
The Impact of RNA Interference in Tick Research



1
SaBio, Instituto de Investigación en Recursos Cinegéticos IREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain
2
The Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA
*
Author to whom correspondence should be addressed.
Pathogens 2022, 11(8), 827; https://doi.org/10.3390/pathogens11080827
Submission received: 9 July 2022
/
Revised: 21 July 2022
/
Accepted: 22 July 2022
/
Published: 23 July 2022
(This article belongs to the Special Issue 10th Anniversary of Pathogens—Classic Papers in Tick Research)
{kind=link}
{kind=link}
Abstract
:Over the past two decades, RNA interference (RNAi) in ticks, in combination with omics technologies, have greatly advanced the discovery of tick gene and molecular function. While mechanisms of RNAi were initially elucidated in plants, fungi, and nematodes, the classic 2002 study by Aljamali et al. was the first to demonstrate RNAi gene silencing in ticks. Subsequently, applications of RNAi have led to the discovery of genes that impact tick function and tick-host-pathogen interactions. RNAi will continue to lead to the discovery of an array of tick genes and molecules suitable for the development of vaccines and/or pharmacologic approaches for tick control and the prevention of pathogen transmission.
1. Introduction
RNA interference (RNAi) is a molecular methodology that, in combination with recent omics technologies, has substantially advanced the understanding of tick gene function [1,2,3]. RNAi is effected by the introduction of double-stranded RNA (dsRNA) homologs to specific messenger RNA (mRNA), resulting in sequence-specific degradation, thereby interfering with gene expression, causing a subsequent loss of gene function.
The RNAi method used today evolved from studies on cellular phenomena identified initially in 1990 by Napoli et al. [4] in petunia plants. In this study, the authors hypothesized that the introduction of a chimeric gene would result in reversible co-suppression of homologous genes in trans. A similar phenomenon was subsequently reported in the fungus Neurospora crassa, which proposed that the introduction of homologous RNA would cause quelling of a target gene [5]. Subsequently, RNAi was first reported in 1995 in the nematode Caenorhabditis elegans [6].
In 1998, a pioneering study by Fire et al. [7] elucidated the mechanisms by which dsRNA impacted the phenomena of co-suppression in plants [4], quelling in fungi [5], and RNAi in nematodes [6]. Further key advances in RNAi research included evidence of parent-to-offspring transmission in C. elegans [8,9], identification of small interfering RNAs (siRNAs) as stable RNAi intermediates [10,11,12,13], siRNA-mediated silencing of endogenous and heterologous genes in mammalian cells [14], and identification of enzymes and other components of the RNA-induced silencing complex (RISC) [15,16,17].
The first report of RNAi in ticks in 2002 by Majd N. Aljamali, John R. Sauer, and Richard C. Essenberg from Oklahoma State University (Stillwater, OK, USA) [18], based on these previous studies, was a fundamentally important advance in the study of ticks. RNAi in ticks led to the elucidation of tick gene function and the molecular interactions between ticks and pathogens required for pathogen development and transmission. These studies allowed for the identification of tick genes for use in the development of novel interventions for the control of tick infestations and tick-borne diseases [19].
2. Discovery/Methodology
The classic proof-of-concept study of dsRNA-mediated RNAi in ticks by Aljamali et al. [18] was done using Amblyomma americanum female ticks. In this study, dsRNA targeted to the histamine binding protein (HBP) by RNAi was incubated in vitro with extracted tick salivary glands or injected into female ticks. The incubation and injection of this specific dsRNA caused a reduction in HBP mRNA levels impacting tick feeding, most likely due to higher histamine concentrations at the feeding site [18].
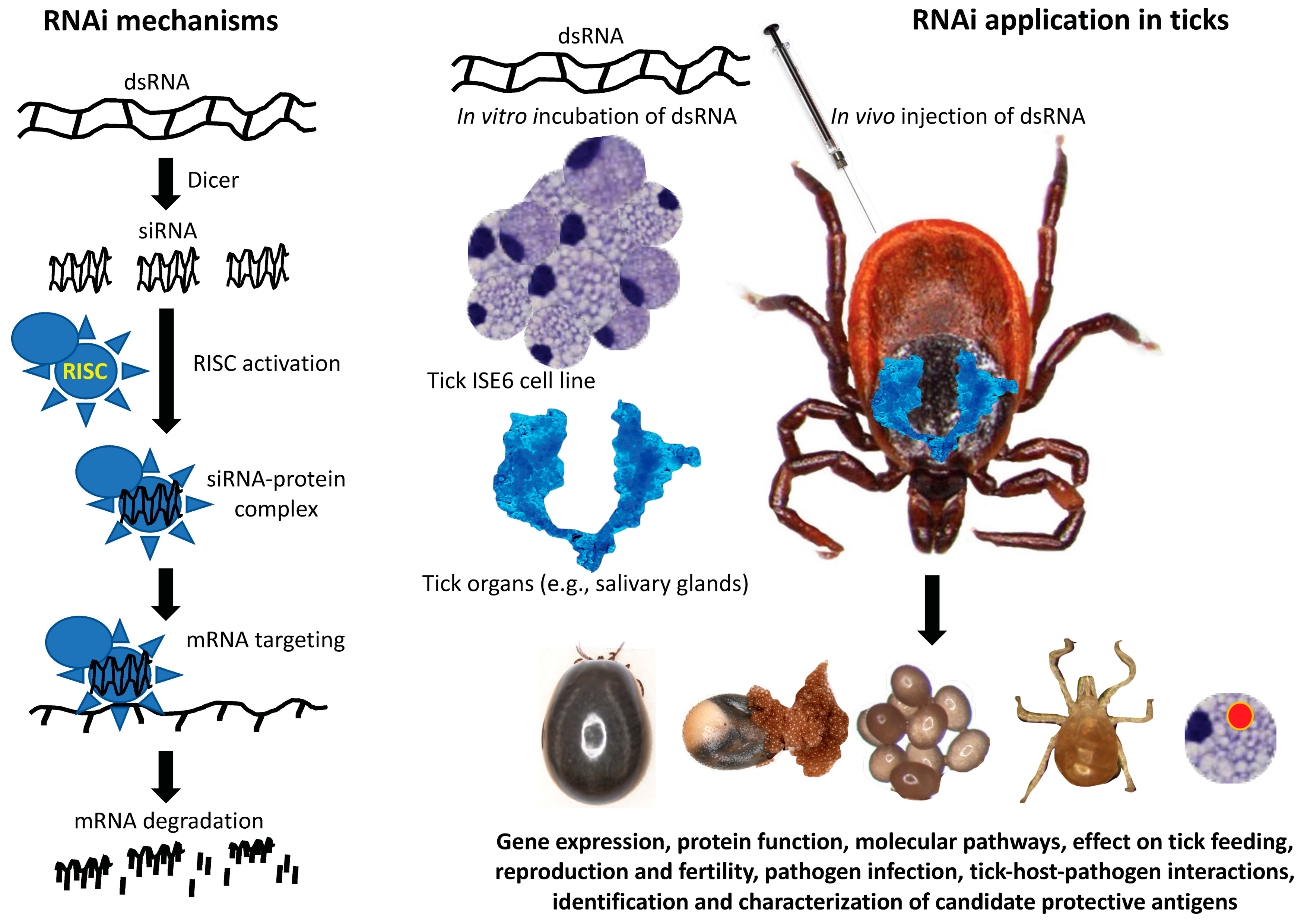
These and other methodologies subsequently developed for RNAi in ticks confirmed previous reports using other species, provided methods for the study of tick gene function, and also led to development of applications for studies on other arthropod ectoparasites (Figure 1).
RNAi in ticks is induced with endogenously present or exogenously introduced dsRNA cleaved to produce siRNAs (21–25 bp) by the ATP-dependent RNase III-like enzyme Dicer. The siRNAs then recruit and activate RISC resulting in unwinds of the siRNA in the siRNA-protein complex. Each strand of the siRNA binds to complementary sequences with activated RISC binding to the targeted RNA, cleaving it and resulting in mRNA degradation. For RNAi in ticks, dsRNA can either be injected in vivo into live ticks or incubated with tick tissues (e.g., salivary glands) or cultured tick cells [20]. The assessment of the effect of RNAi can subsequently be done by analysis of gene mRNA expression and by the impact on the target gene protein function. Additionally, the effect of RNAi on molecular pathways, tick feeding, reproduction and fertility, pathogen infection, and tick-host-pathogen interactions can be evaluated. Through this methodology, candidate protective antigens can be identified, studied, and used for the discovery of novel antigens for vaccine development or for use in other tick interventions.
3. Impact
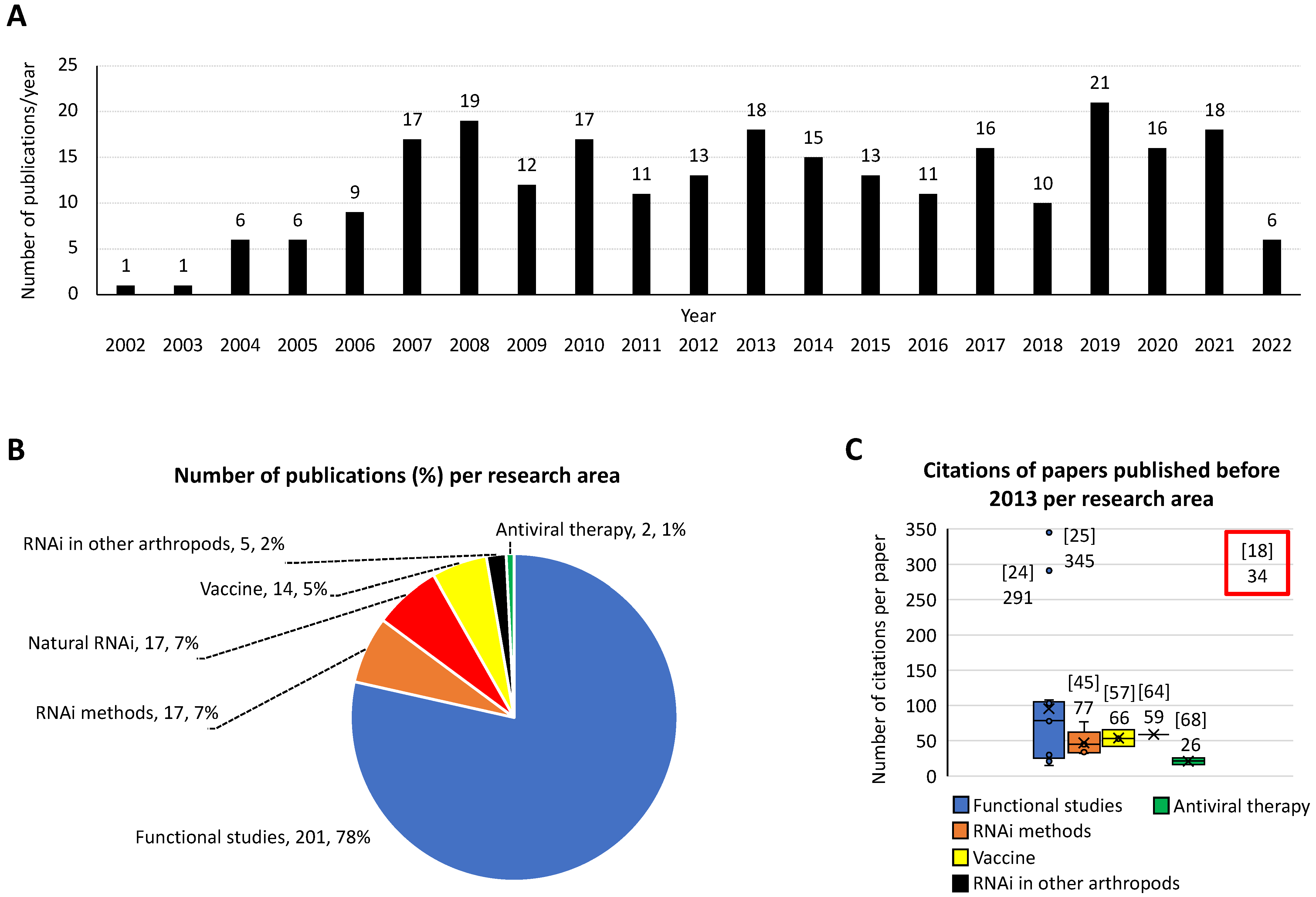
The development and validation of RNAi in ticks has impacted scientific research by advancing studies on a wide variety of arthropod species, ranging from basic biology to biotechnological studies [19]. Using scientometric and bibliometric analyses, 256 publications were identified in PubMed (https://www.ncbi.nlm.nih.gov) searching with “RNA”, “interference”, and “tick” terms on 10 June 2022 (Figure 2A). As shown in this analysis, the number of publications per year increased from one in 2002, the year of the publication of first classic paper [18], to over ten publications per year from 2007–present (Figure 2A).
While the application of RNAi has advanced research in several areas (Figure 2B,C), most of the investigations (201 publications, 78%) with the highest citation scores were focused on tick functional studies. Furthermore, research on naturally occurring RNAi mechanisms and RNAi methods represented 7% (17 publications each) of the total number of publications. Other research areas included vaccines for the control of tick infestations and tick-borne pathogens (14 publications, 5%), RNAi in other arthropods (5, 2%), and antiviral therapy (2, 1%). The examples below illustrate the impact of RNAi on various areas of tick research.
RNAi has been used to address many aspects of tick biology by enabling the functional analysis of individual genes or combinations of genes. Soon after the publication of the classic paper of this commentary, Aljamali et al. [21] reported a silencing of the histamine binding protein (HBP) in A. americanum in vivo and salivary glands, resulting in reduced histamine binding capacity and altered tick feeding. A second paper by Narasimhan et al. 2004 [22] reported a disruption of the anticoagulation response in I. scapularis by silencing the salivary gland gene that expresses the anticoagulant Salp14. In 2006, de la Fuente et al. demonstrated the impact of the silencing of a single gene (subolesin), resulting in sterile male ticks that were unable to mate successfully with female ticks, thus inhibiting completion of female engorgement and oviposition [23]. Collectively, these initial RNAi functional studies in ticks demonstrated the relevance of this methodology for enhancing the understanding of tick biology. Additional subsequent studies have included the knockdown of different tick genes for the characterization of multiple biological pathways involved in tick-host-pathogen interactions [24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41]. In many studies, the results were used to propose new candidate protective antigens for vaccine development or for use as pharmaceutical targets [23,33,34,35,37].
RNA viruses were studied early [42], and RNAi is the major antiviral mechanism against arboviruses in arthropod vectors. As in other organisms, this natural antiviral response was also subsequently characterized in ticks [43,44,45].
RNAi methodology evolved from the original in vitro incubation of dsRNA with tick salivary glands and from in vivo studies in live female ticks injected with dsRNA [18,20]. The variety of methodologies for tick RNAi subsequently included, in chronological order, (a) RNAi in cultured tick cells [46], (b) transovarial RNAi [47], (c) in vitro feeding assays for hard ticks [48], (d) dsRNA electroporation in tick eggs and nymphs [49], (e) cytoplasmic RNA viruses as vehicles for the efficient delivery of therapeutic small RNAs [50], (f) dual luciferase reporter systems for optimization of RNAi [51], (g) non-invasive delivery of dsRNA into de-waxed tick eggs by electroporation [52], (h) tick immersion in dsRNA [53], (i) liposome mediated dsRNA delivery [54], (j) delivery of a genetically marked Serratia AS1 for RNAi [55], (k) functional RNAi analyses using tick organ cultures [56], and (l) cationic glycopolyelectrolytes for RNAi in tick cells [57].
RNAi has also been used to identify and characterize candidate protective antigens for the control of tick infestations and tick-borne pathogens [23,58,59,60,61,62]. In this approach, the selection of protective antigens can be done prior to testing in animal vaccine trials.
Following validation of RNAi in ticks [18], the methodologies were also used in other arthropod species, including the fruit fly Drosophila melanogaster [63], salmon louse Lepeophtheirus salmonis [64], honeybee mite Varroa destructor [65,66], and mosquito Anopheles arabiensis [67]. These studies were focused on the characterization of the function of proteins in the biological processes of the arthropod life cycle, and on pathogen infection and transmission.
The proposal of RNAi-based therapeutics for controlling the tick-borne encephalitis virus (TBE) [68] and other flavivirus [69] infections was also based on the development of antisense-based approaches derived from RNAi research in ticks and other arthropod species.
Recently, RNAi has been incorporated into research on the Alpha-Gal Syndrome (AGS), an IgE-mediated, delayed-type allergic reaction in response to the oligosaccharide galactose-α-1,3-galactose (α-gal). Alpha-gal is present in tick biomolecules and injected into humans during tick feeding [70].
In conclusion, RNAi is the leading gene expression manipulation tool in arthropods followed by gene editing via the bacterial type II Clustered Regularly Interspaced Short Palindromic Repeats and associated protein 9 system (CRISPR-Cas9) [71]. Characterization of the microbiota composition in ticks and other arthropods may also provide targets for the production of modified bacteria using paratransgenesis or RNAi [54,72,73,74]. The recent report of the successful application of CRISPR-Cas9 in ticks [75] provides an opportunity to combine these methodologies for the manipulation of tick gene expression and the development of paratransgenic interventions for the control of tick infestations and transmission of tick-borne pathogens.
Author Contributions
Conceptualization, J.d.l.F. and K.M.K.; methodology, J.d.l.F.; writing—original draft preparation, J.d.l.F. and K.M.K.; writing—review and editing, J.d.l.F. and K.M.K.; visualization, J.d.l.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Alejandro Cabezas-Cruz (UMR BIPAR, INRAE, ANSES, Ecole Nationale Vétérinaire d’Alfort, Université Paris-Est, 94700 Maisons-Alfort, France) for leading this initiative in the Special Issue “10th Anniversary of Pathogens—Classic Papers in Tick Research”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sen, G.L.; Blau, H.M. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. Overview ofRNAInterference and Related Processes. Curr. Protoc. Mol. Biol. 2003, 62, 26.1.1–26.1.6. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K. RNA interference: Historical overview and significance. Methods Mol. Biol. 2004, 265, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a Chimeric Chalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in trans. Plant Cell 1990, 2, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Romano, N.; Macino, G. Quelling: Transient inactivation of gene expression in Neurospora crassa by transformation with homologous sequences. Mol. Microbiol. 1992, 6, 3343–3353. [Google Scholar] [CrossRef]
- Guo, S.; Kemphues, K.J. par-1, a gene required for establishing polarity in C. elegans embryos, encodes a putative Ser/Thr kinase that is asymmetrically distributed. Cell 1995, 81, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Grishok, A.; Tabara, H.; Mello, C.C. Genetic Requirements for Inheritance of RNAi in C. elegans. Science 2000, 287, 2494–2497. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O.; Baulcombe, D. Systemic signalling in gene silencing. Nature 1997, 389, 553. [Google Scholar] [CrossRef]
- Hamilton, A.J.; Baulcombe, D.C. A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [Green Version]
- Hammond, S.M.; Bernstein, E.; Beach, D.; Hannon, G.J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 2000, 404, 293–296. [Google Scholar] [CrossRef]
- Zamore, P.D.; Tuschl, T.; Sharp, P.A.; Bartel, D.P. RNAi: Double-Stranded RNA Directs the ATP-Dependent Cleavage of mRNA at 21 to 23 Nucleotide Intervals. Cell 2000, 101, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Elbashir, S.M.; Lendeckel, W.; Tuschl, T. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Dev. 2001, 15, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar] [CrossRef]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef]
- Martinez, J.; Patkaniowska, A.; Urlaub, H.; Lührmann, R.; Tuschl, T. Single-Stranded Antisense siRNAs Guide Target RNA Cleavage in RNAi. Cell 2002, 110, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Hutvágner, G.; Zamore, P.D. RNAi: Nature abhors a double-strand. Curr. Opin. Genet. Dev. 2002, 12, 225–232. [Google Scholar] [CrossRef]
- Aljamali, M.N.; Sauer, J.R.; Essenberg, R.C. RNA Interference: Applicability in Tick Research. Exp. Appl. Acarol. 2002, 28, 89–96. [Google Scholar] [CrossRef]
- de la Fuente, J.; Kocan, K.M.; Almazán, C.; Blouin, E.F. RNA interference for the study and genetic manipulation of ticks. Trends Parasitol. 2007, 23, 427–433. [Google Scholar] [CrossRef]
- Kocan, K.M.; Blouin, E.; De La Fuente, J. RNA Interference in Ticks. J. Vis. Exp. 2011, e2474. [Google Scholar] [CrossRef] [Green Version]
- Aljamali, M.N.; Bior, A.D.; Sauer, J.R.; Essenberg, R.C. RNA interference in ticks: A study using histamine binding protein dsRNA in the female tick Amblyomma americanum. Insect Mol. Biol. 2003, 12, 299–305. [Google Scholar] [CrossRef]
- Narasimhan, S.; Montgomery, R.R.; DePonte, K.; Tschudi, C.; Marcantonio, N.; Anderson, J.F.; Sauer, J.R.; Cappello, M.; Kantor, F.S.; Fikrig, E. Disruption of Ixodes scapularis anticoagulation by using RNA interference. Proc. Natl. Acad. Sci. USA 2004, 101, 1141–1146. [Google Scholar] [CrossRef] [Green Version]
- de la Fuente, J.; Almazán, C.; Naranjo, V.; Blouin, E.F.; Meyer, J.M.; Kocan, K.M. Autocidal control of ticks by silencing of a single gene by RNA interference. Biochem. Biophys. Res. Commun. 2006, 344, 332–338. [Google Scholar] [CrossRef]
- Karim, S.; Ramakrishnan, V.G.; Tucker, J.S.; Essenberg, R.C.; Sauer, J.R. Amblyomma americanum salivary glands: Double-stranded RNA-mediated gene silencing of synaptobrevin homologue and inhibition of PGE2 stimulated protein secretion. Insect Biochem. Mol. Biol. 2004, 34, 407–413. [Google Scholar] [CrossRef]
- Pal, U.; Li, X.; Wang, T.; Montgomery, R.R.; Ramamoorthi, N.; Desilva, A.M.; Bao, F.; Yang, X.; Pypaert, M.; Pradhan, D.; et al. TROSPA, an Ixodes scapularis Receptor for Borrelia burgdorferi. Cell 2004, 119, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorthi, N.; Narasimhan, S.; Pal, U.; Bao, F.; Yang, X.F.; Fish, D.; Anguita, J.; Norgard, M.V.; Kantor, F.S.; Anderson, J.F.; et al. The Lyme disease agent exploits a tick protein to infect the mammalian host. Nature 2005, 436, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Kenny, B.; Troiano, E.; Mather, T.N. RNAi-mediated gene silencing in tick synganglia: A proof of concept study. BMC Biotechnol. 2008, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Hajdusek, O.; Sojka, D.; Kopacek, P.; Buresova, V.; Franta, Z.; Sauman, I.; Winzerling, J.; Grubhoffer, L. Knockdown of proteins involved in iron metabolism limits tick reproduction and development. Proc. Natl. Acad. Sci. USA 2009, 106, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Booth, C.J.; Paley, M.A.; Wang, X.; DePonte, K.; Fikrig, E.; Narasimhan, S.; Montgomery, R.R. Inhibition of Neutrophil Function by Two Tick Salivary Proteins. Infect. Immun. 2009, 77, 2320–2329. [Google Scholar] [CrossRef] [Green Version]
- Mulenga, A.; Khumthong, R. Silencing of three Amblyomma americanum (L.) insulin-like growth factor binding protein-related proteins prevents ticks from feeding to repletion. J. Exp. Biol. 2010, 213, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Narasimhan, S.; Zhang, L.; Liu, L.; Wang, P.; Fikrig, E. Tick Histamine Release Factor Is Critical for Ixodes scapularis Engorgement and Transmission of the Lyme Disease Agent. PLoS Pathog. 2010, 6, e1001205. [Google Scholar] [CrossRef] [PubMed]
- Aung, K.M.; Boldbaatar, D.; Umemiya-Shirafuji, R.; Liao, M.; Xuenan, X.; Suzuki, H.; Galay, R.L.; Tanaka, T.; Fujisaki, K. Scavenger Receptor Mediates Systemic RNA Interference in Ticks. PLoS ONE 2011, 6, e28407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, F.A.; Pohl, P.C.; Gandara, A.C.; Ferreira, J.D.S.; Nascimento-Silva, M.C.; Bechara, G.H.; Sorgine, M.H.F.; Almeida, I.C.; Vaz, I.D., Jr.; Oliveira, P.L. ATP Binding Cassette Transporter Mediates Both Heme and Pesticide Detoxification in Tick Midgut Cells. PLoS ONE 2015, 10, e0134779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzugaray, M.F.; Parizi, L.F.; Seixas, A.; Benavides, U.; Vaz, I.D.S. Molecular and functional characterization of Bm05br antigen from Rhipicephalus microplus. Ticks Tick-Borne Dis. 2017, 8, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.M.; Gulia-Nuss, M.; Kuhn, R.J.; Hill, C.A. RNAi reveals proteins for metabolism and protein processing associated with Langat virus infection in Ixodes scapularis (black-legged tick) ISE6 cells. Parasites Vectors 2017, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narasimhan, S.; Schuijt, T.J.; Abraham, N.M.; Rajeevan, N.; Coumou, J.; Graham, M.; Robson, A.; Wu, M.-J.; Daffre, S.; Hovius, J.W.; et al. Modulation of the tick gut milieu by a secreted tick protein favors Borrelia burgdorferi colonization. Nat. Commun. 2017, 8, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowski, J.M.; Tsetsarkin, K.A.; Long, D.; Scott, D.P.; Rosenke, R.; Schwan, T.G.; Mlera, L.; Offerdahl, D.K.; Pletnev, A.G.; Bloom, M.E. Flavivirus Infection of Ixodes scapularis (Black-Legged Tick) Ex Vivo Organotypic Cultures and Applications for Disease Control. mBio 2017, 8, e01255-17. [Google Scholar] [CrossRef] [Green Version]
- Carroll, E.E.M.; Wang, X.; Shaw, D.K.; O’Neal, A.J.; Chávez, A.S.O.; Brown, L.J.; Boradia, V.M.; Hammond, H.L.; Pedra, J.H.F. p47 licenses activation of the immune deficiency pathway in the tick Ixodes scapularis. Proc. Natl. Acad. Sci. USA 2019, 116, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Hussein, H.E.; Johnson, W.C.; Taus, N.S.; Suarez, C.E.; Scoles, G.A.; Ueti, M.W. Silencing expression of the Rhipicephalus microplus vitellogenin receptor gene blocks Babesia bovis transmission and interferes with oocyte maturation. Parasites Vectors 2019, 12, 7. [Google Scholar] [CrossRef]
- Wu, F.; Luo, J.; Chen, Z.; Ren, Q.; Xiao, R.; Liu, W.; Hao, J.; Liu, X.; Guo, J.; Qu, Z.; et al. MicroRNA let-7 regulates the expression of ecdysteroid receptor (ECR) in Hyalomma asiaticum (Acari: Ixodidae) ticks. Parasites Vectors 2019, 12, 235. [Google Scholar] [CrossRef]
- Agwunobi, D.O.; Wang, N.; Huang, L.; Zhang, Y.; Chang, G.; Wang, K.; Li, M.; Wang, H.; Liu, J. Phosphoproteomic Analysis of Haemaphysalis longicornis Saliva Reveals the Influential Contributions of Phosphoproteins to Blood-Feeding Success. Front. Cell. Infect. Microbiol. 2022, 11, 769026. [Google Scholar] [CrossRef]
- Andzhaparidze, O.G.; Bogomolova, N.N.; Boriskin, I.; Desiatskova, R.G.; Bektemirova, M.S. Sravnitel’noe izuchenie ersistentsii trekh RNK-soderzhashchikh virusov v perevivaemoĭ kul’ture kletok chelovecheskogo proiskhozhdeniia [Comparative study of the persistence of 3 RNA-containing viruses in a continuous cell culture of human origin]. Vopr. Virusol. 1980, 3, 323–327. [Google Scholar]
- Schnettler, E.; Tykalová, H.; Watson, M.; Sharma, M.; Sterken, M.; Obbard, D.; Lewis, S.H.; McFarlane, M.; Bell-Sakyi, L.; Barry, G.; et al. Induction and suppression of tick cell antiviral RNAi responses by tick-borne flaviviruses. Nucleic Acids Res. 2014, 42, 9436–9446. [Google Scholar] [CrossRef]
- Hart, C.E.; Thangamani, S. Tick-virus interactions: Current understanding and future perspectives. Parasite Immunol. 2021, 43, e12815. [Google Scholar] [CrossRef]
- Xu, Y.; Zhong, Z.; Ren, Y.; Ma, L.; Ye, Z.; Gao, C.; Wang, J.; Li, Y. Antiviral RNA interference in disease vector (Asian longhorned) ticks. PLoS Pathog. 2021, 17, e1010119. [Google Scholar] [CrossRef]
- de la Fuente, J.; Blouin, E.F.; Manzano-Roman, R.; Naranjo, V.; Almazán, C.; de la Lastra, J.M.P.; Zivkovic, Z.; Jongejan, F.; Kocan, K.M. Functional genomic studies of tick cells in response to infection with the cattle pathogen, Anaplasma marginale. Genomics 2007, 90, 712–722. [Google Scholar] [CrossRef] [Green Version]
- Kocan, K.M.; Manzano-Roman, R.; De La Fuente, J. Transovarial silencing of the subolesin gene in three-host ixodid tick species after injection of replete females with subolesin dsRNA. Parasitol. Res. 2007, 100, 1411–1415. [Google Scholar] [CrossRef]
- Kröber, T.; Guerin, P.M. In vitro feeding assays for hard ticks. Trends Parasitol. 2007, 23, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Troiano, E.; Mather, T.N. Functional genomics tool: Gene silencing in Ixodes scapularis eggs and nymphs by electroporated dsRNA. BMC Biotechnol. 2010, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Usme-Ciro, J.A.; Campillo-Pedroza, N.; Almazán, F.; Gallego-Gomez, J.C. Cytoplasmic RNA viruses as potential vehicles for the delivery of therapeutic small RNAs. Virol. J. 2013, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Tuckow, A.P.; Temeyer, K.B. Discovery, adaptation and transcriptional activity of two tick promoters: Construction of a dual luciferase reporter system for optimization of RNA interference in Rhipicephalus (Boophilus) microplus cell lines. Insect Mol. Biol. 2015, 24, 454–466. [Google Scholar] [CrossRef]
- Ruiz, N.; De Abreu, L.A.; Parizi, L.F.; Kim, T.K.; Mulenga, A.; Braz, G.R.C.; Vaz, I.D.S.; Logullo, C. Non-Invasive Delivery of dsRNA into De-Waxed Tick Eggs by Electroporation. PLoS ONE 2015, 10, e0130008. [Google Scholar] [CrossRef]
- Galay, R.L.; Hernandez, E.P.; Talactac, M.R.; Maeda, H.; Kusakisako, K.; Umemiya-Shirafuji, R.; Mochizuki, M.; Fujisaki, K.; Tanaka, T. Induction of gene silencing in Haemaphysalis longicornis ticks through immersion in double-stranded RNA. Ticks Tick-Borne Dis. 2016, 7, 813–816. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, J.; Zhou, Y.; Cao, J.; Gong, H.; Zhang, H.; Zhou, J. Liposome mediated double-stranded RNA delivery to silence ribosomal protein P0 in the tick Rhipicephalus haemaphysaloides. Ticks Tick-Borne Dis. 2018, 9, 638–644. [Google Scholar] [CrossRef]
- Koosha, M.; Vatandoost, H.; Karimian, F.; Choubdar, N.; Oshaghi, M.A. Delivery of a Genetically Marked Serratia AS1 to Medically Important Arthropods for Use in RNAi and Paratransgenic Control Strategies. Microb. Ecol. 2019, 78, 185–194. [Google Scholar] [CrossRef]
- Grabowski, J.M.; Kissinger, R. Ixodid Tick Dissection and Tick Ex Vivo Organ Cultures for Tick-Borne Virus Research. Curr. Protoc. Microbiol. 2020, 59, e118. [Google Scholar] [CrossRef]
- Stockmal, K.A.; Downs, L.P.; Davis, A.N.; Kemp, L.K.; Karim, S.; Morgan, S.E. Cationic Glycopolyelectrolytes for RNA Interference in Tick Cells. Biomacromolecules 2022, 23, 34–46. [Google Scholar] [CrossRef]
- De La Fuente, J.; Almazán, C.; Blouin, E.F.; Naranjo, V.; Kocan, K.M. RNA interference screening in ticks for identification of protective antigens. Parasitol. Res. 2005, 96, 137–141. [Google Scholar] [CrossRef]
- De La Fuente, J.; Almazán, C.; Naranjo, V.; Blouin, E.F.; Kocan, K.M. Synergistic effect of silencing the expression of tick protective antigens 4D8 and Rs86 in Rhipicephalus sanguineus by RNA interference. Parasitol. Res. 2006, 99, 108–113. [Google Scholar] [CrossRef]
- Merino, O.; Almazán, C.; Canales, M.; Villar, M.; Moreno-Cid, J.A.; Peña, A.E.; Kocan, K.M.; De La Fuente, J. Control of Rhipicephalus (Boophilus) microplus infestations by the combination of subolesin vaccination and tick autocidal control after subolesin gene knockdown in ticks fed on cattle. Vaccine 2011, 29, 2248–2254. [Google Scholar] [CrossRef]
- Mulenga, A.; Kim, T.K.; Ibelli, A.M.G. Deorphanization and target validation of cross-tick species conserved novel Amblyomma americanum tick saliva protein. Int. J. Parasitol. 2013, 43, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjunathachar, H.V.; Azhahianambi, P.; Kumar, B.; Ghosh, S. Screening for the “Achilles Heel” of Hyalomma anatolicum Ticks by RNA Interference Technology and an Update on Anti-Tick Vaccine Design. Methods Mol. Biol. 2022, 2411, 307–330. [Google Scholar] [CrossRef] [PubMed]
- Drolia, R.; Von Ohlen, T.; Chapes, S.K. Ehrlichia chaffeensis replication sites in adult Drosophila melanogaster. Int. J. Med Microbiol. 2013, 303, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichner, C.; Øvergård, A.-C.; Nilsen, F.; Dalvin, S. Molecular characterization and knock-down of salmon louse (Lepeophtheirus salmonis) prostaglandin E synthase. Exp. Parasitol. 2015, 159, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.M.; Budge, G.E.; Bowman, A.S. Gene-knockdown in the honey bee mite Varroa destructor by a non-invasive approach: Studies on a glutathione S-transferase. Parasites Vectors 2010, 3, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.Y.; Bian, G.; Xi, Z.; Xie, X. Genes important for survival or reproduction inVarroa destructoridentified by RNAi. Insect Sci. 2019, 26, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Letinić, B.D.; Dahan-Moss, Y.; Koekemoer, L.L. Characterising the effect of Akirin knockdown on Anopheles arabiensis (Diptera: Culicidae) reproduction and survival, using RNA-mediated interference. PLoS ONE 2020, 15, e0228576. [Google Scholar] [CrossRef] [Green Version]
- Achazi, K.; Patel, P.; Paliwal, R.; Radonić, A.; Niedrig, M.; Donoso-Mantke, O. RNA interference inhibits replication of tick-borne encephalitis virus in vitro. Antivir. Res. 2012, 93, 94–100. [Google Scholar] [CrossRef] [Green Version]
- David, A.S.; Stein, D.A.; Shi, P.-Y. Nucleic acid-based inhibition of flavivirus infections. Front. Biosci. 2008, 13, 1385–1395. [Google Scholar] [CrossRef]
- Sharma, S.R.; Crispell, G.; Mohamed, A.; Cox, C.; Lange, J.; Choudhary, S.; Commins, S.P.; Karim, S. Alpha-Gal Syndrome: Involvement of Amblyomma americanum α-D-Galactosidase and β-1,4 Galactosyltransferase Enzymes in α-Gal Metabolism. Front. Cell. Infect. Microbiol. 2021, 11, 775371. [Google Scholar] [CrossRef]
- Nganso, B.T.; Pines, G.; Soroker, V. Insights into gene manipulation techniques for Acari functional genomics. Insect Biochem. Mol. Biol. 2022, 143, 103705. [Google Scholar] [CrossRef]
- Aguilar-Díaz, H.; Quiroz-Castañeda, R.E.; Cobaxin-Cárdenas, M.; Salinas-Estrella, E.; Amaro-Estrada, I. Advances in the Study of the Tick Cattle Microbiota and the Influence on Vectorial Capacity. Front. Vet. Sci. 2021, 8, 710352. [Google Scholar] [CrossRef]
- Choubdar, N.; Karimian, F.; Koosha, M.; Oshaghi, M.A. An integrated overview of the bacterial flora composition of Hyalomma anatolicum, the main vector of CCHF. PLoS Negl. Trop. Dis. 2021, 15, e0009480. [Google Scholar] [CrossRef]
- de la Fuente, J. Translational biotechnology for the control of ticks and tick-borne diseases. Ticks Tick-borne Dis. 2021, 12, 101738. [Google Scholar] [CrossRef]
- Sharma, A.; Pham, M.N.; Reyes, J.B.; Chana, R.; Yim, W.C.; Heu, C.C.; Kim, D.; Chaverra-Rodriguez, D.; Rasgon, J.L.; Harrell, R.A.; et al. Cas9-mediated gene editing in the black-legged tick, Ixodes scapularis, by embryo injection and ReMOT Control. iScience 2022, 25, 103781. [Google Scholar] [CrossRef]
Figure 1.
Model of the mechanism of RNAi applications in tick biology. Both model and applications were inspired in the classic paper by Aljamali et al., 2002 [18]. Ixodes scapularis tick and ISE6 cell images are courtesy of the authors.
Figure 1.
Model of the mechanism of RNAi applications in tick biology. Both model and applications were inspired in the classic paper by Aljamali et al., 2002 [18]. Ixodes scapularis tick and ISE6 cell images are courtesy of the authors.

Figure 2.
Scientometric and bibliometric analyses of RNAi applications in ticks and other arthropod species. Analyses were conducted based on publications identified in PubMed (https://www.ncbi.nlm.nih.gov; accessed on 10 June 2022) using search with “RNA” and “interference” and “tick” terms on 10 June 2022. (A) Number of publications per year. (B) Number of publications and percentage per research area. (C) Number of citations per paper published before 2013 (in the first decade after the classic paper by Aljamali et al., 2002 [18] was published) per research area. Reference to the papers with highest citation score with corresponding number of citations at Web of Science (https://www.webofscience.com) accessed on 10 June 2022, are shown. The citations of the classic paper by Aljamali et al., 2002 [18], highlighted in this commentary, also shown.
Figure 2.
Scientometric and bibliometric analyses of RNAi applications in ticks and other arthropod species. Analyses were conducted based on publications identified in PubMed (https://www.ncbi.nlm.nih.gov; accessed on 10 June 2022) using search with “RNA” and “interference” and “tick” terms on 10 June 2022. (A) Number of publications per year. (B) Number of publications and percentage per research area. (C) Number of citations per paper published before 2013 (in the first decade after the classic paper by Aljamali et al., 2002 [18] was published) per research area. Reference to the papers with highest citation score with corresponding number of citations at Web of Science (https://www.webofscience.com) accessed on 10 June 2022, are shown. The citations of the classic paper by Aljamali et al., 2002 [18], highlighted in this commentary, also shown.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
de la Fuente, J.; Kocan, K.M. The Impact of RNA Interference in Tick Research. Pathogens 2022, 11, 827. https://doi.org/10.3390/pathogens11080827
AMA Style
de la Fuente J, Kocan KM. The Impact of RNA Interference in Tick Research. Pathogens. 2022; 11(8):827. https://doi.org/10.3390/pathogens11080827
Chicago/Turabian Stylede la Fuente, José, and Katherine M. Kocan. 2022. "The Impact of RNA Interference in Tick Research" Pathogens 11, no. 8: 827. https://doi.org/10.3390/pathogens11080827
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.