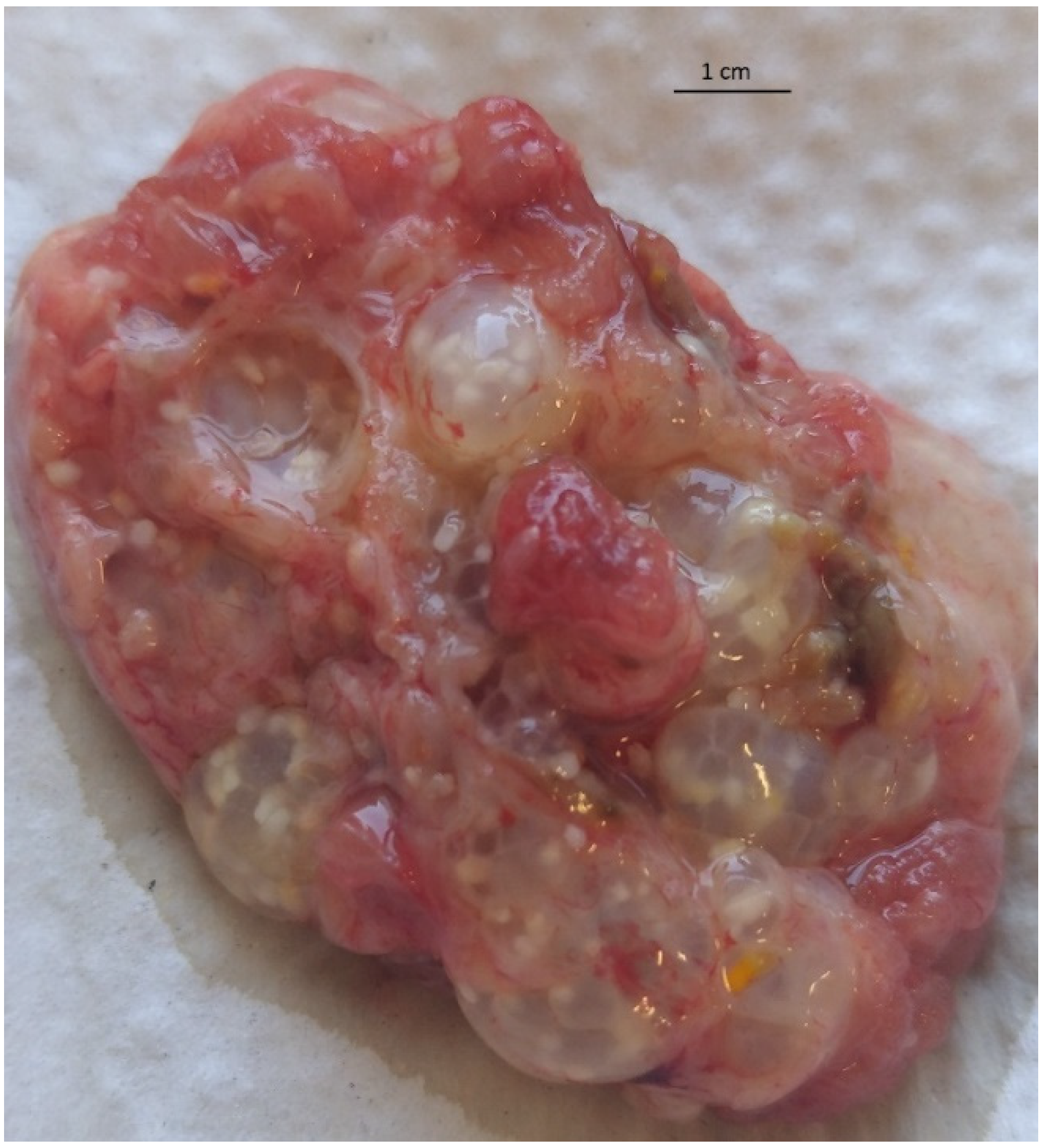
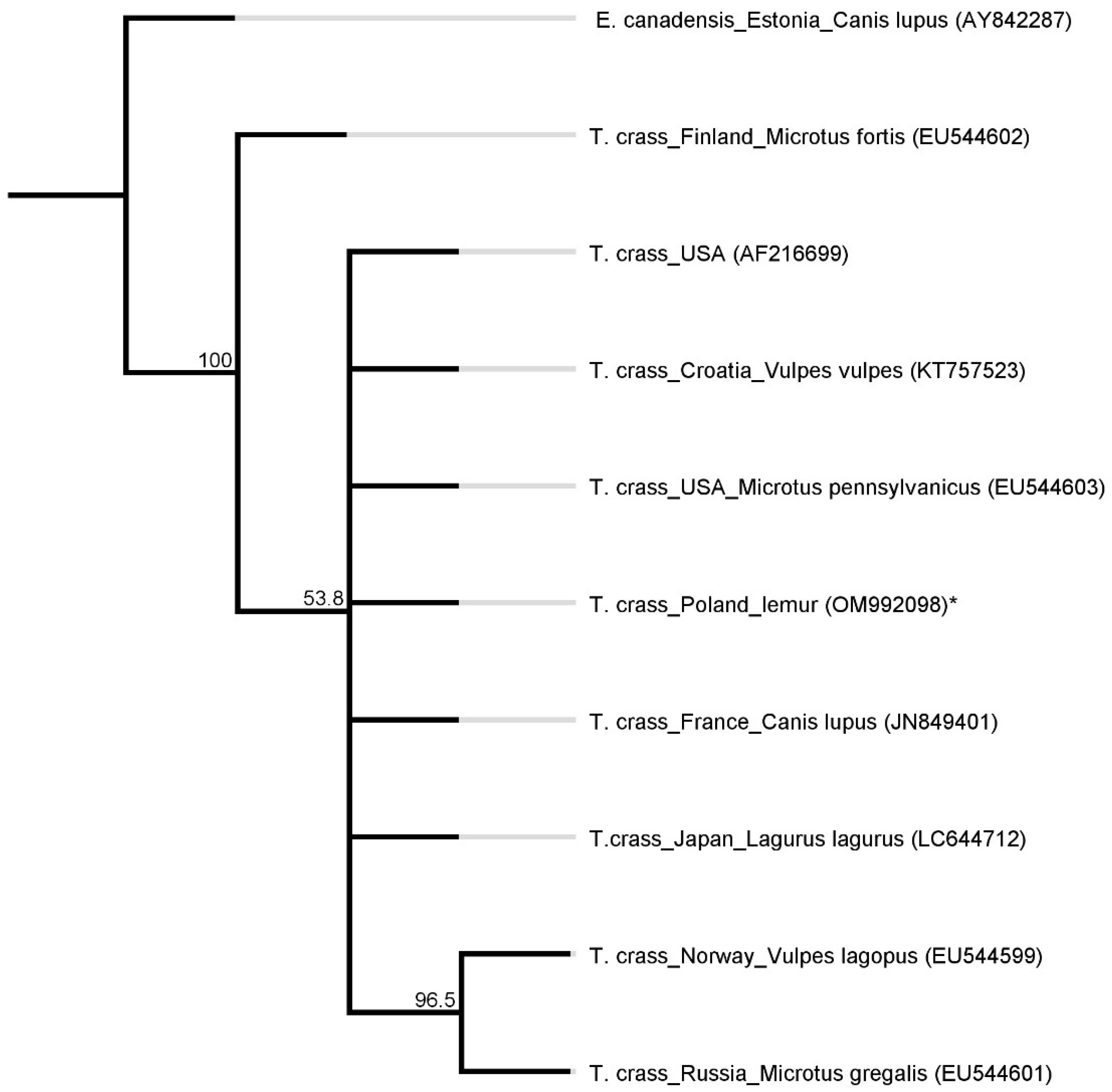
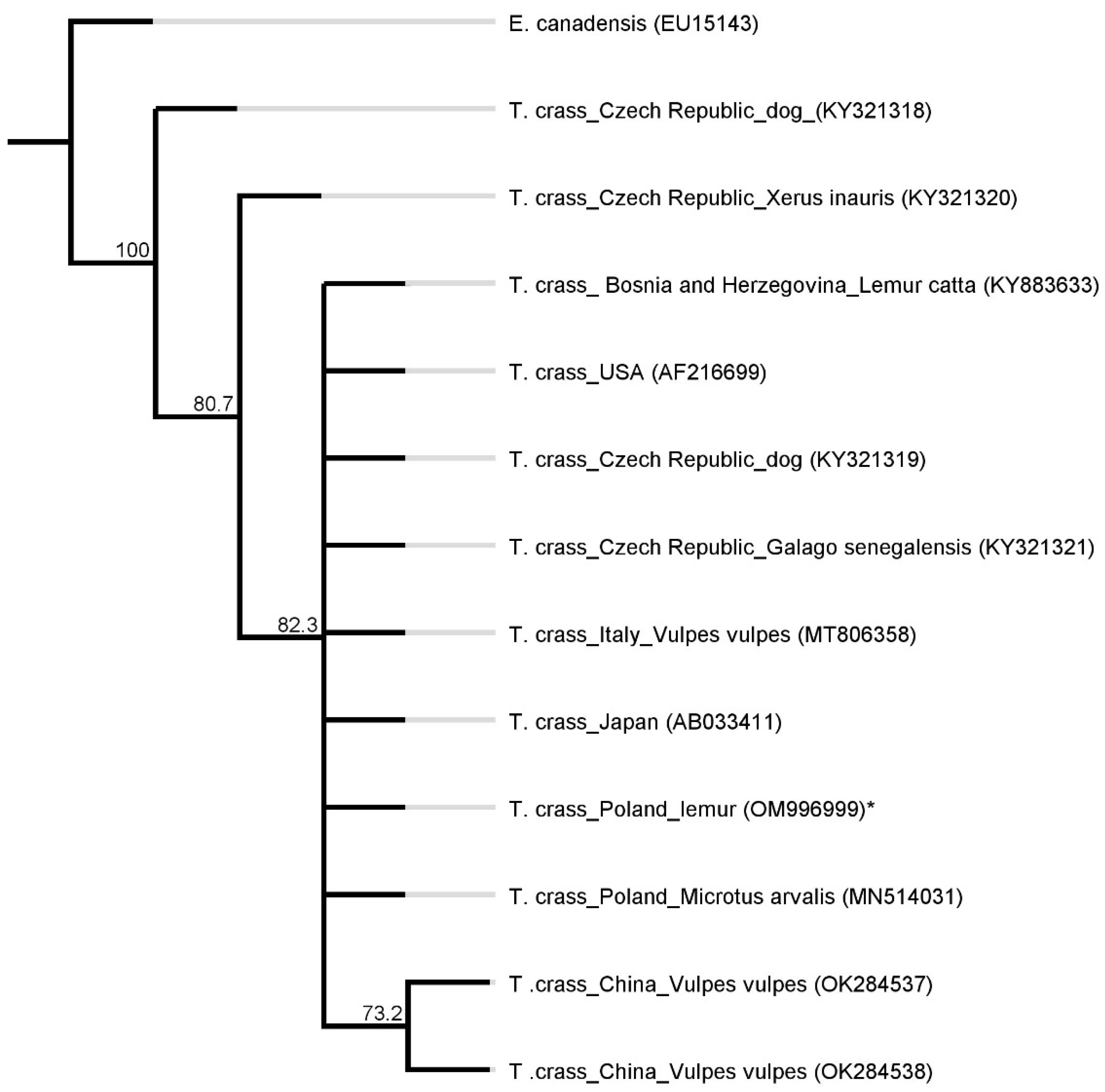
2. Results
3. Discussion
4. Materials and Methods
4.1. Case Description
4.2. Surgery and Sample Collection
4.3. DNA Isolation and PCR Amplification
5. Sequencing and Phylogenetic Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lescano, A.G.; Zunt, J. Other cestodes: Sparganosis, coenurosis and Taenia crassiceps cysticercosis. In Handbook of Clinical Neurolology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 114, pp. 335–345. [Google Scholar] [CrossRef] [Green Version]
- Loos-Frank, B. An up-date of Verster’s (1969) ‘Taxonomic revision of the genus Taenia Linnaeus’ (Cestoda) in table format. Syst. Parasitol. 2000, 45, 155–183. [Google Scholar] [CrossRef] [PubMed]
- Basso, W.; Rutten, M.; Deplazes, P.; Grimm, F. Generalized Taenia crassiceps cysticercosis in a chinchilla (Chinchilla lanigera). Vet. Parasitol. 2014, 199, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Konjevic, D.; Zivicnjak, T.; Kurilj, A.G.; Sindicic, M.; Martinkovic, F.; Jan, D.S. When things go wrong: Cysticercus longicollis in an adult wild red fox (Vulpes vulpes). Parasitol. Res. 2016, 115, 1345–1348. [Google Scholar] [CrossRef] [PubMed]
- Deplazes, P.; van Knapen, F.; Schweiger, A.; Overgaauw, P.A. Role of pet dogs and cats in the transmission of helminthic zoonoses in Europe, with a focus on echinococcosis and toxocarosis. Vet. Parasitol. 2011, 182, 41–53. [Google Scholar] [CrossRef]
- Whipp, C.J.; Daoust, P.Y.; Conboy, G.; Gelens, H. Abdominal Cysticercosis in a Red Fox (Vulpes vulpes). J. Wildl. Dis. 2017, 53, 197–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoberg, E.P.; Ebinger, W.; Render, J.A. Fatal cysticercosis by Taenia crassiceps (Cyclophyllidea: Taeniidae) in a presumed immunocompromised canine host. J. Parasitol. 1999, 85, 1174–1178. [Google Scholar] [CrossRef] [Green Version]
- Ballweber, L.R. Taenia crassiceps subcutaneous cysticercosis in an adult dog. Vet. Rec. 2009, 165, 693–694. [Google Scholar] [PubMed]
- Hofmannova, L.; Mikes, L.; Jedlickova, L.; Pokorny, J.; Svobodova, V. Unusual cases of Taenia crassiceps cysticercosis in naturally infected animals in the Czech Republic. Vet. Med. 2018, 63, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Loos-Frank, B.; Zeyhle, E. The intestinal helminths of the red fox and some other carnivores in southwest Germany. Z. Parasitenkd. 1982, 67, 99–113. [Google Scholar] [CrossRef]
- Wessbecher, H.; Dalchow, W.; Stoye, M. The helminth fauna of the red fox (Vulpes vulpes LINNE 1758) in the administrative district of Karlsruhe. 1. Cestodes. Dtsch. Tierarztl. Wochenschr. 1994, 101, 322–326. [Google Scholar] [PubMed]
- Pfeifer, F.; Kuschfeldt, S.; Stoye, M. Helminth fauna of red foxes (Vulpes vulpes LINNE 1758) in southern Saxony--2: Nematodes. Dtsch. Tierarztl. Wochenschr. 1997, 104, 475–477. [Google Scholar] [PubMed]
- Hofer, S.; Gloor, S.; Muller, U.; Mathis, A.; Hegglin, D.; Deplazes, P. High prevalence of Echinococcus multilocularis in urban red foxes (Vulpes vulpes) and voles (Arvicola terrestris) in the city of Zurich, Switzerland. Parasitology 2000, 120 Pt 2, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, M.F.; Iglesias, R.; Garcia, J.; Paniagua, E.; Sanmartin, M.L. Intestinal helminths of the red fox (Vulpes vulpes L.) in Galicia (Northwest Spain). Wiad. Parazytol. 1995, 41, 429–442. [Google Scholar]
- Citterio, C.V.; Obber, F.; Trevisiol, K.; Dellamaria, D.; Celva, R.; Bregoli, M.; Ormelli, S.; Sgubin, S.; Bonato, P.; Da Rold, G.; et al. Echinococcus multilocularis and other cestodes in red foxes (Vulpes vulpes) of northeast Italy, 2012–2018. Parasites Vectors 2021, 14, 29. [Google Scholar] [CrossRef]
- Bagrade, G.; Kirjusina, M.; Vismanis, K.; Ozolins, J. Helminth parasites of the wolf Canis lupus from Latvia. J. Helminthol. 2009, 83, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Bruzinskaite-Schmidhalter, R.; Sarkunas, M.; Malakauskas, A.; Mathis, A.; Torgerson, P.R.; Deplazes, P. Helminths of red foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) in Lithuania. Parasitology 2012, 139, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Umhang, G.; Comte, S.; Raton, V.; Hormaz, V.; Boucher, J.M.; Favier, S.; Combes, B.; Boue, F. Echinococcus multilocularis infections in dogs from urban and peri-urban areas in France. Parasitol. Res. 2014, 113, 2219–2222. [Google Scholar] [CrossRef] [PubMed]
- Karamon, J.; Sroka, J.; Dabrowska, J.; Bilska-Zajac, E.; Zdybel, J.; Kochanowski, M.; Rozycki, M.; Cencek, T. First report of Echinococcus multilocularis in cats in Poland: A monitoring study in cats and dogs from a rural area and animal shelter in a highly endemic region. Parasites Vectors 2019, 12, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoberg, E.P. Taenia tapeworms: Their biology, evolution and socioeconomic significance. Microbes Infect. 2002, 4, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Deplazes, P.; Eichenberger, R.M.; Grimm, F. Wildlife-transmitted Taenia and Versteria cysticercosis and coenurosis in humans and other primates. Int. J. Parasitol. Parasites Wildl. 2019, 9, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Ntoukas, V.; Tappe, D.; Pfutze, D.; Simon, M.; Holzmann, T. Cerebellar cysticercosis caused by larval Taenia crassiceps tapeworm in immunocompetent woman, Germany. Emerg. Infect. Dis. 2013, 19, 2008–2011. [Google Scholar] [CrossRef] [PubMed]
- Bobes, R.J.; Estrada, K.; Rios-Valencia, D.G.; Calderon-Gallegos, A.; de la Torre, P.; Carrero, J.C.; Sanchez-Flores, A.; Laclette, J.P. The Genomes of Two Strains of Taenia crassiceps the Animal Model for the Study of Human Cysticercosis. Front. Cell. Infect. Microbiol. 2022, 12, 876839. [Google Scholar] [CrossRef] [PubMed]
- Heldwein, K.; Biedermann, H.G.; Hamperl, W.D.; Bretzel, G.; Loscher, T.; Laregina, D.; Frosch, M.; Buttner, D.W.; Tappe, D. Subcutaneous Taenia crassiceps infection in a patient with non-Hodgkin’s lymphoma. Am. J. Trop. Med. Hyg. 2006, 75, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Chermette, R.; Bussieras, J.; Marionneau, J.; Boyer, E.; Roubin, C.; Prophette, B.; Maillard, H.; Fabiani, B. Invasive cysticercosis due to Taenia crassiceps in an AIDS patient. Bull. Acad. Natl. Med. 1995, 179, 777–780. [Google Scholar] [PubMed]
- Maillard, H.; Marionneau, J.; Prophette, B.; Boyer, E.; Celerier, P. Taenia crassiceps cysticercosis and AIDS. AIDS 1998, 12, 1551–1552. [Google Scholar] [CrossRef] [PubMed]
- Flammer Anikpeh, Y.; Grimm, F.; Lindenblatt, N.; Zinkernagel, A. It isn’t always caviar. BMJ Case Rep. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goesseringer, N.; Lindenblatt, N.; Mihic-Probst, D.; Grimm, F.; Giovanoli, P. Taenia crassiceps upper limb fasciitis in a patient with untreated acquired immunodeficiency syndrome and chronic hepatitis C infection—The role of surgical debridement. J. Plast. Reconstr. Aesthet. Surg. 2011, 64, e174–e176. [Google Scholar] [CrossRef] [PubMed]
- Chuck, R.S.; Olk, R.J.; Weil, G.J.; Akduman, L.; Benenson, I.L.; Smith, M.E.; Kaplan, H.J. Surgical removal of a subretinal proliferating cysticercus of Taeniaeformis crassiceps. Arch. Ophthalmol. 1997, 115, 562–563. [Google Scholar] [CrossRef]
- Schmid, S.; Grimm, F.; Huber, M.; Beck, B.; Custer, P.; Bode, B. Taenia crassiceps infection—An unusual presentation of a tapeworm diagnosed by FNA cytology and PCR. Cytopathology 2014, 25, 340–341. [Google Scholar] [CrossRef] [PubMed]
- Alić, A.; Hodžić, A.; Škapur, V.; Alić, A.Š.; Prašović, S.; Duscher, G.G. Fatal pulmonary cysticercosis caused by Cysticercus longicollis in a captive ring-tailed lemur (Lemur catta). Vet. Parasitol. 2017, 241, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Bleyer, M.; Risch, T.; Roos, C.; Kaup, F.J.; Matz-Rensing, K. Taenia Crassiceps Cysticercosis in a Nilgiri Langur (Semnopithecus Johnii). J. Zoo Wildl. Med. 2018, 49, 501–504. [Google Scholar] [CrossRef]
- Dyer, N.W.; Greve, J.H. Severe Cysticercus longicollis cysticercosis in a black lemur (Eulemur macaco macaco). J. Vet. Diagn. Investig. 1998, 10, 362–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luzon, M.; de la Fuente-Lopez, C.; Martinez-Nevado, E.; Fernandez-Moran, J.; Ponce-Gordo, F. Taenia crassiceps cysticercosis in a ring-tailed lemur (Lemur catta). J. Zoo Wildl. Med. 2010, 41, 327–330. [Google Scholar] [CrossRef]
- Young, L.A.; Morris, P.J.; Keener, L.; Gerdiner, C.H.; Stalis, I.H.; Bickense, E.; Sutherland-Smith, M.; Janseen, D.L. Subcutaneous Taenia crassiceps Cysticercosis in a Red Ruffed Lemur (Varecia variegata rubra). In Proceedings of the AAZV and IAAAM Joint Conference, New Orleans, LA, USA, 17–21 September 2000. [Google Scholar]
- Chou, S.; Ozawa, M.; Matsubara, K.; Tamukai, K.; Tokiwa, T. Subcutaneous cysticercosis due to Taenia crassiceps (Cestoda: Taeniidae) in an imported steppe lemming in Japan. Parasitol. Int. 2022, 87, 102524. [Google Scholar] [CrossRef]
- Bajer, A.; Alsarraf, M.; Dwuznik, D.; Mierzejewska, E.J.; Kolodziej-Sobocinska, M.; Behnke-Borowczyk, J.; Banasiak, L.; Grzybek, M.; Tolkacz, K.; Kartawik, N.; et al. Rodents as intermediate hosts of cestode parasites of mammalian carnivores and birds of prey in Poland, with the first data on the life-cycle of Mesocestoides melesi. Parasites Vectors 2020, 13, 95. [Google Scholar] [CrossRef]
- Delling, C.; Bottcher, D.; Schiffbauer, V.; Bernhard, A.; Schmaschke, R. First report of pulmonary cysticercosis caused by Taenia crassiceps in a Cape fur seal (Arctocephalus pusillus). Int. J. Parasitol. Parasites Wildl. 2019, 10, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Federer, K.; Armua-Fernandez, M.T.; Gori, F.; Hoby, S.; Wenker, C.; Deplazes, P. Detection of taeniid (Taenia spp., Echinococcus spp.) eggs contaminating vegetables and fruits sold in European markets and the risk for metacestode infections in captive primates. Int. J. Parasitol. Parasites Wildl. 2016, 5, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.Z.; Yan, H.B.; Guo, A.J.; Zhu, X.Q.; Wang, Y.C.; Shi, W.G.; Chen, H.T.; Zhan, F.; Zhang, S.H.; Fu, B.Q.; et al. Complete mitochondrial genomes of Taenia multiceps, T. hydatigena and T. pisiformis: Additional molecular markers for a tapeworm genus of human and animal health significance. BMC Genom. 2010, 11, 447. [Google Scholar] [CrossRef] [Green Version]
- Bowles, J.; McManus, D.P. NADH dehydrogenase 1 gene sequences compared for species and strains of the genus Echinococcus. Int. J. Parasitol. 1993, 23, 969–972. [Google Scholar] [CrossRef]
- Dybicz, M.; Borkowski, P.K.; Padzik, M.; Baltaza, W.; Chomicz, L. Molecular determination of suspected alveolar echinococcosis requiring surgical treatment in human cases from Poland. Ann. Parasitol. 2018, 64, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Samorek-Pierog, M.; Karamon, J.; Brzana, A.; Bilska-Zajac, E.; Zdybel, J.; Cencek, T. Molecular Confirmation of Massive Taenia pisiformis Cysticercosis in One Rabbit in Poland. Pathogens 2021, 10, 1029. [Google Scholar] [CrossRef] [PubMed]
- Bowles, J.; Blair, D.; McManus, D.P. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol. Biochem. 1992, 54, 165–174. [Google Scholar] [CrossRef]
- Casulli, A.; Manfredi, M.T.; La Rosa, G.; Cerbo, A.R.; Genchi, C.; Pozio, E. Echinococcus ortleppi and E. granulosus G1, G2 and G3 genotypes in Italian bovines. Vet. Parasitol. 2008, 155, 168–172. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amplified Gene | Primer Name | Sequence (5′-3′) | Amplicon Size [bp] | References |
|---|---|---|---|---|
| nad1 | JB11 | AGATTCGTAAGGGGCCTAATA | ~500 | [41,42] |
| JB12 | ACCACTAACTAATTCACTTTC | |||
| cox1 | CO1F | TTTTTTGGCCATCCTGAGGTTTAT | ~446 | [44,45] |
| CO1R | TAACGACATAACATAATGAAAATG |
| Amplified Gene | Initial Denaturation | Number of Cycles | Denaturation | Annealing | Elongation | Final Extension Step |
|---|---|---|---|---|---|---|
| Temp./Time [min] | Temp./Time [s] | Temp./Time [min] | ||||
| nad1 | 95 °C/3 min | 35 | 95 °C/60 s | 50 °C/60 s | 72 °C/60 s | 72 °C/5 min |
| cox1 | 94 °C/7 min | 38 | 94 °C/30 s | 55 °C/30 s | 72 °C/30 s | 72 °C/5 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samorek-Pieróg, M.; Karamon, J.; Brzana, A.; Sobieraj, L.; Włodarczyk, M.; Sroka, J.; Bełcik, A.; Korpysa-Dzirba, W.; Cencek, T. Molecular Confirmation of Taenia crassiceps Cysticercosis in a Captive Ring-Tailed Lemur (Lemur catta) in Poland. Pathogens 2022, 11, 835. https://doi.org/10.3390/pathogens11080835
Samorek-Pieróg M, Karamon J, Brzana A, Sobieraj L, Włodarczyk M, Sroka J, Bełcik A, Korpysa-Dzirba W, Cencek T. Molecular Confirmation of Taenia crassiceps Cysticercosis in a Captive Ring-Tailed Lemur (Lemur catta) in Poland. Pathogens. 2022; 11(8):835. https://doi.org/10.3390/pathogens11080835
Chicago/Turabian StyleSamorek-Pieróg, Małgorzata, Jacek Karamon, Adam Brzana, Lesław Sobieraj, Mariusz Włodarczyk, Jacek Sroka, Aneta Bełcik, Weronika Korpysa-Dzirba, and Tomasz Cencek. 2022. "Molecular Confirmation of Taenia crassiceps Cysticercosis in a Captive Ring-Tailed Lemur (Lemur catta) in Poland" Pathogens 11, no. 8: 835. https://doi.org/10.3390/pathogens11080835