
An Overview of Anthropogenic Actions as Drivers for Emerging and Re-Emerging Zoonotic Diseases
 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Anthropogenic Actions
2.1. Climate Change
2.2. Deforestation
2.3. Trafficking and Consumption of Wild Animals
3. Human–Host–Environment Interaction
4. Selected Emerging and Re-Emerging Viral Pathogens
4.1. Ebola Virus
4.2. Hantaviruses
4.3. Hendravirus
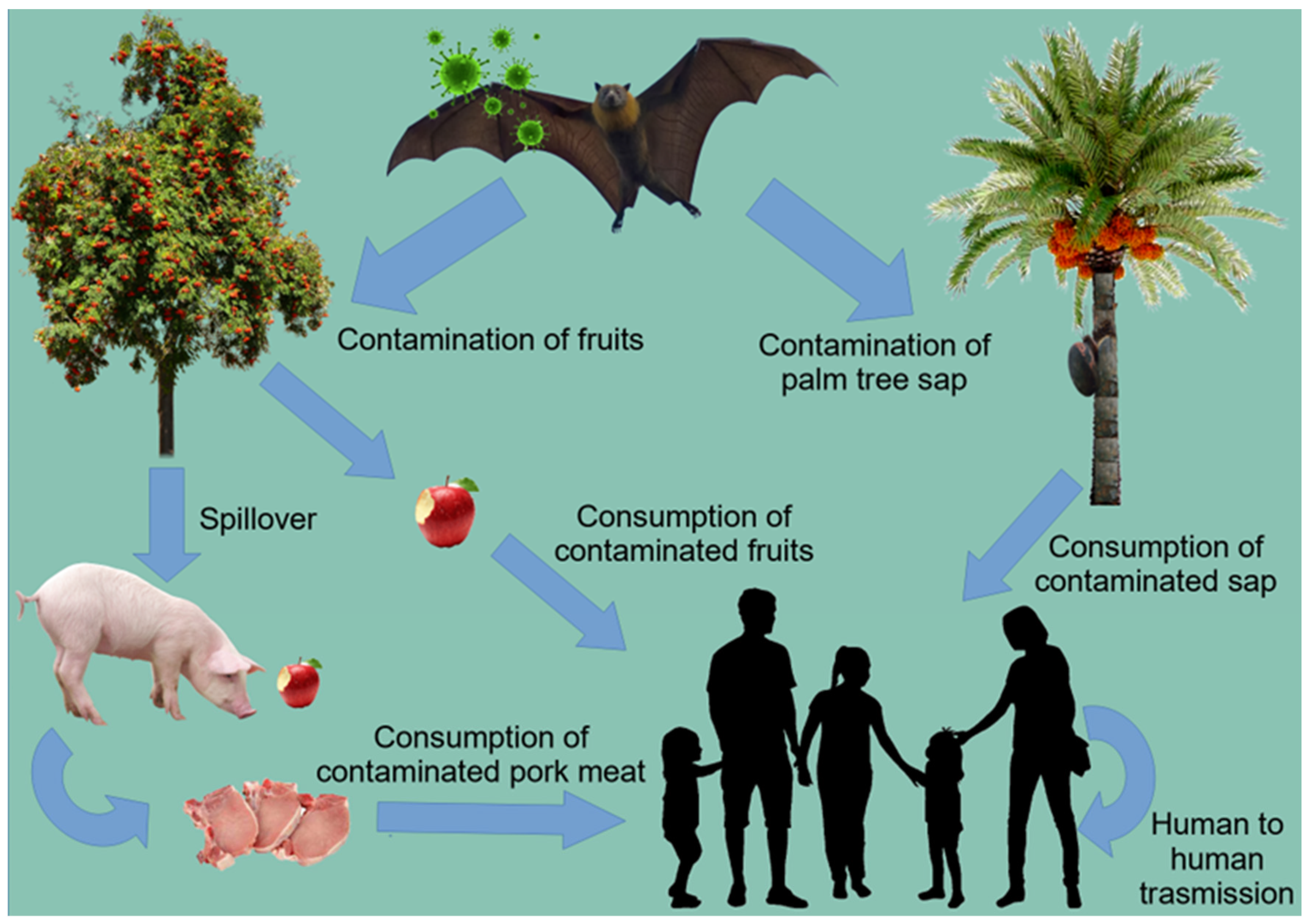
4.4. Nipah Virus
4.5. Rabies
4.5.1. Rabies and Urbanization
4.5.2. Rabies and Garbage Accumulation
4.5.3. Rabies and Pets
4.5.4. Rabies and Deforestation
4.5.5. Rabies and Food Supply
4.6. SARS-CoV-2
5. Selected Bacterial Diseases
5.1. Leptospirosis
5.1.1. Leptospirosis and Urbanization
5.1.2. Leptospirosis and Extreme Weather Events
5.1.3. Leptospirosis and Socio-Economic Phenomena
5.1.4. Leptospirosis and Pets
5.1.5. Leptospirosis and Wildlife Animals
5.2. Lyme-Borreliosis
5.2.1. Borrelia spp. and Geographical Distribution
5.2.2. Impacts of Climatic Changes and Anthropogenic Activities on Lyme Disease
5.3. Tuberculosis
6. Selected Parasitic Diseases
Leishmaniasis
7. Fungal Diseases
8. Recommendations
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Acha, P.N.; Szyfres, B. Preface of the first English edition. In Zoonoses and Communicable Diseases Common to Man and Animals, 3rd ed.; OIE: Paris, France, 2005; Volume 1, p. ix. [Google Scholar]
- Cutler, S.J.; Fooks, A.R.; van der Poel, W.H.M. Public health threat of new, reemerging, and neglected zoonoses in the industrialized world. Emerg. Infect. Dis. 2010, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.; Kamel, M. Climatic changes and their role in emergence and re-emergence of diseases. Environ. Sci. Pollut. Res. 2020, 27, 22336–22352. [Google Scholar] [CrossRef] [PubMed]
- WHO. Anticipating Emerging Infectious Disease Epidemics; Meeting Report; WHO: Geneva, Switzerland, 2015; Available online: https://apps.who.int/iris/bitstream/handle/10665/252646/WHO-OHE-PED-2016.2-eng.pdf (accessed on 1 August 2022).
- Adhikari, S.P.; Meng, S.; Wu, Y.; Mao, Y.; Ye, R.; Wang, Q.; Sun, C.; Sylvia, S.; Rozelle, S.; Raat, H.; et al. Epidemiology, causes, clinical manifestation and diagnosis, prevention, and control of coronavirus disease (COVID-19) during the early outbreak period: A scoping review. Infect. Dis. Poverty 2020, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morand, S. Emerging diseases, livestock expansion and biodiversity loss are positively related at global scale. Biol. Conserv. J. 2020, 248, 108707. [Google Scholar] [CrossRef] [PubMed]
- Sironi, M.; Hasnain, S.E.; Rosenthal, B.; Phan, T.; Luciani, F. SARS-CoV-2 and COVID-19: A genetic, epidemiological, and evolutionary perspective. Infect. Genet. Evol. 2020, 84, 104384. [Google Scholar] [CrossRef]
- Beyer, R.M.; Manica, A.; Mora, C. Shifts in global bat diversity suggest a possible role of climate change in the emergence of SARS-CoV-1 and SARS-CoV-2. Sci. Total Environ. 2021, 767, 145413. [Google Scholar] [CrossRef]
- Halonen, J.I.; Erhola, M.; Furman, E.; Haahtela, T.; Jousilahti, P.; Barouki, R.; Bergman, A.; Billo, N.E.; Fuller, R.; Haines, A.; et al. A call for urgent action to safeguard our planet and our health in line with the Helsinki declaration. Environ. Res. 2021, 193, 110600. [Google Scholar] [CrossRef]
- Morand, S.; Lajaunie, C. Outbreaks of vector-borne and zoonotic diseases are associated with changes in Forest Cover and Oil Palm expansion at global scale. Front. Vet. Sci. 2021, 8, 1–11. [Google Scholar] [CrossRef]
- Bowler, D.E.; Bjorkman, A.D.; Dornelas, M.; Myers-Smith, I.H.; Navarro, L.M.; Niamir, A.; Supp, S.R.; Waldock, C.; Winter, M.; Vellend, M.; et al. Mapping human pressures on biodiversity across the planet uncovers anthropogenic threat complexes. People Nat. 2020, 2, 380–394. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014 Synthesis Report—Summary Chapter for Policymakers. 2014. Available online: https://www.ipcc.ch/site/assets/uploads/2018/02/AR5_SYR_FINAL_SPM.pdf (accessed on 1 August 2022).
- Farber, D.A.; Carlarne, C.P. Climate Change Law; Law Work Paper; West Academic Publishing: St. Paul, MN, USA, 2017; p. 419. [Google Scholar]
- IPBES. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Díaz, S., Settele, J., Brondízio, E.S., Ngo, H.T., Guèze, M., Agard, J., Arneth, A., Balvanera, P., Brauman, K.A., Butchart, M.S.H., et al., Eds.; IPBES Secretariat: Bonn, Germany, 2019; 56p, Available online: https://ipbes.net/sites/default/files/inline/files/ipbes_global_assessment_report_summary_for_policymakers.pdf (accessed on 20 January 2022).
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Gregg, J.W.; Lenton, T.M.; Palomo, I.; Eikelboom, J.A.J.; Law, B.E.; Huq, S.; Duffy, P.B.; et al. World scientists’ warning of a climate emergency 2021. Bioscience 2021, 71, 894–898. [Google Scholar] [CrossRef]
- Collins, A.; Galli, A.; Hipwood, T.; Murthy, A. Living within a one planet reality: The contribution of personal footprint calculators. Environ. Res. Lett. 2020, 15, 025008. [Google Scholar] [CrossRef]
- Volpato, G.; Fontefrancesco, M.F.; Gruppuso, P.; Zocchi, D.M.; Pieroni, A. Baby pangolins on my plate: Possible lessons to learn from the COVID-19 pandemic. J. Ethnobiol. Ethnomed. 2020, 16, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulet, C. A Multi-Scale Evaluation of Eastern Hognose Snake (Heterodon platirhinos) Habitat Selection at the Northern Extent of Its Range. Master’s Thesis, University of New Hampshire, Durham, NH, USA, 2010. Available online: https://scholars.unh.edu/cgi/viewcontent.cgi?article=1546&context=thesis (accessed on 20 January 2022).
- Patz, J.; Githeko, A.; McCarty, J.; Hussain, S.; Confalonieri, U.; de Wet, N. Climate change and infectious diseases. In Climate Change and Human Health: Risks and Responses; World Health Organization: Geneva, Switzerland, 2003; pp. 103–110. [Google Scholar]
- Gorris, M.E. Environmental Infectious Disease Dynamics in Relation to Climate and Climate Change. Ph.D. Thesis, University of California, Irvine, CA, USA, 2019. [Google Scholar]
- Mogrovejo-Arias, D.C.; Brill, F.H.H.; Wagner, D. Potentially pathogenic bacteria isolated from diverse habitats in Spitsbergen, Svalbard. Environ. Earth Sci. 2020, 79, 109. [Google Scholar] [CrossRef] [Green Version]
- Panda, A.K.; Thakur, S.D.; Katoch, R.C. Rabies: Control strategies for Himalayan states of the Indian subcontinent. J. Commun. Dis. 2008, 40, 169–175. [Google Scholar] [PubMed]
- Wu, T.; Perrings, C.; Kinzig, A.; Collins, J.P.; Minteer, B.A.; Daszak, P. Economic growth, urbanization, globalization, and the risks of emerging infectious diseases in China: A review. Ambio 2017, 46, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Bloomfield, L.S.P.; McIntosh, T.L.; Lambin, E. Habitat fragmentation, livelihood behaviors, and contact between people and nonhuman primates in Africa. Landsc. Ecol 2020, 35, 985–1000. [Google Scholar] [CrossRef]
- Van der Werf, G.R.; Morton, D.C.; DeFries, R.S.; Giglio, L.; Randerson, J.T.; Collatz, G.J.; Kasibhatla, P.S. Estimates of fire emissions from an active deforestation region in the southern Amazon based on satellite data and biogeochemical modelling. Biogeosciences 2009, 6, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Americo, T. Dia da Amazônia: Desmatamento da Floresta É 8,2 % Maior em 2021. CNN Brasil. 2021. Available online: https://www.cnnbrasil.com.br/nacional/dia-da-amazonia-desmatamento-da-floresta-e-82-maior-em-2021/ (accessed on 20 January 2022).
- Chaves, L.S.M.; Conn, J.E.; López, R.V.M.; Sallum, M.A.M. Abundance of impacted forest patches less than 5 km is a key driver of the incidence of malaria in Amazonian Brazil. Sci. Rep. 2018, 8, 7077. [Google Scholar] [CrossRef] [Green Version]
- Shuman, E.K. Global climate change and infectious diseases. N. Engl. J. Med. 2010, 362, 1061–1063. [Google Scholar] [CrossRef]
- Nagy, L.; Forsberg, B.R.; Artaxo, P. Interactions between biosphere, atmosphere and human land use in the Amazon Basin analysis and synthesis. In Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2016; Volume 227, ISBN 978-3-662-49900-9. [Google Scholar]
- Wilkinson, D.A.; Marshall, J.C.; French, N.P.; Hayman, D.T.S. Habitat fragmentation, biodiversity loss and the risk of novel infectious disease emergence. J. R. Soc. Interface 2018, 15, 20180403. [Google Scholar] [CrossRef]
- Mackenstedt, U.; Jenkins, D.; Romig, T. The role of wildlife in the transmission of parasitic zoonoses in peri-urban and urban areas. Int. J. Parasitol. Parasites Wildl. 2015, 4, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellwanger, J.H.; Kulmann-Leal, B.; Kaminski, V.L.; Valverde-Villegas, J.M.; DA VEIGA, A.B.G.; Spilki, F.R.; Fearnside, P.M.; Caesar, L.; Giatti, L.L.; Wallau, G.L.; et al. Beyond diversity loss and climate change: Impacts of Amazon deforestation on infectious diseases and public health. An. Acad. Bras. Cienc. 2020, 92, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Chomel, B.B.; Belotto, A.; Meslin, F.X. Wildlife, exotic pets, and emerging zoonoses. Emerg. Infect. Dis. 2007, 13, 6–11. [Google Scholar] [CrossRef] [PubMed]
- CONAB. Boletim da Safra de Grãos. 2021. Available online: https://www.conab.gov.br/info-agro/safras/graos/boletim-da-safra-de-graos (accessed on 20 January 2022).
- Madeiro, C. Pecuária Responde por 75% do Desmatamento em Terras Públicas da Amazônia. UOL. 2021. Available online: https://noticias.uol.com.br/meio-ambiente/ultimas-noticias/redacao/2021/10/27/amazonia-87-do-desmate-em-terras-publicas-ocorreu-em-areas-nao-destinadas.htm (accessed on 20 January 2022).
- UOL. WWF: Aumento do Desmatamento no Brasil Piora Risco de Zoonoses e COVID. UOL. 2021. Available online: https://noticias.uol.com.br/meio-ambiente/ultimas-noticias/redacao/2021/01/12/estudo-wwf-desmatamento-brasil-amazonia.htm (accessed on 10 January 2022).
- Rattis, L.; Brando, P.M.; Macedo, M. Climatic limit for agriculture in Brazil. Nat. Clim. Chang. 2021, 11, 1098–1104. [Google Scholar] [CrossRef]
- Rahimi, P.; Islam, M.S.; Duarte, P.M.; Tazerji, S.S.; Sobur, M.A.; El Zowalaty, M.E.; Ashour, H.M.; Rahman, M.T. Impact of the COVID-19 pandemic on food production and animal health. Trends Food Sci. Technol. 2022, 121, 105–113. [Google Scholar] [CrossRef]
- Rahman, M.T.; Islam, M.S.; Shehata, A.A.; Basiouni, S.; Hafez, H.M.; Azhar, E.I.; Khafaga, A.F.; Bovera, F.; Attia, Y.A. Influence of COVID-19 on the sustainability of livestock performance and welfare on a global scale. Trop. Anim. Health Prod. 2022, 54, 309. [Google Scholar] [CrossRef]
- Attia, Y.A.; Rahman, M.T.; Shehata, A.A.; Hafez, H.M.; Hossain, M.J.; Basiouni, S.; Khafaga, A.F. Poultry production and sustainability in developing countries under the COVID-19 crisis: Lessons learned. Animals 2022, 12, 644. [Google Scholar] [CrossRef]
- United Nations. More than Half a Billion Pushed into Extreme Poverty Due to Health Costs. 2021. Available online: https://news.un.org/en/story/2021/12/110775 (accessed on 20 January 2022).
- FAO Commission on Genetic Resources for Food and Agriculture Assessments. The State of the World’s Biodiversity for Food and Agriculture; Bélanger, J., Pilling, D., Eds.; FAO: Rome, Italy, 2019; 57p, Available online: http://www.fao.org/3/CA3129EN/CA3129EN.pdf (accessed on 20 September 2022).
- Ribeiro, V.M.F.; De Carvalho, Y.K.; Peruquetti, R.C.; Medeiros, L.S.; Freitas, H.J. De Consumo e Comercialização de Carnes Silvestres: Potencial econômico para a Amazônia Ocidental. J. Amaz. Health Sci. 2016, 2, 1–11. [Google Scholar]
- Cajaiba, R.L.; Da Silva, W.B.; Piovesan, P.R.R. Animais silvestres utilizados como recurso alimentar em assentamentos rurais no município de Uruará, Pará, Brasil. Desenvolv. Meio Ambient. 2015, 34. [Google Scholar] [CrossRef]
- De Souza, L.S.; Progênio, M.; de Souza, L.S.; de Araújo Santos, F.G. Consumption of wild animals in extractive communities in the State of Acre, Brazilian Amazon. Biota Amaz. 2021, 11, 27–30. [Google Scholar]
- Jerozolimski, A.; Peres, C.A. Bringing home the biggest bacon: A cross-site analysis of the structure of hunter-kill profiles in Neotropical forests. Biol. Conserv. 2003, 111, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Dias, C.J.; Almeida, M.W.B. A floresta como mercado: Caça e conflito na Reserva Extrativista do Alto Juruá (AC). Bol. Rede Amaz. 2004, 3, 9–27. [Google Scholar]
- Chow, A.T.; Cheung, S.; Yip, P.K. Wildlife markets in south China. Hum. -Wildl. Interact. 2014, 8, 108–112. [Google Scholar]
- Contini, C.; Di Nuzzo, M.; Barp, N.; Bonazza, A.; de Giorgio, R.; Tognon, M.; Rubino, S. The novel zoonotic COVID-19 pandemic: An expected global health concern. J. Infect. Dev. Ctries. 2020, 14, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.M.; Li, R.; Ling, J.; Grace, D.; Nguyen-Viet, H.; Lindahl, J.F. Live and wet markets: Food Access versus the risk of disease emergence. Trends Microbiol. 2021, 29, 573–581. [Google Scholar] [CrossRef]
- Thomas, J.; Balseiro, A.; Gortázar, C.; Risalde, M.A. Diagnosis of tuberculosis in wildlife: A systematic review. Vet. Res. 2021, 52, 31. [Google Scholar] [CrossRef]
- Goosen, W.; Moseley, M.H.; Kerr, T.J.; Potts, A.; Miller, M. The seroepidemiology of a neglected zoonotic and livestock pathogen in free-ranging bovids: Leptospirosis in African buffaloes (Syncerus caffer). Pathogens 2021, 10, 1072. [Google Scholar] [CrossRef]
- Pinto, C.C.; Amin da Silva, B.L.; Santos, E.S.A.; Oliveira, S.R.M.; Amorim, M.T.; Amaro, O.; Gomes, E.P.A.; Casseb, S.M.M. Perfil Epidemiológico da Raiva Humana na Região Norte do Estado do Pará durante o período de 2000 a 2019. Saúde Coletiva 2021, 11, 6937–6948. [Google Scholar] [CrossRef]
- Godfroid, J.; Al Dahouk, S.; Pappas, G.; Roth, F.; Matope, G.; Muma, J.; Marcotty, T.; Pfeiffer, D.; Skjerve, E. A “One Health” surveillance and control of brucellosis in developing countries: Moving away from improvisation. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Formenti, N.; Calò, S.; Parisio, G.; Guarneri, F.; Birbes, L.; Pitozzi, A.; Scali, F.; Tonni, M.; Guadagno, F.; Giovannini, S.; et al. ESBL/AmpC-producing Escherichia coli in wild boar: Epidemiology and risk factors. Animals 2021, 11, 1855. [Google Scholar] [CrossRef]
- Zoological Society of London. Illegal wildlife Trade Impacts. 2022. Available online: https://www.zsl.org/conservation/how-we-work/illegal-wildlife-trade-crisis/illegal-wildlife-trade-impacts (accessed on 20 January 2022).
- Koh, L.P.; Li, Y.; Lee, J.S.H. The value of China’s ban on wildlife trade and consumption. Nat. Sustain. 2021, 4, 2–4. [Google Scholar] [CrossRef]
- Zhang, L.; Hua, N.; Sun, S. Wildlife trade, consumption, and conservation awareness in southwest China. Biodivers. Conserv. 2008, 17, 1493–1516. [Google Scholar] [CrossRef] [PubMed]
- Bernardo-Cravo, A.P.; Schmeller, D.S.; Chatzinotas, A.; Vredenburg, V.T.; Loyau, A. Environmental factors and host microbiomes shape host–pathogen dynamics. Trends Parasitol. 2020, 36, 616–633. [Google Scholar] [CrossRef] [PubMed]
- Despommier, D.; Ellis, B.R.; Wilcox, B.A. The role of ecotones in emerging infectious diseases. Ecohealth 2006, 3, 281–289. [Google Scholar] [CrossRef]
- Destoumieux-Garzón, D.; Mavingui, P.; Boetsch, G.; Boissier, J.; Darriet, F.; Duboz, P.; Fritsch, C.; Giraudoux, P.; Le Roux, F.; Morand, S.; et al. The one health concept: 10 years old and a long road ahead. Front. Vet. Sci. 2018, 5, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marco, M.; Baker, M.L.; Daszak, P.; de Barro, P.; Eskew, E.A.; Godde, C.M.; Harwood, T.D.; Herrero, M.; Hoskins, A.J.; Johnson, E.; et al. Sustainable development must account for pandemic risk. Proc. Natl. Acad. Sci. USA 2020, 117, 3888–3892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, K.A.; Daszak, P. Human ecology in pathogenic landscapes: Two hypotheses on how. Curr. Opin. Virol. 2013, 3, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change, and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Patz, J.A.; Daszak, P.; Tabor, G.M.; Aguirre, A.A.; Pearl, M.C.; Epstein, J.; Wolfe, N.D.; Kilpatrick, A.M.; Foufopoulos, J.; Molyneux, D.; et al. Unhealthy landscapes: Policy recommendations on land use change and infectious disease emergence. Environ. Health Perspect. 2004, 112, 1092–1098. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, N.D.; Dunavan, C.P.; Diamond, J. Origins of major human infectious diseases. Nature 2007, 447, 279–283. [Google Scholar] [CrossRef]
- Dimakakou, E.; Johnston, H.J.; Streftaris, G.; Cherrie, J.W. Exposure to environmental and occupational particulate air pollution as a potential contributor to neurodegeneration and diabetes: A systematic review of epidemiological research. Int. J. Environ. Res. Public Health 2018, 15, 1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roca, M.; Manes, F.; Chade, A.; Gleichgerrcht, E.; Gershanik, O.; Arévalo, G.G.; Torralva, T.; Duncan, J. The relationship between executive functions and fluid intelligence in Parkinson’s disease. Psychol. Med. 2012, 42, 2445–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, C.E.; Ma, J.; Gaudet, M.M.; Newman, L.A.; Miller, K.D.; Goding Sauer, A.; Jemal, A.; Siegel, R.L. Breast cancer statistics, 2019. CA. Cancer J. Clin. 2019, 69, 438–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocco, P.; Blair, A.; Congia, P.; Saba, G.; Ecca, A.R.; Palmas, C. Long-term health effects of the occupational exposure to DDT. A preliminary report. Ann. N. Y. Acad. Sci. 1997, 837, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Krüger, M.; Basiouni, S.; Eder, I.; Rodloff, A. Susceptibility of extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae to Roundup. Ger. J. Microbiol. 2021, 1, 7–15. [Google Scholar] [CrossRef]
- Kaonga, N.; Hang’ombe, B.M.; Lupindu, A.M.; Hoza, A.S. Detection of CTX-M-type extended-spectrum beta-lactamase producing Salmonella Typhimurium in commercial poultry farms in Copperbelt Province, Zambia. Ger. J. Vet. Res. 2021, 1, 27–34. [Google Scholar] [CrossRef]
- Shehata, A.A.; Basiouni, S.; Abd Elrazek, A.; Sultan, H.; Tarabees, R.; Elsayed, M.S.A.; Talat, S.; Moharam, E.; Said, A.; Mohsen, W.A.; et al. Characterization of Salmonella enterica isolated from poultry hatcheries and commercial broiler chickens. Pak. Vet. J. 2019, 39, 515–520. [Google Scholar] [CrossRef]
- Benrabia, I.; Hamdi, T.M.; Shehata, A.A.; Neubauer, H.; Wareth, G. Methicillin-resistant Staphylococcus aureus (MRSA) in poultry species in Algeria: Long-term study on prevalence and antimicrobial resistance. Vet. Sci. 2020, 7, 54. [Google Scholar] [CrossRef]
- Njeru, J. Emerging carbapenem resistance in ESKAPE pathogens in Sub-Saharan Africa and the way forward. Ger. J. Microbiol. 2021, 1, 3–6. [Google Scholar] [CrossRef]
- Bogitsh, B.J.; Carter, C.E.; Oeltmann, T.N. Human Parasitology, 3rd ed.; Academic Press: New York, NY, USA, 2005. [Google Scholar]
- Baseler, L.; Chertow, D.S.; Johnson, K.M.; Feldmann, H.; Morens, D.M. The pathogenesis of Ebola virus disease. Annu. Rev. Pathol. 2017, 12, 387–418. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Bào, Y.; Bavari, S.; Becker, S.; Bradfute, S.; Brauburger, K.; Rodney Brister, J.; Bukreyev, A.A.; Caì, Y.; Chandran, K.; et al. Virus nomenclature below the species level: A standardized nomenclature for filovirus strains and variants rescued from cDNA. Arch. Virol. 2014, 159, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.; Bollinger, L.; Johnson, J.C.; Wada, J.; Radoshitzky, S.R.; Palacios, G.; Bavari, S.; Jahrling, P.B.; Kuhn, J.H. Neglected filoviruses. FEMS Microbiol. Rev. 2016, 40, 494–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, T.; Anthony, S.J.; Gbakima, A.; Bird, B.H.; Bangura, J.; Tremeau-Bravard, A.; Belaganahalli, M.N.; Wells, H.L.; Dhanota, J.K.; Liang, E.; et al. The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat. Microbiol. 2018, 3, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Forbes, K.M.; Webala, P.W.; Jääskeläinen, A.J.; Abdurahman, S.; Ogola, J.; Masika, M.M.; Kivistö, I.; Alburkat, H.; Plyusnin, I.; Levanov, L.; et al. Bombali Virus in Mops condylurus Bat, Kenya Kristian. Emerg. Infect. Dis. 2019, 25, 955–957. [Google Scholar] [CrossRef]
- Olivero, J.; Fa, J.E.; Farfán, M.; Márquez, A.L.; Real, R.; Juste, F.J.; Leendertz, S.A.; Nasi, R. Human activities link fruit bat presence to Ebola virus disease outbreaks. Mamm. Rev. 2020, 50, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Avšič-Županc, T.; Saksida, A.; Korva, M. Hantavirus infections. Clin. Microbiol. Infect. 2019, 21, e6–e16. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, J.G.; Blunsum, A.E.; Carmichael, S.; Smollett, K.; Maxwell-Scott, H.; Farmer, E.C.W.; Osborne, J.; MacLean, A.; Ashraf, S.; Shah, R.; et al. Seoul virus associated with pet rats, scotland, uk, 2019. Emerg. Infect. Dis. 2021, 27, 2677–2980. [Google Scholar] [CrossRef]
- Clement, J.; LeDuc, J.W.; Lloyd, G.; Reynes, J.M.; Reynes, J.; McElhinney, L.; Lee, H.W. Wild rats, laboratory rats, pet rats: Global seoul hantavirus disease revisited. Viruses 2019, 11, 652. [Google Scholar] [CrossRef] [Green Version]
- Adesiyun, A.; Dookeran, S.; Stewart-Johnson, A.; Rahaman, S.; Bissessar, S.; Thompson, N. Serological evidence of hantavirus infection in farm and abattoir workers in Trinidad—A preliminary study. J. Agromed. 2011, 16, 194–199. [Google Scholar] [CrossRef]
- Friesema, I.H.M.; Bakker, J.; Maas, M.; Goris, M.G.A.; van der Giessen, J.W.B.; Rockx, B.H.G. Seroprevalence of hantaviruses and Leptospira in muskrat and coypu trappers in The Netherlands, 2016. Infect. Ecol. Epidemiol. 2018, 8, 1–4. [Google Scholar]
- Akar, N.; Çalişkan, E.; Öztürk, C.E.; Ankarali, H.; Kilinçel, Ö.; Öksüz, Ş.; Şahin, İ. Seroprevalence of hantavirus and Borrelia burgdorferi in Düzce (Turkey) forest villages and the relationship with sociodemographic features. Turk. J. Med. Sci. 2019, 49, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, P.; Tia, M.; Alabi, A.; Anon, J.C.; Auste, B.; Essbauer, S.; Gnionsahe, A.; Kigninlman, H.; Klempa, B.; Kraef, C.; et al. Human infections by non-rodent-associated hantaviruses in Africa. J. Infect. Dis. 2016, 214, 1507–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, J.F.; Jonsson, C.B.; Esteban, E.; Galligan, D.; Basombrio, M.A.; Peralta-Ramos, M.; Bharadwaj, M.; Torrez-Martinez, N.; Callahan, J.; Segovia, A.; et al. High prevalence of hantavirus infection in Indian communities of the Paraguayan and Argentinean Gran Chaco. Am. J. Trop. Med. Hyg. 1998, 59, 438–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, M.T.; Vial, P.C.; Castillo, C.H.; Godoy, P.M.; Hjelle, B.; Ferrés, M.G. Hantavirus prevalence in the IX Region of Chile. Emerg. Infect. Dis. 2003, 9, 827–832. [Google Scholar]
- Gonzalez, L.M.; Lindsey, A.E.; Hjelle, B.; Dominguez, D.; Brown, J.; Goade, D.; Jonsson, C.B. Prevalence of antibodies to Sin Nombre virus in humans living in rural areas of southern New Mexico and western Texas. Virus Res. 2001, 74, 177–179. [Google Scholar] [CrossRef]
- Guzmán, C.; Mattar, S.; Levis, S.; Pini, N.; Figueiredo, T.; Mills, J.; Salazar-bravo, J. Prevalence of antibody to hantaviruses in humans and rodents in the Caribbean region of Colombia determined using Araraquara and Maciel virus antigens Camilo. Mem. Instit. Oswaldo Cruz 2013, 108, 167–171. [Google Scholar] [CrossRef]
- Jääskeläinen, A.J.; Voutilainen, L.; Lehmusto, R.; Henttonen, H.; Lappalainen, M.; Kallio-Kokko, H.; Vaheri, A.; Vapalahti, O. Serological survey in the Finnish human population implies human-to-human transmission of Ljungan virus or antigenically related viruses. Epidemiol. Infect. 2016, 144, 1278–1285. [Google Scholar] [CrossRef] [Green Version]
- Hukic, M.; Nikolic, J.; Valjevac, A.; Seremet, M.; Tesic, G.; Markotic, A. A serosurvey reveals Bosnia and Herzegovina as a Europe’s hotspot in hantavirus seroprevalence. Epidemiol. Infect. 2010, 138, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Khan, M.; Ullah, S.; Wei, D.Q. Hantavirus: The next pandemic we are waiting for? Interdiscip. Sci. Comput. Life Sci. 2021, 13, 147–152. [Google Scholar] [CrossRef]
- Zeimes, C.B.; Quoilin, S.; Henttonen, H.; Lyytikäinen, O.; Vapalahti, O.; Reynes, J.M.; Reusken, C.; Swart, A.N.; Vainio, K.; Hjertqvist, M.; et al. Landscape and regional environmental analysis of the spatial distribution of hantavirus human cases in Europe. Front. Public Health 2015, 3, 54. [Google Scholar] [CrossRef] [Green Version]
- Chua, K.B.; Bellini, W.J.; Rota, P.A.; Harcourt, B.H.; Tamin, A.; Lam, S.K.; Ksiazek, T.G.; Rollin, P.E.; Zaki, S.R.; Shieh, W.; et al. Nipah virus: A recently emergent deadly paramyxovirus. Science 2000, 288, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.E.; Hero, J.M. Rainforest frogs of the Australian Wet Tropics: Guild classification and the ecological similarity of declining species. Proc. R. Soc. B Biol. Sci. 1998, 265, 597–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ksiazek, T.G.; Rota, P.A.; Rollin, P.E. A review of Nipah and Hendra viruses with an historical aside. Virus Res. 2011, 162, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Harcourt, B.H.; Tamin, A.; Ksiazek, T.G.; Rollin, P.E.; Anderson, L.J.; Bellini, W.J.; Rota, P.A. Molecular characterization of Nipah virus, a newly emergent Paramyxovirus. Virology 2000, 349, 334–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamin, A.; Harcourt, B.H.; Ksiazek, T.G.; Rollin, P.E.; Bellini, W.J.; Rota, P.A. Functional properties of the fusion, and attachment glycoproteins of Nipah virus. Virology 2002, 200, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Negrete, O.A.; Levroney, E.L.; Aguilar, H.C.; Bertolotti-Ciarlet, A.; Nazarian, R.; Tajyar, S.; Lee, B. EphrinB2 the entry receptor for Nipah virus, an emergent deadly paramyxovirus. Nature 2005, 436, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Halpin, K.; Young, P.L.; Field, H.E.; Mackenzie, J.S. Isolation of Hendra virus from pteropid bats: A natural reservoir of Hendra virus. J. Gen. Virol. 2000, 81, 1927–1932. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Lek Koh, C.; Hooi, P.S.; Wee, K.F.; Khong, J.H.; Chua, B.H.; Chan, Y.P.; Lim, M.E.; Lam, S.K. Isolation of Nipah virus from Malaysian Island flying-foxes. Microbes Infect. 2002, 4, 145–151. [Google Scholar] [CrossRef]
- Luby, S.P.; Gurley, E.S.; Hossain, M.J. Transmission of human infection with Nipah virus. Clin. Infect. Dis. 2009, 49, 1743–1748. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Dhama, K.; Chakraborty, S.; Tiwari, R.; Natesan, S.; Khandia, R.; Munjal, A.; Vora, K.S.; Latheef, S.K.; Karthik, K.; et al. Nipah virus: Epidemiology, pathology, immunobiology and advances in diagnosis, vaccine designing and control strategies–a comprehensive review. Vet. Q. 2019, 39, 26–55. [Google Scholar] [CrossRef] [Green Version]
- Pillai, V.S.; Krishna, G.; Veettil, M.V. Nipah virus: Past outbreaks and future containment. Viruses 2020, 12, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, M.S.H. The implications of population growth and climate change on sustainable development in Bangladesh. Jamba J. Disaster Risk Stud. 2019, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, B.; Dhimal, M.; Latheef, A.T.; Ghosh, U. Climate change: Health effects and response in South Asia. BMJ 2017, 359, j5117. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B. Risk factors, prevention, and communication strategy during Nipah virus outbreak in Malaysia. Malays. J. Pathol. 2010, 32, 75–80. [Google Scholar] [PubMed]
- Chua, K.B.; Chua, B.H.; Wang, C.W. Anthropogenic deforestation, El Niño, and the emergence of Nipah virus in Malaysia. Malays. J. Pathol. 2002, 24, 15–21. [Google Scholar] [PubMed]
- Field, H.; Young, P.; Yob, J.M.; Mills, J.; Hall, L.; Mackenzie, J. The natural history of Hendra and Nipah viruses. Microbes Infect. 2001, 3, 307–314. [Google Scholar] [CrossRef]
- Plowright, R.K.; Foley, P.; Field, H.E.; Dobson, A.P.; Foley, J.E.; Eby, P.; Daszak, P. Urban habituation, ecological connectivity, and epidemic dampening: The emergence of hendra virus from flying foxes (Pteropus spp.). Proc. R. Soc. B Biol. Sci. 2011, 278, 3703–3712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Hossain, M.J.; Sultana, S.; Homaira, N.; Khan, S.U.; Rahman, M.; Gurley, E.S.; Rollin, P.E.; Lo, M.K.; Comer, J.A.; et al. Date palm sap linked to Nipah virus outbreak in Bangladesh, 2008. Vector-Borne Zoonotic Dis. 2012, 12, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Nahar, N.; Asaduzzaman, M.; Mandal, U.K.; Rimi, N.A.; Gurley, E.S.; Rahman, M.; Garcia, F.; Zimicki, S.; Sultana, R.; Luby, S.P. Hunting Bats for Human Consumption in Bangladesh. Ecohealth 2020, 17, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Ang, B.S.P.; Lim, T.C.C.; Wang, L. Nipah virus infection. J. Clin. Microbiol. 2018, 56, e01875-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurley, E.S.; Hegde, S.T.; Hossain, K.; Sazzad, H.M.S.; Hossain, M.J.; Rahman, M.; Yushuf Sharker, M.A.; Salje, H.; Islam, M.S.; Epstein, J.H.; et al. Convergence of humans, bats, trees, and culture in Nipah virus transmission, Bangladesh. Emerg. Infect. Dis. 2017, 23, 1446–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantha, S.; Subedi, D.; Poudel, U.; Subedi, S.; Kaphle, K.; Dhakal, S. Review of rabies in Nepal. One Health 2020, 10, 100155. [Google Scholar] [CrossRef] [PubMed]
- Rohde, R.E.; Rupprecht, C.E. Update on lyssaviruses and rabies: Will past progress play as prologue in the near term towards future elimination? Fac. Rev. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.; Subedi, D.; Pantha, S.; Prasad Acharya, K.; Hendrik Nel, L. The role of waste management in control of rabies: A neglected issue. Viruses 2021, 13, 225. [Google Scholar] [CrossRef]
- Singh, R.; Singh, K.P.; Cherian, S.; Saminathan, M.; Kapoor, S.; Reddy, G.B.M.; Panda, S.; Dhama, K. Rabies—Epidemiology, pathogenesis, public health concerns and advances in diagnosis and control: A comprehensive review. Vet. Q. 2017, 37, 212–251. [Google Scholar] [CrossRef] [Green Version]
- Ghasemzadeh, I.; Namazi, S.H. Review of bacterial and viral zoonotic infections transmitted by dogs. J. Med. Life 2015, 8, 1–5. [Google Scholar]
- World Health Organization. WHO Expert Consultation on Rabies: Third Report. 2018. Available online: https://apps.who.int/iris/handle/10665/272364 (accessed on 5 September 2022).
- Deshmukh, D.G.; Damle, A.S.; Bajaj, J.K.; Bhakre, J.B.; Patil, N.S. Fatal rabies despite post-exposure prophylaxis. Indian J. Med. Microbiol. 2011, 29, 178–180. [Google Scholar] [CrossRef]
- Ngoepe, C.E.; Sabeta, C.; Nel, L. The spread of canine rabies into Free State province of South Africa: A molecular epidemiological characterization. Virus Res. 2009, 142, 175–180. [Google Scholar] [CrossRef]
- WHO. Frequently Asked Questions about Rabies for the General Public. 2018. Available online: https//www who int/rabies/Rabies_General_Public_FAQs_20Sep2018 pdf (accessed on 20 September 2022).
- Jackson, A.C. Rabies: A medical perspective. Rev. Sci. Tech. 2018, 37, 569–580. [Google Scholar] [CrossRef]
- Rahman, M.T.; Sobur, M.A.; Islam, M.S.; Ievy, S.; Hossain, M.J.; Zowalaty, M.E.E.; Rahman, A.M.M.T.; Ashour, H.M. Zoonotic diseases: Etiology, impact, and control. Microorganisms 2020, 8, 1405. [Google Scholar] [CrossRef]
- Liu, Q.; Cao, L.; Zhu, X.Q. Major emerging and re-emerging zoonoses in China: A matter of global health and socioeconomic development for 1.3 billion. Int. J. Infect. Dis. 2014, 25, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Neyra, R.; Zegarra, E.; Monroy, Y.; Bernedo, R.F.; Cornejo-Rosello, I.; Paz-Soldan, V.A.; Levy, M.Z. Spatial association of canine rabies outbreak and ecological urban corridors, Arequipa, Peru. Trop. Med. Infect. Dis. 2017, 2, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Dávila, J.D.; Allen, A.; Haklay, M.; Tacoli, C.; Fèvre, E.M. Does urbanization make emergence of zoonosis more likely? Evidence, myths, and gaps. Environ. Urban. 2019, 31, 443–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benavides, J.A.; Valderrama, W.; Recuenco, S.; Uieda, W.; Suzán, G.; Avila-Flores, R.; Velasco-Villa, A.; Almeida, M.; de Andrade, F.A.G.; Molina-Flores, B.; et al. Defining new pathways to manage the ongoing emergence of bat Rabies in Latin America. Viruses 2020, 12, 1002. [Google Scholar] [CrossRef]
- Muehlenbein, M.P.; Angelo, K.M.; Schlagenhauf, P.; Chen, L.; Grobusch, M.P.; Gautret, P.; Duvignaud, A.; Chappuis, F.; Kain, K.C.; Bottieau, E.; et al. Traveller exposures to animals: A GeoSentinel analysis. J. Travel Med. 2021, 27, taaa010. [Google Scholar] [CrossRef]
- Gautret, P.; Diaz-Menendez, M.; Goorhuis, A.; Wallace, R.M.; Msimang, V.; Blanton, J.; Dacheux, L.; Parize, P.; Blumberg, L.; Bourhy, H.; et al. Epidemiology of rabies cases among international travellers, 2013–2019: A retrospective analysis of published reports. Travel Med. Infect. Dis. 2020, 36, 101766. [Google Scholar] [CrossRef]
- Polak, K. Dog Transport and Infectious Disease Risk: An International Perspective. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 599–613. [Google Scholar] [CrossRef]
- Seetahal, J.F.R.; Vokaty, A.; Vigilato, M.A.N.; Carrington, C.V.F.; Pradel, J.; Louison, B.; van Sauers, A.; Roopnarine, R.; González Arrebato, J.C.; Millien, M.F.; et al. Rabies in the Caribbean: A situational analysis and historic review. Trop. Med. Infect. Dis. 2018, 3, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S.A.; Chipman, R.B.; Slate, D.; Huyvaert, K.P.; VerCauteren, K.C.; Gilbert, A.T. Management, and modeling approaches for controlling raccoon rabies: The road to elimination. PLoS Negl. Trop. Dis. 2017, 11, e0005249. [Google Scholar] [CrossRef] [Green Version]
- Brunker, K.; Lemey, P.; Marston, D.A.; Fooks, A.R.; Lugelo, A.; Ngeleja, C.; Hampson, K.; Biek, R. Landscape attributes governing local transmission of an endemic zoonosis: Rabies virus in domestic dogs. Mol. Ecol. 2018, 27, 773–788. [Google Scholar] [CrossRef] [Green Version]
- De Andrade, F.A.G.; Gomes, M.N.; Uieda, W.; Begot, A.L.; Ramos, O.D.S.; Fernandes, M.E.B. Geographical analysis for detecting high-risk areas for bovine/human rabies transmitted by the common hematophagous bat in the Amazon region, Brazil. PLoS ONE 2016, 11, e0157332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bányai, T.; Tamás, P.; Illés, B.; Stankevičiūtė, Ž.; Bányai, Á. Optimization of municipal waste collection routing: Impact of industry 4.0 technologies on environmental awareness and sustainability. Int. J. Environ. Res. Public Health 2019, 16, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, T.N.; Roland, M.E.; Françoise, K.M.M.; Francis, Z.; Livo, E.F.; Clovis, S.T.H. Do open garbage dumps play a role in canine rabies transmission in Biyem-Assi health district in Cameroon? Infect. Ecol. Epidemiol. 2015, 5, 26055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thahaby, N.; Akand, A.H.; Hamdani, S.A.; Bhat, A.H.; Hussain, S.A.; Shiekh, I.; Shubeena, S. Epidemiological pattern of dog bites and the occurrence of rabies in humans within Srinagar district of Kashmir Valley, India. Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101556. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Hostedde, A.I.; Mazal, Z.; Jardine, C.M.; Gagnon, J. Enhanced access to anthropogenic food waste is related to hyperglycemia in raccoons (Procyon lotor). Conserv. Physiol. 2018, 6, coy026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brookes, V.J.; Ward, M.P.; Rock, M.; Degeling, C. One Health promotion and the politics of dog management in remote, northern Australian communities. Sci. Rep. 2020, 10, 12451. [Google Scholar] [CrossRef]
- Burgos-Cáceres, S. Canine rabies: A looming threat to public. Animals 2011, 1, 326–342. [Google Scholar] [CrossRef]
- Kennedy, B.P.A.; Cumming, B.; Brown, W.Y. Global strategies for population management of domestic cats (Felis catus): A systematic review to inform best practice management for remote indigenous communities in Australia. Animals 2020, 10, 663. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, E.J.C.; Abrahamian, F.M. Diseases transmitted by cats. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Bedford, E. Number of Dogs in the United States from 2000 to 2017. 2019. Available online: https://www.statista.com/statistics/198100/dogs-in-the-united-states-since-2000/#:~:text=According%20to%20a%20pet%20owners,Why%20has%20this%20figure%20increased%3F (accessed on 20 September 2022).
- Bhalla, S.J.; Kemmers, R.; Vasques, A.; Vanak, A.T. ‘Stray appetites’: A socio-ecological analysis of free-ranging dogs living alongside human communities in Bangalore, India. Urban Ecosyst. 2021, 24, 1245–1258. [Google Scholar] [CrossRef]
- Sikana, L.; Lembo, T.; Hampson, K.; Lushasi, K.; Mtenga, S.; Sambo, M.; Wight, D.; Coutts, J.; Kreppel, K. Dog ownership practices and responsibilities for children’s health in terms of rabies control and prevention in rural communities in Tanzania. PLoS Negl. Trop. Dis. 2021, 15, e0009220. [Google Scholar] [CrossRef] [PubMed]
- Dimkić, I.; Fira, D.; Janakiev, T.; Kabić, J.; Stupar, M.; Nenadić, M.; Unković, N.; Grbić, M.L. The microbiome of bat guano: For what is this knowledge important? Appl. Microbiol. Biotechnol. 2021, 105, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Soler-Rangel, S.; Rosselli, D.; Jiménez-restrepo, N.; Nariño, D. Patient bitten by a domestic cat. J. Sao Paulo Inst. Trop. Med. 2020, 62, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.A.; McManus, D.P.; Jones, M.K.; Gray, D.J.; Gobert, G.N. The increase of exotic zoonotic helminth infections: The impact of urbanization, climate change and globalization. Adv. Parasitol. 2016, 91, 311–397. [Google Scholar]
- Arshad, Z.; Robaina, M.; Shahbaz, M.; Veloso, A.B. The effects of deforestation and urbanization on sustainable growth in Asian countries. Environ Sci Pollut Res. 2020, 27, 10065–10086. [Google Scholar] [CrossRef]
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Müller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Prim. 2017, 3, 17091. [Google Scholar] [CrossRef]
- Fisher, C.R.; Streicker, D.G.; Schnell, M.J. The spread and evolution of rabies virus: Conquering new frontiers. Nat. Rev. Microbiol. 2018, 16, 241–255. [Google Scholar] [CrossRef]
- Rodriguez, J.T.; Lesmeister, D.B.; Levi, T. Mesocarnivore landscape use along a gradient of urban, rural, and forest cover. PeerJ 2021, 9, e11083. [Google Scholar] [CrossRef]
- Orlando, S.A.; Panchana, V.F.; Calderón, J.L.; Muñoz, O.S.; Campos, D.N.; Torres-Lasso, P.R.; Arcos, F.J.; Quentin, E. Risk factors associated with attacks of hematophagous bats (Desmodus rotundus) on cattle in Ecuador. Vector-Borne Zoonotic Dis. 2019, 19, 407–413. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar]
- Helmy, Y.A.; Fawzy, M.; Elaswad, A.; Sobieh, A.; Kenney, S.P.; Shehata, A.A. The COVID-19 pandemic: A comprehensive review of taxonomy, genetics, epidemiology, diagnosis, treatment, and control. J. Clin. Med. 2020, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.A.; Parvin, R.; Nagy, A.; Wang, Y.; Azhar, T.M.; Attia, Y.A.; Azhar, E.I.; Paul, A.K.; Rahmatullah, M. An overview of the ongoing challenges in SARS-CoV-2 global control. Ger. J. Microbiol. 2021, 1, 1–18. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.L. Origin, and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Chinazzi, M.; Davis, J.T.; Ajelli, M.; Gioannini, C.; Litvinova, M.; Merler, S.; Pastore y Piontti, A.; Mu, K.; Rossi, L.; Sun, K.; et al. The effect of travel restrictions on the spread of the 2019 novel coronavirus (COVID-19) outbreak. Science 2020, 368, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Tayoun, A.A.; Loney, T.; Khansaheb, H.; Ramaswamy, S.; Harilal, D.; Deesi, Z.O.; Varghese, R.M.; Al Suwaidi, H.; Alkhajeh, A.; AlDabal, L.M.; et al. Multiple early introductions of SARS-CoV-2 into a global travel hub in the Middle East. Sci. Rep. 2020, 10, 17720. [Google Scholar] [CrossRef]
- Deng, X.; Gu, W.; Federman, S.; du Plessis, L.; Pybus, O.G.; Faria, N.R.; Wang, C.; Yu, G.; Bushnell, B.; Pan, C.Y.; et al. Genomic surveillance reveals multiple introductions of SARS-CoV-2 into Northern California. Science 2020, 369, 582–587. [Google Scholar] [CrossRef]
- Candido, D.S.; Claro, I.M.; de Jesus, J.G.; Souza, W.M.; Moreira, F.R.R.; Dellicour, S.; Mellan, T.A.; du Plessis, L.; Pereira, R.H.M.; Sales, F.C.S.; et al. Evolution and epidemic spread of SARS-CoV-2 in Brazil. Science 2020, 369, 1255–1260. [Google Scholar] [CrossRef]
- Rader, B.; Scarpino, S.V.; Nande, A.; Hill, A.L.; Adlam, B.; Reiner, R.C.; Pigott, D.M.; Gutierrez, B.; Zarebski, A.E.; Shrestha, M.; et al. Crowding, and the shape of COVID-19 epidemics. Nat. Med. 2020, 26, 1829–1834. [Google Scholar] [CrossRef]
- Noyma, N.P.; de Magalhães, L.; Furtado, L.L.; Mucci, M.; van Oosterhout, F.; Huszar, V.L.M.; Marinho, M.M.; Lürling, M. Controlling cyanobacterial blooms through effective flocculation and sedimentation with combined use of flocculants and phosphorus adsorbing natural soil and modified clay. Water Res. 2016, 97, 26–38. [Google Scholar] [CrossRef]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global morbidity and mortality of leptospirosis: A Systematic review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagan, J.E.; Moraga, P.; Costa, F.; Capian, N.; Ribeiro, G.S.; Wunder, E.A.; Felzemburgh, R.D.M.; Reis, R.B.; Nery, N.; Santana, F.S.; et al. Spatiotemporal determinants of urban leptospirosis transmission: Four-year prospective cohort study of Slum residents in Brazil. PLoS Negl. Trop. Dis. 2016, 10, e0004275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, W.A. Animal leptospirosis. In Leptospira and Leptospirosis: Current Topics in Microbiology and Immunology; Adler, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 387. [Google Scholar] [CrossRef]
- Picardeau, M. Diagnosis and epidemiology of leptospirosis. Med. Mal. Infect. 2013, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar] [PubMed] [Green Version]
- Wasinski, B.; Dutkiewicz, J. Leptospirosis—Current risk factors connected with human activity and the environment. Ann. Agric. Environ. Med. 2013, 20, 239–244. [Google Scholar]
- Bacallao, J.; Schneider, M.C.; Najera, P.; Aldighieri, S.; Soto, A.; Marquiño, W.; Sáenz, C.; Jiménez, E.; Moreno, G.; Chávez, O.; et al. Socioeconomic factors and vulnerability to outbreaks of leptospirosis in Nicaragua. Int. J. Environ. Res. Public Health 2014, 11, 8301–8318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galan, D.I.; Roess, A.A.; Pereira, S.V.C.; Schneider, M.C. Epidemiology of human leptospirosis in urban and rural areas of Brazil, 2000–2015. PLoS ONE 2021, 16, e0247763. [Google Scholar] [CrossRef] [PubMed]
- Kembhavi, R.S.; Velhal, G.D.; Anuradha, K. Shah Epidemiological determinants of leptospirosis in rural and urban districts of Maharashtra, India Ravindra. J. Fam. Med. Prim. Care 2021, 10, 3361–3367. [Google Scholar]
- Tan, W.L.; Soelar, S.A.; Suan, M.A.M.; Hussin, N.; Cheah, W.K.; Verasahib, K.; Goh, P.P. Leptospirosis incidence and mortality in Malaysia. Southeast Asian J. Trop. Med. Public Health 2016, 47, 434–440. [Google Scholar]
- Cosson, J.F.; Picardeau, M.; Mielcarek, M.; Tatard, C.; Chaval, Y.; Suputtamongkol, Y.; Buchy, P.; Jittapalapong, S.; Herbreteau, V.; Morand, S. Epidemiology of leptospira transmitted by rodents in Southeast Asia. PLoS Negl. Trop. Dis. 2014, 8, e2902. [Google Scholar] [CrossRef]
- Blessington, T.; Schenck, A.P.; Levine, J.F. Frequency of animal leptospirosis in the Southern United States and the implications for human health. South. Med. J. 2020, 113, 240–249. [Google Scholar] [CrossRef] [PubMed]
- de Vries, S.G.; Visser, B.J.; Nagel, I.M.; Goris, M.G.A.; Hartskeerl, R.A.; Grobusch, M.P. Leptospirosis in Sub-Saharan Africa: A systematic review. Int. J. Infect. Dis. 2014, 28, e47–e64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelloni, F.; Cilia, G.; Turchi, B.; Pinzauti, P.; Cerri, D.; Fratini, F. Epidemiology of leptospirosis in North-Central Italy: Fifteen years of serological data (2002–2016). Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Un-Habitat. Enhancing Urban Safety and Security: Global Report on Human Settlements 2007; Routledge: London, UK, 2012. [Google Scholar]
- Fry, S.; Cousins, B.; Olivola, K. Health of Children Living in Urban Slums in Asia and the Near East: Review of Existing Literature and Data; Environmental health project, US Agency for International Development: Washington, DC, USA, 2002. Available online: https://pdf.usaid.gov/pdf_docs/PNACQ101.pdf (accessed on 1 September 2022).
- Faine, S. Leptospira and Leptospirosis; CRC Press Inc.: Boca Raton, FL, USA, 1994. [Google Scholar]
- Blasdell, K.R.; Morand, S.; Perera, D.; Firth, C. Association of rodent-borne leptospira spp. with urban environments in Malaysian Borneo. PLoS Negl. Trop. Dis. 2019, 13, e0007141. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.; Singh, R.; Gupta, B.; Patel, R.; Soni, D.; Dhakad, B.; Reddy, B.M.; Gupta, S.; Sharma, N. Leptospira: An emerging zoonotic pathogen of climate change, global warming, and unplanned urbanization: A review. J. Entomol. Zool. Stud. 2021, 9, 564–571. [Google Scholar] [CrossRef]
- Vinetz, J.M.; Glass, G.E.; Flexner, C.E.; Mueller, P.; Kaslow, D.C. Sporadic urban leptospirosis. Ann. Intern. Med. 1996, 125, 794–798. [Google Scholar] [CrossRef]
- Adler, H.; Vonstein, S.; Deplazes, P.; Stieger, C.; Frei, R. Prevalence of Leptospira spp. in various species of small mammals caught in an inner-city area in Switzerland. Epidemiol. Infect. 2002, 128, 107–109. [Google Scholar] [CrossRef]
- Van Delden, L.; Larsen, E.; Rowlings, D.; Scheer, C.; Grace, P. Establishing turf grass increases soil greenhouse gas emissions in peri-urban environments. Urban Ecosyst. 2016, 19, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Dechet, A.M.; Parsons, M.; Rambaran, M.; Mohamed-Rambaran, P.; Florendo-Cumbermack, A.; Persaud, S.; Baboolal, S.; Ari, M.D.; Shadomy, S.V.; Zaki, S.R.; et al. Leptospirosis outbreak following severe flooding: A rapid assessment and mass prophylaxis campaign; Guyana, January–February 2005. PLoS ONE 2012, 7, e39672. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.L.; Smythe, L.D.; Craig, S.B.; Weinstein, P. Climate change, flooding, urbanisation and leptospirosis: Fuelling the fire? Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 631–638. [Google Scholar] [CrossRef]
- Vijayachari, P.; Sugunan, A.P.; Shriram, A.N. Leptospirosis: An emerging global public health problem. J. Biosci. 2008, 33, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Kupek, E.; de Sousa Santos Faversani, M.C.; de Souza Philippi, J.M. The relationship between rainfall and human leptospirosis in Florianópolis, Brazil, 1991–1996. Braz. J. Infect. Dis. 2000, 4, 131–134. [Google Scholar] [PubMed]
- Mwachui, M.A.; Crump, L.; Hartskeerl, R.; Zinsstag, J.; Hattendorf, J. Environmental and behavioral determinants of leptospirosis transmission: A systematic review. PLoS Negl. Trop. Dis. 2015, 9, e0003843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prescott, J.F.; McEwen, B.; Taylor, J.; Woods, J.P.; Abrams-Ogg, A.; Wilcock, B. Resurgence of leptospirosis in dogs in Ontario: Recent findings. Can. Vet. J. 2002, 43, 955–961. [Google Scholar] [PubMed]
- André-Fontaine, G. Canine leptospirosis—Do we have a problem? Vet. Microbiol. 2006, 117, 19–24. [Google Scholar] [CrossRef]
- Hotez, P.J.; Gurwith, M. Europe’s neglected infections of poverty. Int. J. Infect. Dis. 2011, 15, e611–e619. [Google Scholar] [CrossRef] [Green Version]
- Cruz, L.S.; Vargas, R.; Lopes, A.A. Leptospirosis: A worldwide resurgent zoonosis and important cause of acute renal failure and death in developing nations. Ethn. Dis. 2009, 19 (Suppl. 1), 37–41. [Google Scholar]
- Childs, J.E.; Schwartz, B.S.; Ksiazek, T.G.; Graham, R.R.; LeDuc, J.W.; Glass, G.E. Risk factors associated with antibodies to leptospires in inner-city residents of Baltimore: A protective role for cats. Am. J. Public Health 1992, 82, 597–599. [Google Scholar] [CrossRef] [Green Version]
- Demers, R.Y.; Thiermann, A.; Demers, P.; Frank, R. Exposure to Leptospira icterohaemorrhagiae in inner-city and suburban children: A serologic comparison. J. Fam. Pract. 1983, 17, 1007–1011. [Google Scholar]
- Zhang, X.; House, L.A. How Pet Owner Involvement in Pet Care Influences Veterinary Service Use and Expenditure. In Proceedings of the Annual Meeting of the Agricultural and Applied Economics Association, Austin, TX, USA, 1–3 August 2021. [Google Scholar]
- The European Food Industry (FEDIAF) Annual Report. 2020. Available online: https://svpdz.cz/soubory/zobrazit/ed57b2ad95ce98d5d9ccbc5ea3f6d8167672e74c (accessed on 20 September 2022).
- Tufani, N.A. Microscopic Agglutination Test (MAT) for Leptospirosis in association with acute renal failure in dogs. J. Anim. Res. 2019, 9, 581–584. [Google Scholar] [CrossRef]
- Pratt, N.; Rajeev, S. Leptospira seroprevalence in animals in the Caribbean region: A systematic review. Acta Trop. 2018, 182, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Ananyina, Y.V. Human leptospirosis in Russia: Epidemiological trends across two decades. In Proceedings of the European Meeting of Leptospirosis (Eurolept), Dubrovnic, Croatia, 31 May–2 June 2012; p. 38. [Google Scholar]
- Jansen, A.; Luge, E.; Guerra, B.; Wittschen, P.; Gruber, A.D.; Loddenkemper, C.; Schneider, T.; Lierz, M.; Ehlert, D.; Appel, B.; et al. Leptospirosis in urban wild boars, Berlin, Germany. Emerg. Infect. Dis. 2007, 13, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.S. Epidemiology of Lyme Disease. Infect. Dis. Clin. North Am. 2015, 29, 187–210. [Google Scholar] [CrossRef] [PubMed]
- Ogrinc, K.; Lusa, L.; Lotrič-Furlan, S.; Bogovič, P.; Stupica, D.; Cerar, T.; Ružić-Sabljić, E.; Strle, F. Course and outcome of early European Lyme neuroborreliosis (Bannwarth syndrome): Clinical and laboratory findings. Clin. Infect. Dis. 2016, 63, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Steere, A.C. Lyme disease. N. Engl. J. Med. 1989, 321, 586–596. [Google Scholar] [CrossRef]
- Kim, C.M.; Lee, J.B.; Shin, S.J.; Ahn, J.B.; Lee, M.; Kim, H.S. The efficacy of immune checkpoint inhibitors in elderly patients: A meta-analysis and meta-regression. ESMO Open 2022, 7, 100577. [Google Scholar] [CrossRef]
- Steere, A.C.; Malawista, S.E.; Snydman, D.R.; Shope, R.E.; Andiman, W.A.; Ross, M.R.; Steele, F.M. Lyme arthritis: An epidemic of oligoarticular arthritis in children and adults in three Connecticut communities. Arthritis Rheum. 1977, 20, 7–17. [Google Scholar] [CrossRef]
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme borreliosis. Lancet 2012, 379, 461–473. [Google Scholar] [CrossRef]
- James, M.C.; Bowman, A.S.; Forbes, K.J.; Lewis, F.; McLeod, J.E.; Gilbert, L. Environmental determinants of Ixodes ricinus ticks and the incidence of Borrelia burgdorferi sensu lato, the agent of Lyme borreliosis, in Scotland. Parasitology 2013, 140, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Chao, L.L.; Chen, Y.J.; Shih, C.M. First isolation and molecular identification of Borrelia burgdorferi sensu stricto and Borrelia afzelii from skin biopsies of patients in Taiwan. Int. J. Infect. Dis. 2011, 15, e182–e187. [Google Scholar] [CrossRef] [Green Version]
- Pritt, B.S.; Mead, P.S.; Johnson, D.K.H.; Neitzel, D.F.; Respicio-Kingry, L.B.; Davis, J.P.; Schiffman, E.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; et al. Identification of a novel pathogenic Borrelia species causing Lyme borreliosis with unusually high spirochaetaemia: A descriptive study. Lancet Infect Dis. 2016, 16, 556–564. [Google Scholar] [CrossRef] [Green Version]
- Clark, K.; Hendricks, A.; Burge, D. Molecular identification, and analysis of Borrelia burgdorferi sensu lato in lizards in the southeastern United States. Appl. Environ. Microbiol. 2005, 71, 2616–2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M.; Levine, J.F.; Yang, S.; Howard, P.; Apperson, C.S. Reservoir competence of the southeastern five-lined skink (Eumeces inexpectatus) and the green anole (Anolis carolinensis) for Borrelia burgdorferi. Am. J. Trop. Med. Hyg. 1996, 54, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.S.; Piesman, J.; Burgdorfer, W. Lyme borreliosis: Relation of its causative agent to its vectors and hosts in North America and Europe. Annu. Rev. Entomol. 1991, 36, 587–609. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Lane, R.S.; Barbour, A.G.; Gresbrink, R.A.; Anderson, J.R. The western black-legged tick, Ixodes pacificus: A vector of Borrelia burgdorferi. Am. J. Trop. Med. Hyg. 1985, 34, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Gern, L.; Humair, P.F. Ecology of Borrelia burgdorferi sensu lato in Europe. In Lyme Borreliosis: Biology, Epidemiology and Control; CABI International: New York, NY, USA, 2002. [Google Scholar] [CrossRef] [Green Version]
- Korenberg, E.I.; Gorelova, N.B.; Kovalevskii, Y.V. Ecology of Borrelia burgdorferi sensu lato in Russia. In Lyme Borreliosis: Biology, Epidemiology and Control; CABI International: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Malak, N.; Niaz, S.; Wadood, A.; Nasreen, N.; Ali, I.; Iqbal, J.; Swelum, A.A.; ahmed, E.A.; Alkahtani, M.A.; Zając, Z.; et al. In silico approaches to develop herbal acaricides against R. (Boophilus) Microplus and in vitro anti-tick activities of selected medicinal plants. Saudi J. Biol. Sci. 2022, 29, 103302. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Lees, A.D. The water balance in Ixodes ricinus L. and certain other species of ticks. Parasitology. 1946, 37, 1–20. [Google Scholar] [CrossRef]
- Gassner, F.; van Vliet, A.J.; Burgers, S.L.; Jacobs, F.; Verbaarschot, P.; Hovius, E.K.; Mulder, S.; Verhulst, N.O.; van Overbeek, L.S.; Takken, W. Geographic and temporal variations in population dynamics of Ixodes ricinus and associated Borrelia infections in The Netherlands. Vector Borne Zoonotic Dis. 2011, 11, 523–532. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-peña, A.; George, J.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.; Jensen, P.M.; et al. Artikel Ixodes Ricinus Europa, 2013. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S. Ixodes ricinus seasonal activity: Implications of global warming indicated by revisiting tick and weather data. Int. J. Med. Microbiol. 2008, 298, 19–24. [Google Scholar] [CrossRef]
- Nilsson, A. Seasonal Occurrence of Ixodes Ricinus (Acari) in vegetation and on small mammals in Southern Sweden. Holarct. Ecol. 1988, 11, 161–165. [Google Scholar] [CrossRef]
- Schulz, M.; Mahling, M.; Pfister, K. Abundance, and seasonal activity of questing Ixodes ricinus ticks in their natural habitats in southern Germany in 2011. J. Vector Ecol. 2014, 39, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Grigoryeva, L.A.; Tokarevich, N.K.; Freilikhman, O.A.; Samoylova, E.P.; Lunina, G.A. Seasonal changes in populations of sheep tick, Ixodes ricinus (L., 1758) (Acari: Ixodinae) in natural biotopes of St. Petersburg and Leningrad province, Russian Federation. Syst. Appl. Acarol. 2019, 24, 701–710. [Google Scholar] [CrossRef]
- Fish, D. Population ecology of Ixodes damini. In Ecology and Environmental Management of Lyme Disease; Ginsberg, H., Ed.; Rutgers University Press: New Brunswick, NJ, USA, 1993; pp. 25–42. [Google Scholar]
- Murphree Bacon, R.; Kugeler, K.J.; Mead, P.S. Surveillance for Lyme disease—United States, 1992–2006. In CDC Surveillance Summaries: Morbidity and Mortality Weekly Report; CDC: Washington, DC, USA, 2008; Volume 57, p. SS-10. [Google Scholar]
- Piesman, J.; Gern, L. Lyme borreliosis in Europe and North America. Parasitology 2004, 129, 191–220. [Google Scholar] [CrossRef] [PubMed]
- Kugeler, K.J.; Farley, G.M.; Forrester, J.D.; Mead, P.S. Geographic distribution and expansion of human Lyme disease, United States. Emerg. Infect. Dis. 2015, 21, 1455–1457. [Google Scholar] [CrossRef]
- Brownstein, J.S.; Holford, T.R.; Fish, D. Effect of climate change on Lyme disease risk in North America. EcoHealth 2005, 2, 38–46. [Google Scholar] [CrossRef] [Green Version]
- WHO. Tuberculosis. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 29 October 2022).
- McIver, L.; Viney, K.; Harley, D.; Hanna, L.; Kienene, T. Climate change, overcrowding and non-communicable diseases: The “triple whammy” of tuberculosis transmission risk in Pacific atoll countries. Ann. Australas. Coll. Trop. Med. 2015, 16, 57–61. [Google Scholar]
- Sinha, P.; Lönnroth, K.; Bhargava, A.; Heysell, S.K.; Sarkar, S.; Salgame, P.; Rudgard, W.; Boccia, D.; van Aartsen, D.; Hochberg, N.S. Food for thought: Addressing undernutrition to end tuberculosis. Lancet Infect. Dis. 2021, 21, e318–e325. [Google Scholar] [CrossRef]
- Sinha, P.; Carwile, M.E.; Cintron, C.; de Perez, E.C.; Hochberg, N.S. Climate change and TB: The soil and seed conceptual framework. Public Health Action 2021, 11, 108. [Google Scholar] [CrossRef]
- Harries, A.D.; Martinez, L.; Chakaya, J.M. Reply to: Climate change and TB: The soil and seed conceptual framework. Public Health Action 2021, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Rosypal, A.C.; Zajac, A.M.; Lindsay, D.S. Canine visceral leishmaniasis and its emergence in the United States. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 921–937. [Google Scholar] [CrossRef]
- Baneth, G. Leishmaniasis. In Infectious Diseases of the Dog and Cat, 3rd ed.; Greene, C.E., Ed.; W.B. Saunders: Philadelphia, PA, USA, 2006; pp. 685–698. [Google Scholar]
- WHO. WHA60.13: Control of Leishmaniasis. 2007. Available online: https://www.who.int/publications/i/item/wha60.13 (accessed on 30 October 2022).
- WHO. Health topics: Leishmaniasis. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 20 October 2022).
- Ashford, R.W. Leishmaniasis reservoirs and their significance in control. Clin. Dermatol. 1996, 14, 523–532. [Google Scholar] [CrossRef]
- Luppi, M.M.; Malta, M.C.C.; Silva, T.M.A.; Silva, F.L.; Motta, R.O.C.; Miranda, I.; Ecco, R.; Santos, R.L. Visceral leishmaniasis in captive wild canids in Brazil. Vet. Parasitol. 2008, 155, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Diniz, S.A.; Silva, F.L.; Carvalho Neta, A.C.; Bueno, R.; Guerra, R.M.S.N.C.; Abreu-Silva, A.L.; Santos, R.L. Animal reservoirs for visceral leishmaniasis in densely populated urban areas. J. Infect. Dev. Ctries. 2008, 2, 24–33. [Google Scholar]
- Marzochi, M.C.d.A.; Marzochi, K.B.F. Tegumentary and visceral leishmaniases in Brazil: Emerging anthropozoonosis and possibilities for their control. Cad. Saude Publica 1994, 10, 359–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siviero Do Vale, E.C.; Furtado, T. Tegumentary leishmaniasis in Brazil: A historical review related to the origin, expansion, and etiology. An. Bras. Dermatol. 2005, 80, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Travi, B.L.; Adler, G.H.; Lozano, M.; Cadena, H.; Montoya-Lerma, J. Impact of habitat degradation on Phlebotominae (Diptera: Psychodidae) of Tropical Dry Forests in Northern Colombia. J. Med. Entomol. 2002, 39, 451–456. [Google Scholar] [CrossRef]
- De Oliveira Góes, M.A.; Moura de Melo, C.; de Lourdes Sierpe Jeraldo, V. Time series of visceral leishmaniasis in Aracaju, state of Sergipe, Brazil (1999 to 2008): Human and canine aspects. Ver. Bras. Epidemiol. 2012, 15, 298–307. [Google Scholar] [CrossRef]
- Boggiatto, P.M.; Gibson-Corley, K.N.; Metz, K.; Gallup, J.M.; Hostetter, J.M.; Mullin, K.; Petersen, C.A. Transplacental transmission of Leishmania infantum as a means for continued disease incidence in North America. PLoS Negl. Trop. Dis. 2011, 5, e1019. [Google Scholar] [CrossRef]
- Boechat, V.C.; Mendes Junior, A.A.V.; Madeira, M.d.F.; Ferreira, L.C.; Figueiredo, F.B.; Rodrigues, F.d.C.d.C.; Oliveira, V.d.C.; de Oliveira, R.d.V.C.; Menezes, R.C. Occurrence of Leishmania infantum and associated histological alterations in the genital tract and mammary glands of naturally infected dogs. Parasitol. Res. 2016, 115, 2371–2379. [Google Scholar] [CrossRef] [PubMed]
- Naucke, T.J.; Lorentz, S. First report of venereal and vertical transmission of canine leishmaniosis from naturally infected dogs in Germany. Parasites Vectors 2012, 5, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otranto, D.; Dantas-Torres, F. The prevention of canine leishmaniasis and its impact on public health. Trends Parasitol. 2013, 29, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J.; Albery, G.F.; Merow, C.; Trisos, C.H.; Zipfel, C.M.; Eskew, E.A.; Olival, K.J.; Ross, N.; Bansal, S. Climate change increases cross-species viral transmission risk. Nature 2022, 607, 555–562. [Google Scholar] [CrossRef]
- Ready, P.D. Leishmaniasis emergence and climate change. OIE Rev. Sci. Tech. 2008, 27, 399–412. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Brandão-Filho, S.P. Visceral leishmaniasis in Brazil: Revisiting paradigms of epidemiology and control. Rev. Inst. Med. Trop. Sao Paulo 2006, 48, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Kaya, Y.; Dokuzeylül, B.; Bakırel, U.; Or, M.E. Antifungal resistance and clinical significance in small animals. Ger. J. Vet. Res. 2022, 2, 28–36. [Google Scholar] [CrossRef]
- Nnadi, N.E.; Carter, D.A. Climate change and the emergence of fungal pathogens. PLoS Pathog. 2021, 17, e1009503. [Google Scholar] [CrossRef]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef]
- Rhodes, J.; Fisher, M.C. Global epidemiology of emerging Candida auris. Curr. Opin. Microbiol. 2019, 52, 84–89. [Google Scholar] [CrossRef]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the emergence of Candida auris: Climate change, azoles, swamps, and birds. Host-Microbe Biol. 2019, 10, e01397-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Crecy, E.; Jaronski, S.; Lyons, B.; Lyons, T.; Keyhani, N. Directed evolution of a filamentous fungus for thermotolerance. BMC Biotechnol. 2009, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-Montes, M.d.R.; Duarte-Escalante, E.; Frías-De-León, M.G.; Pérez-Rodríguez, A.; Meraz-Ríos, B. Impact of climate change on dermatophytosis. In Impact of Climate Change on Fungal Diseases; fungal biology series; Frías-De-León, M.G., Brunner-Mendoza, C., del Rocío Reyes-Montes, M., Duarte-Escalante, E., Eds.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Casadevall, A. Climate change brings the specter of new infectious diseases. J. Clin. Investig. 2020, 130, 553–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadre, A.; Enbiale, W.; Andersen, L.K.; Coates, S.J. The effects of climate change on fungal diseases with cutaneous manifestations: A report from the International Society of Dermatology Climate Change Committee. J. Clim. Chang. Health 2022, 6, 100156. [Google Scholar] [CrossRef]
- Opulente, D.A.; Langdon, Q.K.; Buh, K.V.; Haase, M.A.B.; Sylvester, K.; Moriarty, R.V.; Jarzyna, M.; Considine, S.L.; Schneider, R.M.; Hittinger, C.T. Pathogenic budding yeasts isolated outside of clinical settings. FEMS Yeast Res. 2019, 19, foz032. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Snelders, E.; Kema, G.H.; Mellado, E.; Melchers, W.J. Azole resistance in Aspergillus fumigatus: A side-effect of environmental fungicide use? Lancet Infect. Dis. 2009, 9, 789–795. [Google Scholar] [CrossRef]
- Shehata, A.A.; Attia, Y.A.; Rahman, M.T.; Basiouni, S.; El-Seedi, H.R.; Azhar, E.I.; Khafaga, A.F.; Hafez, H.M. Diversity of coronaviruses with particular attention to the interspecies transmission of SARS-CoV-2. Animals 2022, 12, 378. [Google Scholar] [CrossRef]
- Kanda, K.; Jayasinghe, A.; Jayasinghe, C.; Yoshida, T. A Regional analysis of the progress of current dog-mediated rabies control and prevention. Pathogens 2022, 11, 1130. [Google Scholar] [CrossRef]
- Tazerji, S.S.; Shahabinejad, F.; Tokasi, M.; Rad, M.A.; Khan, M.S.; Safdar, M.; Filipiak, K.J.; Szarpak, L.; Dzieciatkowski, T.; Jurgiel, J.; et al. Global data analysis and risk factors associated with morbidity and mortality of COVID-19. Gene Rep. 2022, 26, 101505. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tazerji, S.S.; Nardini, R.; Safdar, M.; Shehata, A.A.; Duarte, P.M. An Overview of Anthropogenic Actions as Drivers for Emerging and Re-Emerging Zoonotic Diseases. Pathogens 2022, 11, 1376. https://doi.org/10.3390/pathogens11111376
Tazerji SS, Nardini R, Safdar M, Shehata AA, Duarte PM. An Overview of Anthropogenic Actions as Drivers for Emerging and Re-Emerging Zoonotic Diseases. Pathogens. 2022; 11(11):1376. https://doi.org/10.3390/pathogens11111376
Chicago/Turabian StyleTazerji, Sina Salajegheh, Roberto Nardini, Muhammad Safdar, Awad A. Shehata, and Phelipe Magalhães Duarte. 2022. "An Overview of Anthropogenic Actions as Drivers for Emerging and Re-Emerging Zoonotic Diseases" Pathogens 11, no. 11: 1376. https://doi.org/10.3390/pathogens11111376