
Temporal Incidence of Eriophyid Mites on Rose Rosette Disease-Symptomatic and -Asymptomatic Roses in Central Georgia, USA


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Site and Plant
2.2. Experimental Design and Sample Collection
2.3. Mite Identification
2.4. Statistical Analyses
3. Results
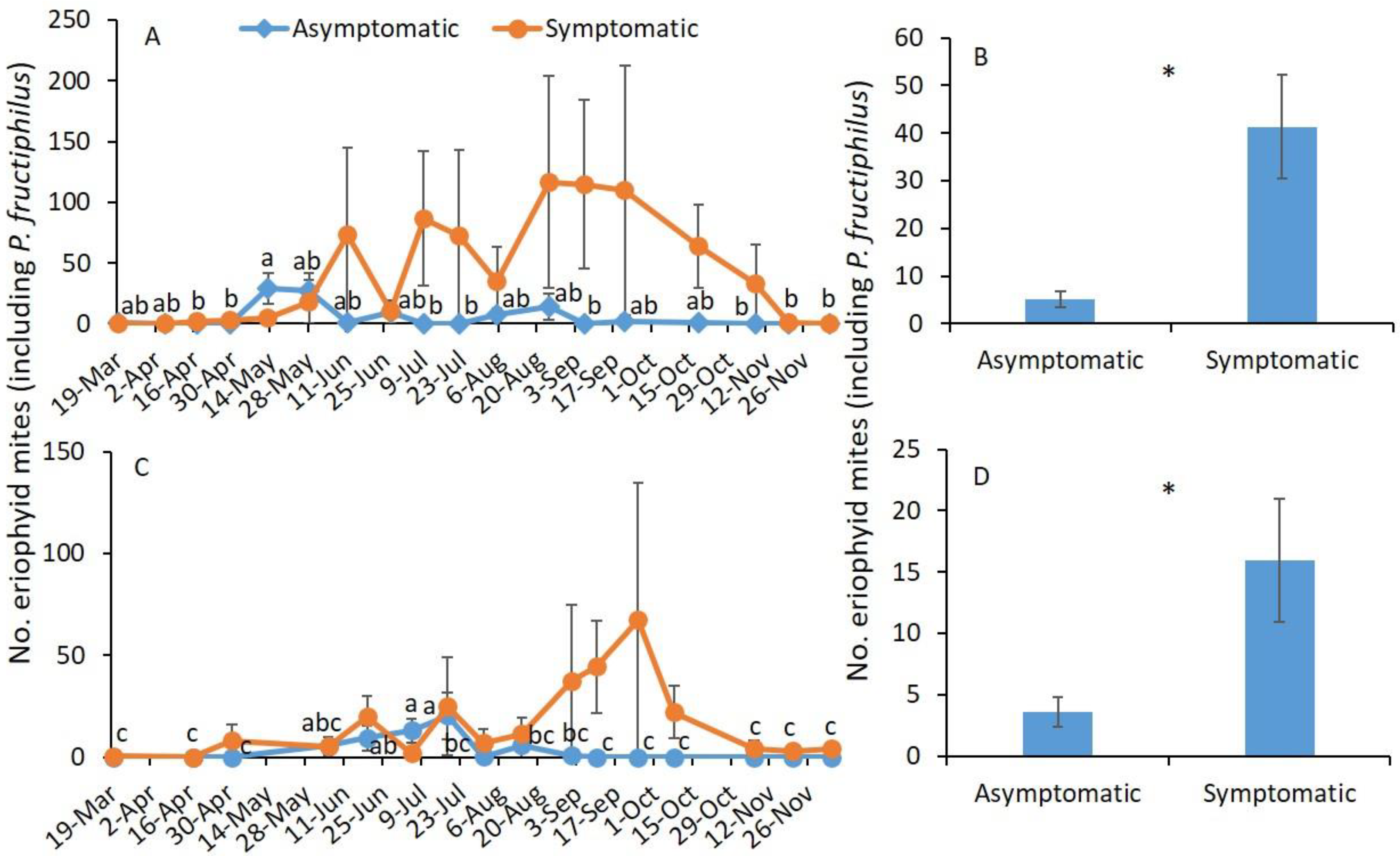
3.1. Sampling Date and RRD Status
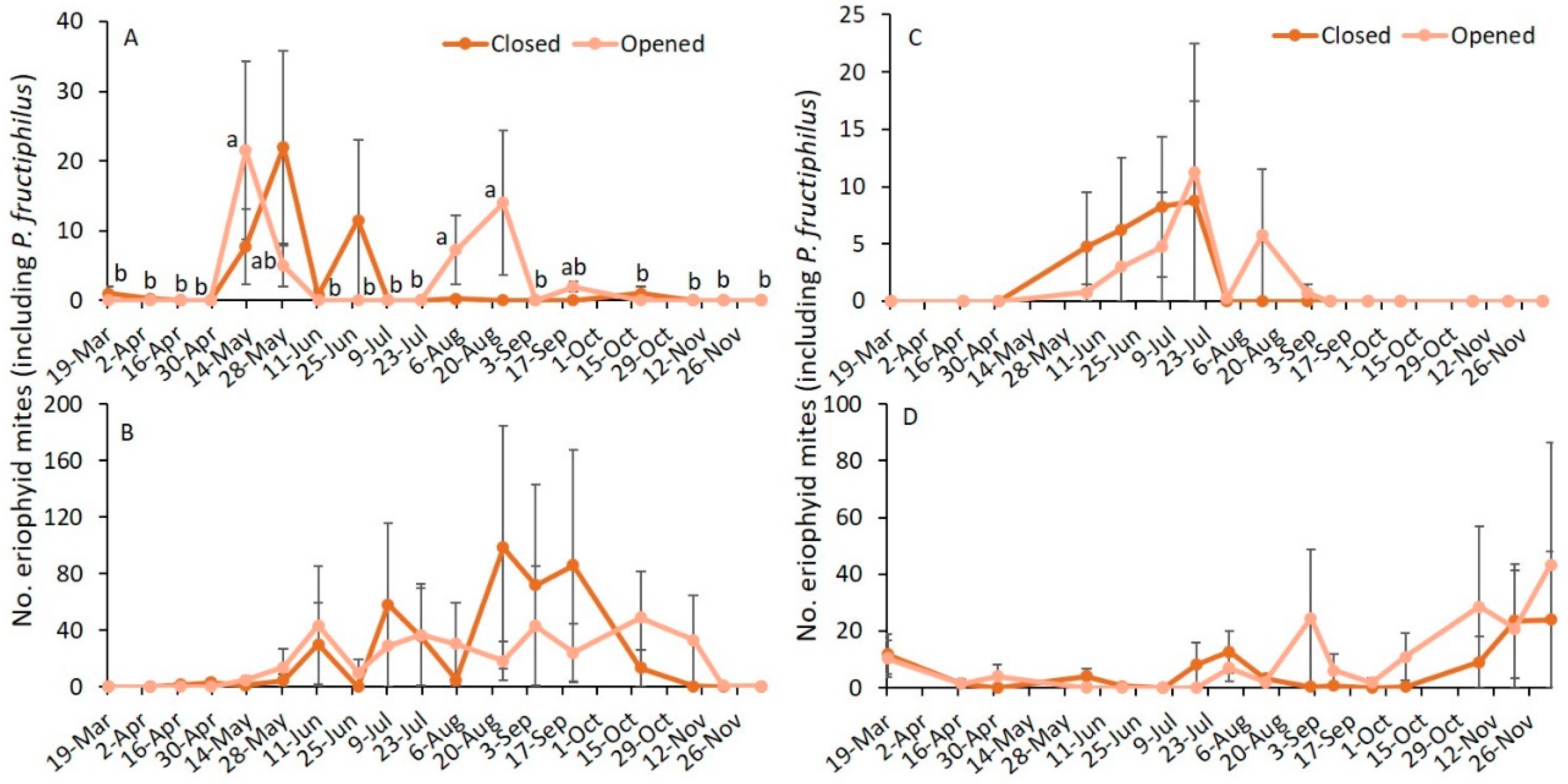
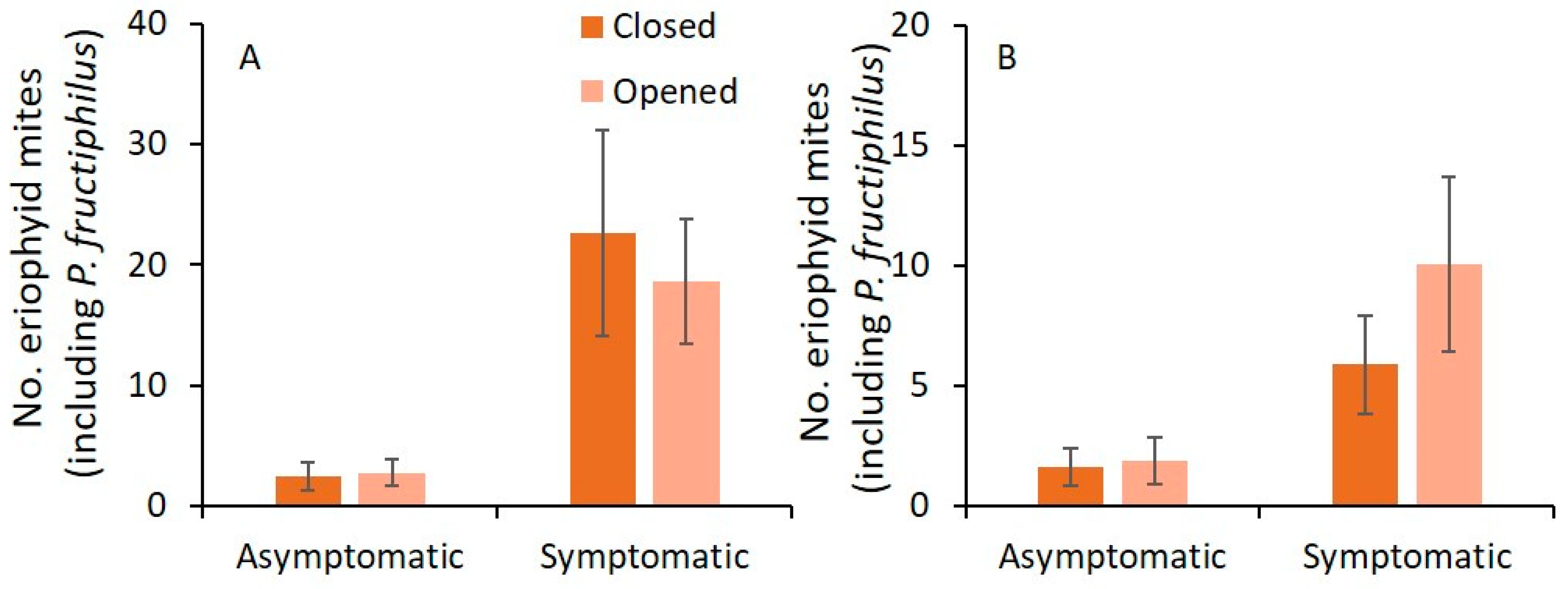
3.2. Sampling Date and Closed or Opened Flower Buds
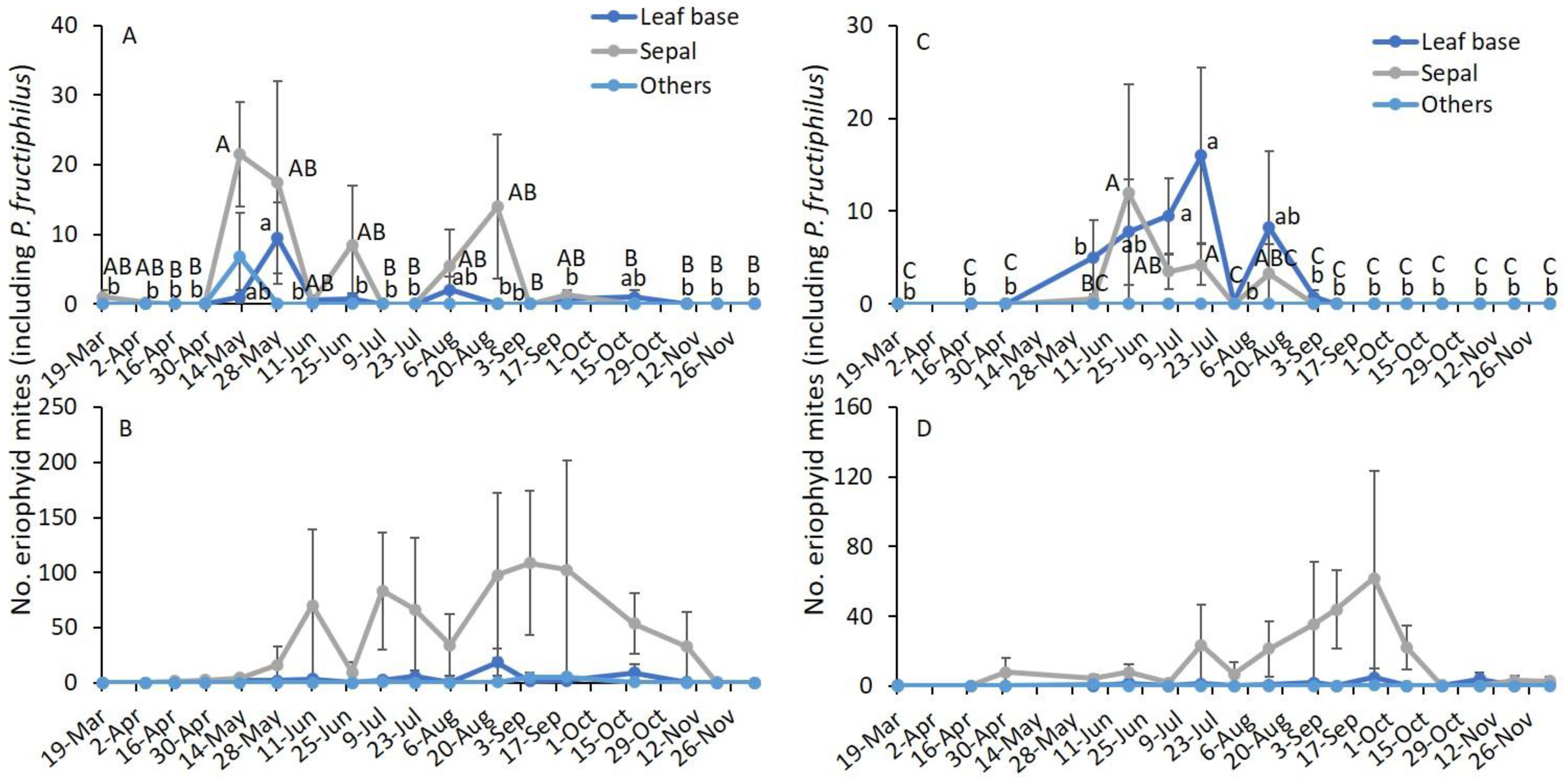
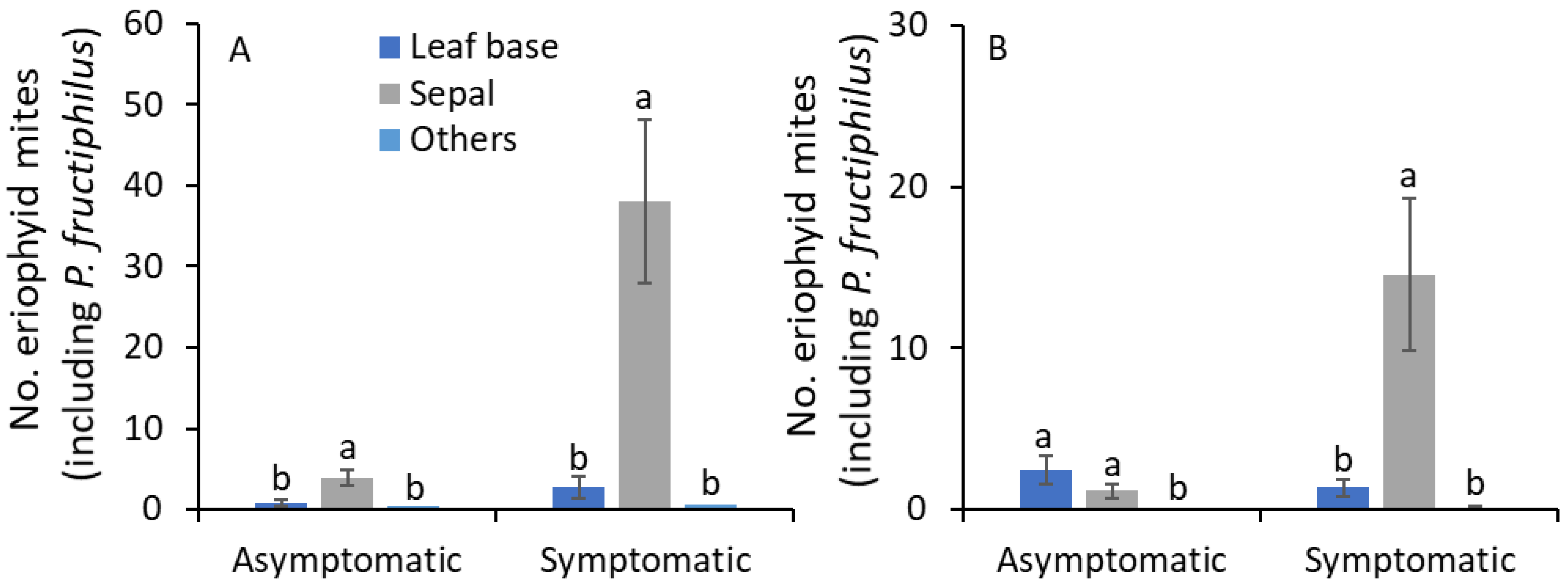
3.3. Sampling Date and Terminal Plant Parts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Bello, P.L.; Thekke-Veetil, T.; Druciarek, T.; Tzanetakis, I.E. Transmission attributes and resistance to rose rosette virus. Plant Path. 2018, 67, 499–504. [Google Scholar] [CrossRef]
- Amrine, J.W., Jr.; Hindal, D.F.; Stasny, T.A.; Williams, R.L.; Coffman, C.C. Transmission of the rose rosette disease agent to Rosa multiflora by Phyllocoptes fructiphilus (Acari: Eriophyidae). Entomol. News 1988, 99, 239–252. [Google Scholar]
- Windham, M.; Windham, A.; Hale, F.; Amrine, J. Observations on rose rosette disease. Am. Rose 2014, 42, 56–62. [Google Scholar]
- USDA-NASS. 2015. Available online: https://www.nass.usda.gov/Publications/AgCensus/2012/Online_Resources/Census_of_Horticulture_Specialties/HORTIC.pdf (accessed on 1 February 2022).
- Amrine, J.W., Jr. Phyllocoptes fructiphilus and biological control of multiflora rose. In Eriophyoid Mites—Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publ.: Amsterdam, The Netherlands, 1996; pp. 741–749. [Google Scholar]
- Otero-Colina, G.; Ochoa, R.; Armine, J.W., Jr.; Hammond, J.; Jordan, R.; Bauchan, G.R. Eriophyid mites found on healthy and rose rosette diseased roses in the United States. J. Environ. Hort. 2018, 36, 146–153. [Google Scholar]
- Allington, W.B.; Staples, R.; Viehmeyer, G. Transmission of Rose Rosette Virus by the eriophid mite Phyllocoptes fructiphilus. J. Econ. Entomol. 1968, 61, 1137–1140. [Google Scholar] [CrossRef]
- Druciarek, T.; Lewandowski, M. A new species of eriophyoid mite (Acari: Eriophyoidea) on Rosa sp. from Israel. Zootaxa 2016, 4066, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Solo, K.; Collins, S.; Shires, M.; Ochoa, R.; Bauchan, G.; Schneider, L.; Henn, A.; Jacobi, J.; Williams-Woodward, J.; Hajimorad, M.R.; et al. A survey of rose rosette virus and eriophyid mites associated with roses in the southeastern United States. HortScience 2020, 55, 1–7. [Google Scholar] [CrossRef]
- Kassar, A.; Armine, J.W. Rearing and development of Phyllocoptes fructiphilus (Acari: Eriophyidae). Entomol. News 1990, 101, 276–282. [Google Scholar]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Hoy, M. Eriophyid mite vector of Rose rosette disease (RRD), Phyllocoptes fructiphilus Keifer (Arachnida: Acari: Eriophyidae). EENY-558. 2013. Available online: http://entnemdept.ufl.edu/creatures/ORN/ph_fructiphilus.htm (accessed on 1 February 2022).
- Swirski, E. Contribution to the knowledge of the fluctuations in population of the citrus rust mite (Phyllocopturata oleivora Ashm.) in the coastal plain of Israel. Israel J. Agric. Res. 1962, 12, 175–187. [Google Scholar]
- Ozman, S.K.; Goolsby, J.A. Biology and phenology of the eriophyid mite, Floracarus perrepae, on its native host in Australia, Old World climbing fern, Lygodium microphyllum. Exp. Appl. Acarol. 2005, 35, 197. [Google Scholar] [CrossRef] [PubMed]
- Mahr, F.A.; Kwong, R.M.; McClaren, D.A.; Jupp, P.W. Redistribution and present status of the mite Aculus hyperici (Acari: Eriophyidae) for the control of St. John’s wort Hypericum perforatum Australia. Plant Prot. Q. 1997, 12, 84–88. [Google Scholar]
- Babu, B.; Jeyaprakash, A.; Jones, D.; Schubert, T.S.; Baker, C.; Washburn, B.K.; Miller, S.H.; Poduch, K.; Knox, G.W.; Ochoa-Corona, F.M.; et al. Development of a rapid, sensitive TaqMan real-time RT-PCR assay for the detection of rose rosette virus using multiple gene targets. J. Virol. Methods 2016, 235, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fife, A.; Bolton, S.; Griesheimer, J.L.; Paret, M.; Martini, X. First report of Phyllocoptes fructiphilus Keifer (Eriophyidae), the vector of the rose rosette virus, in Florida, USA. Fla. Entomol. 2020, 103, 411–414. [Google Scholar] [CrossRef]
- Monfreda, R.; Nuzzaci, G.; De Lillo, E. Detection, extraction, and collection of eriophyoid mites. Zootaxa 2007, 1662, 35–43. [Google Scholar]
- Bauchan, G.B.; Otero-Colina, G.; Hammond, J.; Ochoa, R. Rose rosette disease: It all started with a small mite. In Acta Horticulturae, Proceedings of the VII International Symposium on Rose Research and Cultivation, Angers, France, 2–7 July 2019; Foucher, F., Ed.; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2017; Volume 1232, pp. 227–232. [Google Scholar]
- SAS Institute. SAS Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Druciarek, T.; Lewandowski, M.; Tzanetakis, I. Molecular phylogeny of Phyllocoptes associated with roses discloses the presence of a new species. Infect. Genet. Evol. 2021, 95, 105051. [Google Scholar] [CrossRef] [PubMed]
- Epstein, A.H.; Hill, J.H. Status of rose rosette disease as a biological control for multiflora rose. Plant Dis. 1999, 83, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Jesse, L.C.; Moloney, K.; Obrycki, J. Abundance of arthropods on the branch tips of the invasive plant, Rosa multiflora (Rosaceae). Weed Biol. Manag. 2006, 6, 204–211. [Google Scholar] [CrossRef]
- Craemer, C. A systematic appraisal of the Eriophyoidea (Acari: Prostigmata). Ph.D. Dissertation, Faculty of Natural and Agricultural Sciences, University of Pretoria, Pretoria, South Africa, November 2010. [Google Scholar]
- Amrine, J.W. Multiflora rose. In Biological Control of Invasive Plants in the Eastern United States; Van Driesche, R., Blossey, B., Hoddle, M., Lyon, S., Reardon, R., Eds.; Forest Health Technology Enterprise Team: Morgantown, WV, USA; USDA Publications: Washington, DC, USA, 2002; pp. 265–292. [Google Scholar]
- Shi, A.; Tomczyk, A. Impact of feeding of eriophyid mite Epitrimerus gibbosus (Nalepa) (Acari: Eriophyoidea) on some biochemical components of blackberry (Rubus spp.). Bull. Pol. Acad. Sci. Biol. Sci. 2001, 49, 41–47. [Google Scholar]
- Kumar, N.; Srivastava, G.C.; Kiran, D.; Aarti, M.; Madan, P. Role of carbohydrates in flower bud opening in rose (Rosa hybrida L.). J. Hort. Sci. Biotech. 2007, 82, 235–242. [Google Scholar] [CrossRef]
- Schmitzer, V.; Veberic, R.; Osterc, G.; Stampar, F. Changes in the phenolic concentration during flower development of Rose ‘KORcrisett’. J. Am. Soc. Hort. Sci. 2009, 134, 491–496. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatment | F | df | p |
|---|---|---|---|---|
| 2019 | ||||
| Sampling date | 1.8 | 17,105 | 0.029 | |
| RRD status * | 13.3 | 1105 | 0.004 | |
| Sampling date × RRD status | 1.5 | 17,105 | 0.112 | |
| 2020 | ||||
| Sampling date | 1.5 | 15,101 | 0.106 | |
| RRD status | 9.9 | 1101 | 0.002 | |
| Sampling date × RRD status | 1.1 | 15,101 | 0.359 |
| Variables | 2019 | 2020 | ||||
|---|---|---|---|---|---|---|
| F | df | p | F | df | p | |
| Asymptomatic | ||||||
| Sampling date | 3.1 | 17,105 | <0.001 | 1.5 | 15,101 | 0.109 |
| Flower bud status | 0.3 | 1105 | 0.573 | 0.0 | 1101 | 0.963 |
| Sampling date × flower bud status | 1.2 | 17,105 | 0.319 | 0.2 | 15,101 | 0.999 |
| Symptomatic | ||||||
| Sampling date | 1.4 | 17,105 | 0.157 | 1.2 | 15,101 | 0.307 |
| Flower bud status * | 0.5 | 1105 | 0.499 | 0.0 | 1101 | 0.898 |
| Sampling date × flower bud status | 0.3 | 17,105 | 0.993 | 0.5 | 15,101 | 0.947 |
| Variables | 2019 | 2020 | ||||
|---|---|---|---|---|---|---|
| F | df | p | F | df | p | |
| Asymptomatic | ||||||
| Sampling date | 4.4 | 17,159 | <0.001 | 4.1 | 16,150 | <0.001 |
| Plant parts * | 8.4 | 17,159 | <0.001 | 7.2 | 2150 | 0.001 |
| Sampling date × plant parts | 1.5 | 34,159 | 0.047 | 1.4 | 32,150 | 0.098 |
| Symptomatic | ||||||
| Sampling date | 2.3 | 17,159 | 0.004 | 1.1 | 16,150 | 0.362 |
| Plant parts | 18.5 | 17,159 | <0.001 | 16.5 | 2150 | <0.001 |
| Sampling date × plant parts | 0.6 | 34,159 | 0.941 | 0.9 | 32,150 | 0.567 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monterrosa, A.; Paret, M.L.; Ochoa, R.; Ulsamer, A.; Joseph, S.V. Temporal Incidence of Eriophyid Mites on Rose Rosette Disease-Symptomatic and -Asymptomatic Roses in Central Georgia, USA. Pathogens 2022, 11, 228. https://doi.org/10.3390/pathogens11020228
Monterrosa A, Paret ML, Ochoa R, Ulsamer A, Joseph SV. Temporal Incidence of Eriophyid Mites on Rose Rosette Disease-Symptomatic and -Asymptomatic Roses in Central Georgia, USA. Pathogens. 2022; 11(2):228. https://doi.org/10.3390/pathogens11020228
Chicago/Turabian StyleMonterrosa, Alejandra, Mathews L. Paret, Ronald Ochoa, Andrew Ulsamer, and Shimat V. Joseph. 2022. "Temporal Incidence of Eriophyid Mites on Rose Rosette Disease-Symptomatic and -Asymptomatic Roses in Central Georgia, USA" Pathogens 11, no. 2: 228. https://doi.org/10.3390/pathogens11020228