
Genetic Characterization of the Poultry Red Mite (Dermanyssus gallinae) in Poland and a Comparison with European and Asian Isolates

Abstract
:1. Introduction
2. Materials and Methods
2.1. Dermanyssus gallinae
2.2. Isolation of Genomic DNA
2.3. PCR Assay
2.4. Sequencing
2.5. Phylogenetic Analysis
2.5.1. Consensus Sequences
2.5.2. Homologous Sequences
2.5.3. Sequence Alignment Analysis
2.5.4. Haplotype Analysis
2.5.5. Generation of a Phylogenetic Tree
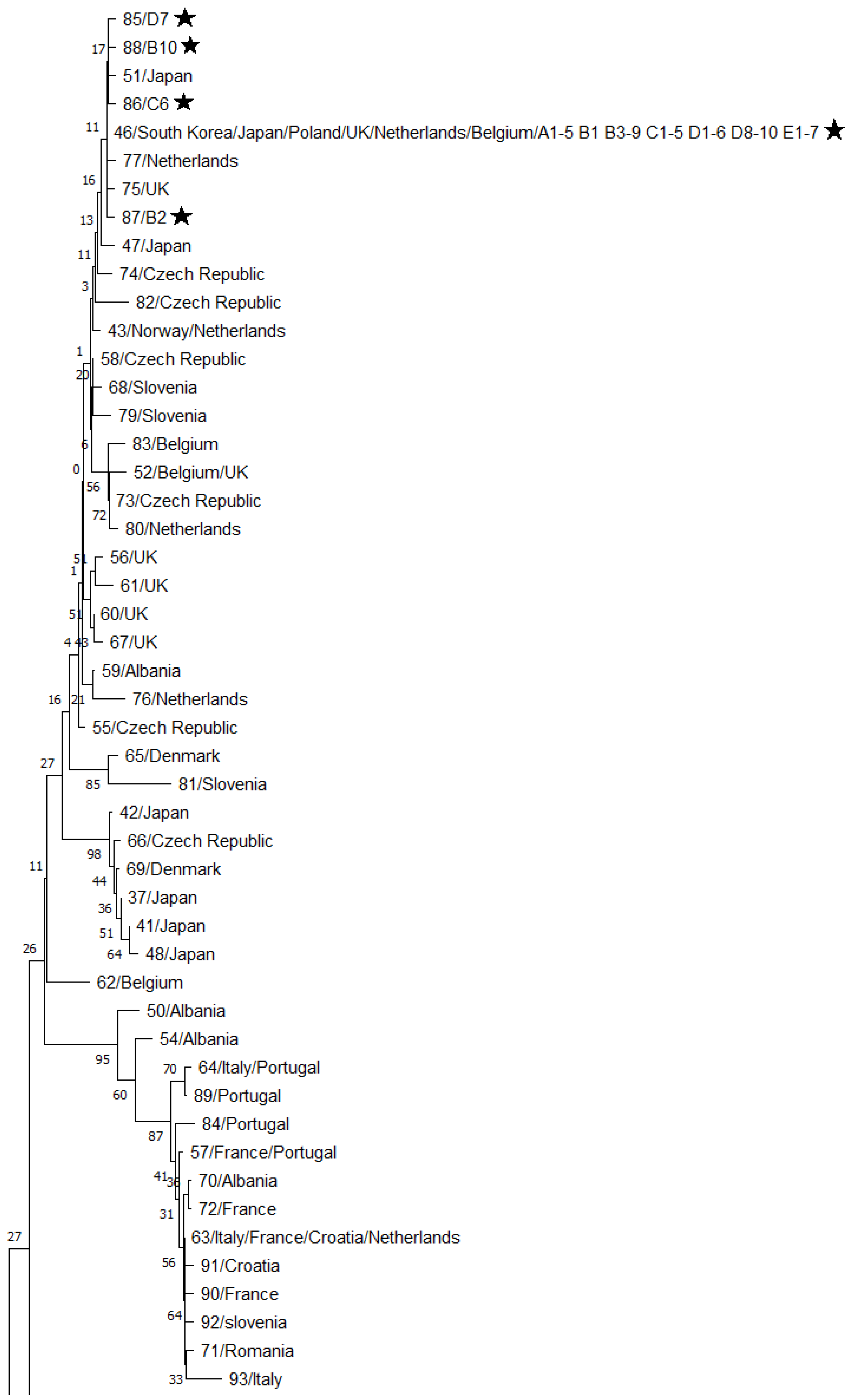
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sparagano, O.A.E.; Pavlicevic, A.; Murano, T.; Camarda, A.; Sahibi, H.; Kilpinen, O.; Mul, M.; Van Emous, R.; Le Bouquin, S.; Hoel, K.; et al. Prevalence and key figures for the poultry red mite Dermanyssus gallinae infections in poultry farm systems. Exp. Appl. Acarol. 2009, 48, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Hoglund, J.; Nordenfors, H.; Uggla, A. Prevalence of the poultry red mite, Dermanyssus gallinae, in different types of production systems for egg layers in Sweden. Poult. Sci. 1995, 74, 1793–1798. [Google Scholar] [CrossRef] [PubMed]
- Cencek, T.; Ziomko, I.; Topór, W. Inwazja Dermanyssus gallinae przyczyną masowych padnięć kacząt brojlerów. Med. Wet. 2002, 58, 353–355. [Google Scholar]
- Guy, J.H.; Khajavi, M.; Hlalel, M.M.; Sparagano, O.A.E. Red mite (Dermanyssus gallinae) prevalence in laying units in northern England. Br. Poult. Sci. 2004, 45, 5–6. [Google Scholar]
- Fiddes, M.D.; Le Gresley, S.; Parsons, D.G.; Epe, C.; Coles, G.C.; Stafford, K.A. Prevalence of the poultry red mite (Dermanyssus gallinae) in England. Vet. Rec. 2005, 157, 233–235. [Google Scholar] [CrossRef]
- Sparagano, O.A.E.; George, D.R.; Harrington, D.W.; Giangaspero, A. Significance and control of the poultry red mite, Dermanyssus gallinae. Annu. Rev. Entomol. 2014, 59, 447–466. [Google Scholar] [CrossRef] [Green Version]
- Sokół, R.; Koziatek-Sadłowska, S.; Michalczyk, M. The influence of Dermanyssus gallinae and different lighting regimens on selected blood proteins, corticosterone levels and egg production in layer hens. Vet. Res. Commun. 2019, 43, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Koziatek-Sadłowska, S.; Sokół, R. Changes in the percentages of B- and T-lymphocytes and antibody titres in laying hens infested with Dermanyssus gallinae—A preliminary study. Animals 2020, 10, 987. [Google Scholar] [CrossRef]
- van Emous, R.A. Wage war against the red mite! Poult. Int. 2005, 44, 26–33. [Google Scholar]
- Flochlay, A.S.; Thomas, E.; Sparagano, O.A.E. Poultry red mite (Dermanyssus gallinae) infestation: A broad impact parasitological disease that still remains a significant challenge for the egg-laying industry in Europe. Parasites Vectors 2017, 10, 357. [Google Scholar] [CrossRef]
- Moss, W.W. The mite genus Dermanyssus: A survey, with description of Dermanyssus trochilinis, n. sp., and a revised key to the species (Acari: Mesostigmata: Dermanyssidae). J. Med. Entomol. 1978, 14, 627–640. [Google Scholar] [CrossRef]
- Roy, L.; Dowling, A.P.G.; Chauve, C.; Buronfosse, T. Delimiting species boundaries within Dermanyssus Dugès, 1834 (Acari: Dermanyssidae) using a total evidence approach. Mol. Phylogenet. Evol. 2009, 50, 446–470. [Google Scholar] [CrossRef] [PubMed]
- Roy, L.; Buronfosse, T. Using mitochondrial and nuclear sequence data for disentangling population structure in complex pest species: A case study with Dermanyssus gallinae. PLoS ONE 2011, 6, e22305. [Google Scholar] [CrossRef] [PubMed]
- Roy, L.; Chauve, C.M.; Buronfosse, T. Contrasted ecological repartition of the northern fowl mite Ornithonyssus sylviarum (Mesostigmata: Macronyssidae) and the chicken red mite Dermanyssus gallinae (Mesostigmata: Dermanyssidae). Acarologia 2010, 50, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Marangi, M.; Cafiero, M.A.; Capelli, G.; Camarda, A.; Sparagano, O.A.E.; Giangaspero, A. Evaluation of the poultry red mite, Dermanyssus gallinae (Acari: Dermanyssidae), susceptibility to some acaricides in field populations from Italy. Exp. Appl. Acarol. 2009, 48, 11–18. [Google Scholar] [CrossRef]
- Øines, Ø.; Brännström, S. Molecular investigations of cytochrome c oxidase subunit I (COI) and the internal transcribed spacer (ITS) in the poultry red mite, Dermanyssus gallinae, in northern Europe and implications for its transmission between laying poultry farms. Med. Vet. Entomol. 2011, 25, 402–412. [Google Scholar] [CrossRef]
- Chu, T.T.; Murano, T.; Uno, Y.; Usui, T.; Yamaguchi, T. Molecular epidemiological characterization of poultry red mite, Dermanyssus gallinae, in Japan. J. Vet. Med. Sci. 2015, 77, 1397–1403. [Google Scholar] [CrossRef]
- Potenza, L.; Cafiero, M.A.; Camarda, A.; La Salandra, G.; Cucchiarini, L.; Dachà, M. Characterization of Dermanyssus gallinae (Acarina: Dermanissydae) by sequence analysis of the ribosomal internal transcribed spacer regions. Vet. Res. Commun. 2009, 33, 611–618. [Google Scholar] [CrossRef]
- Chauve, C. The poultry red mite Dermanyssus gallinae: Current situation and future prospects. Vet. Parasitol. 1998, 8, 364–376. [Google Scholar]
- Marangi, M.; Cantacessi, C.; Sparagano, O.A.; Camarda, A.; Giangaspero, A. Molecular characterization and phylogenetic inferences of Dermanyssus gallinae isolates in Italy within an European framework. Med. Vet. Entomol. 2014, 28, 447–452. [Google Scholar] [CrossRef]
- Sokół, R.; Romaniuk, K. Przebieg i dynamika inwazji Dermanyssus gallinae w fermie kur niosek. Med. Wet. 2007, 63, 484–486. [Google Scholar]
- Di Palma, A.; Giangaspero, A.; Cafiero, M.A.; Germinara, G.S. A gallery of the key characters to ease identification of Dermanyssus gallinae (Acari: Gamasida: Dermanyssidae) and allow differentiation from Ornithonyssus sylviarum (Acari: Gamasida: Macronyssidae). Parasites Vectors 2012, 5, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karp-Tatham, E.; Küster, T.; Angelou, A.; Papadopoulos, E.; Nisbet, A.J.; Xia, D.; Tomley, F.M.; Blake, D.P. Phylogenetic inference using cytochrome c oxidase subunit I (COI) in the poultry red mite, Dermanyssus gallinae in the United Kingdom relative to a European framework. Front. Vet. Sci. 2020, 7, 553. [Google Scholar] [CrossRef]
- Oh, S.; Noh, G.; Yi, S.; Do, Y.; Kim, E.; Yoo, J. Molecular epidemiological characterization of poultry red mite (Dermanyssus gallinae) collected from Korea. Korean J. Vet. Serv. 2019, 42, 161–167. [Google Scholar]
- Gaweł, Ł. Analiza genetyczna populacji ptaszyńca w Polsce. In Proceedings of the III Międzynarodowa Konferencja Techniczna Eimeriana Avia—Aktualne i Przyszłe Wyzwania w Strategiach Kontroli Kokcydiozy i Innych Chorób Inwazyjnych Drobiu—Aspekty Prawne i Terapeutyczne, Wroclaw, Poland, 20–22 February 2020; p. 102. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No. | Number of Haplotypes | Sequence (GenBank ID) | Country | Ref. |
|---|---|---|---|---|
| 1 | 34 | MK599418 | Japan | |
| LC029558, LC029544, LC029537, LC029535, LC029534, LC029533, LC029531, LC029528, LC029524, LC029523, LC029522, LC029521, LC029515, LC029513, LC029512, LC029511, LC029505, LC029494, LC029492, LC029491, LC029488, LC029483, LC029481, LC029480, LC029479, LC029477, LC029476, LC029473, LC029472, LC029469, LC029468, LC029464, LC029460, | Japan | [17] | ||
| 2 | 14 | LC029542, LC029536, LC029504, LC029501, LC029499, LC029493, LC029489, LC029487, LC029482, LC029474, LC029471, LC029470, LC029466, LC029463, | Japan | [17] |
| 3 | 1 | LC029529 | Japan | [17] |
| 4 | 1 | LC029467 | Japan | [17] |
| 5 | 2 | LC029540, LC029503 | Japan | [17] |
| 6 | 2 | LC029520, LC029462 | Japan | [17] |
| 7 | 2 | LC029507, LC029478 | Japan | [17] |
| 8 | 1 | LC029500 | Japan | [17] |
| 9 | 1 | LC029458 | Japan | [17] |
| 10 | 1 | FM208732 | France | [12] |
| 11 | 1 | LC029553 | Japan | [17] |
| 12 | 13 | LC029551, LC029550, LC029549, LC029548, LC029547, LC029526, LC029519, LC029518, LC029517, LC029497, LC029496, | Japan | [17] |
| LR812399 | Greece | [23] | ||
| LR812378 | Great Britain | [23] | ||
| 13 | 1 | LC029502 | Japan | [17] |
| 14 | 1 | LC029465 | Japan | [17] |
| 15 | 1 | LC029525 | Japan | [17] |
| 16 | 1 | AM921857 | France | [12] |
| 17 | 1 | AM921853 | Norway | [12] |
| 18 | 1 | FM208722 | France | [12] |
| 19 | 1 | FM207497 | Netherlands | [12] |
| 20 | 1 | FM207495 | Netherlands | [12] |
| 21 | 1 | AM921858 | France | [12] |
| 22 | 1 | FM208739 | France | [12] |
| 23 | 1 | AM921856 | Denmark | [12] |
| 24 | 1 | AM921863 | France | [12] |
| 25 | 1 | FM207498 | Netherlands | [12] |
| 26 | 1 | AM921865 | France | [12] |
| 27 | 50 | LR812452, LR812451, LR812450, LR812449, LR812448, LR812447, LR812433, LR812432, LR812431, LR812430, LR812429, LR812428, LR812427, LR812418, LR812417, LR812416, LR812415, LR812414, LR812413, LR812412, LR812411, LR812410, LR812409, LR812408, LR812407, LR812405, LR812404, LR812403, LR812402, LR812398, LR812397, LR812396, LR812395, LR812394, LR812393, LR812392, LR812391, | Greece | [23] |
| LR812406 | Slovenia | [23] | ||
| LR812390, LR812373, LR812372, LR812371, LR812370, LR812369, | Great Britain | [23] | ||
| LR812346, LR812345, LR812343, LR812342, LR812341, LR812340, | Romania | [23] | ||
| LR812406, LR812406, LR812406 | Slovenia | [23] | ||
| 28 | 1 | LR812383 | Great Britain | [23] |
| 29 | 1 | LR812375 | Great Britain | [23] |
| 30 | 3 | LR812339, LR812333, LR812332, | Belgium | [23] |
| 31 | 3 | LR812312, LR812311, LR812310 | Portugal | [23] |
| 32 | 2 | LR812290, LR812289 | Albania | [23] |
| 33 | 3 | LR812477, LR812377, LR812376.1 | Great Britain | [23] |
| 34 | 5 | LR812476, LR812475, LR812474, LR81247, LR812472 | Great Britain | [23] |
| 35 | 1 | LR812434 | Greece | [23] |
| 36 | 1 | LR812374 | Great Britain | [23] |
| 37 | 1 | LC029552 | Japan | [17] |
| 38 | 1 | LR812317 | Czechia | [23] |
| 39 | 1 | LR812420 | Greece | [23] |
| 40 | 5 | LR812351, LR812350, LR812349, LR812348, LR812347 | Turkey | [23] |
| 41 | 19 | LC029557, LC029556, LC029554, LC029546, LC029545, LC029541, LC029532, LC029527, LC029514, LC029510, LC029509, LC029506, LC029498, LC029490, LC029486, LC029485, LC029475, LC029459, LC029457, | Japan | [17] |
| 42 | 1 | LC029484 | Japan | [17] |
| 43 | 2 | AM921852 | Norway | [12] |
| LR812361 | Netherlands | [23] | ||
| 44 | 27 | LR812446, LR812445, LR812444, LR812443, LR812442, LR812441, LR812440, LR812439, LR812438, LR812437, LR812436, LR812435, LR812426, LR812425, LR812424, LR812423, LR812422, LR812421, LR812419, LR812401, LR812400 | Netherlands | [23] |
| LR812385, LR812384, LR812381, LR812380 LR812379 | Great Britain | [23] | ||
| LR812367 | Netherlands | [23] | ||
| 45 | 1 | LR812352 | Turkey | [23] |
| 46 | 55 | MN249083, MN249082, MN249080, MN249079, MN249078, MN249077, MN249076, MN249075, MN249074, MN249073, MN249072 | South Korea | [24] |
| LC029538, LC029516, LC029508, LC029495 | Japan | [17] | ||
| AM921854 | Poland | [12] | ||
| LR812388 | Great Britain | [23] | ||
| LR812363, LR812362 | Netherlands | [23] | ||
| LR812337, LR812336 | Belgium | [23] | ||
| A1-A5, B1, B3-9, C1-5, D 1-6, D8-10, E1-7 (OL547403-436) | Poland | |||
| 47 | 2 | LC029543, LC029461 | Japan | [17] |
| 48 | 1 | LC029539 | Japan | [17] |
| 49 | 1 | LR812334 | Belgium | [23] |
| 50 | 1 | MT232060 | Albania | |
| 51 | 2 | LC029555, LC029530 | Japan | [17] |
| 52 | 2 | FM208717 | Belgium | [12] |
| LR812470 | Great Britain | [23] | ||
| 53 | 1 | LR812344 | Romania | [23] |
| 54 | 1 | MT232059 | Albania | |
| 55 | 1 | LR812321 | Czechia | [23] |
| 56 | 2 | LR812468 LR812467 | Great Britain | [23] |
| 57 | 4 | AM921864 | France | [12] |
| LR812307 | Portugal | [23] | ||
| LR812458, LR812455 | Italy | [23] | ||
| 58 | 2 | LR812319, LR812318 | Czechia | [23] |
| 59 | 1 | LR812288 | Albania | [23] |
| 60 | 8 | LR812469, LR812466, LR812465, LR812464, LR812463, LR812462, LR812387, LR812386 | Great Britain | [23] |
| 61 | 1 | LR812389 | Great Britain | [23] |
| 62 | 1 | LR812335 | Belgium | [23] |
| 63 | 22 | MT230034, MT230032, KY025552 | Italy | |
| FM208725, FM208718, FM208719, FM208733 | France | [12] | ||
| LR812301, LR812300, LR812299, LR812298, | France | [23] | ||
| LR812296, LR812294, LR812293 | Croatia | [23] | ||
| LR812460.1, LR812459, LR812457, LR812456, LR812454, LR812453 | Italy | [23] | ||
| LR812366, LR812365 | Netherlands | [23] | ||
| 64 | 4 | MT230033 | Italy | |
| LR812305, LR812304, LR812303 | Portugal | [23] | ||
| 65 | 4 | LR812325, LR812324, LR812323, LR812326 | Denmark | [23] |
| 66 | 3 | LR812315, LR812314, LR812313 | Czechia | [23] |
| 67 | 1 | LR812382 | Great Britain | [23] |
| 68 | 1 | LR812357 | Slovenia | [23] |
| 69 | 5 | LR812331, LR812330, LR812329, LR812328, LR812327 | Denmark | [23] |
| 70 | 8 | MT232061, LR812292, LR812291, LR812287, LR812286, LR812285, LR812284, LR812140 | Albania | [23] |
| 71 | 2 | KX984130, KX984129 | Romania | |
| 72 | 1 | FM208737 | France | [12] |
| 73 | 1 | LR812320 | Czechia | [23] |
| 74 | 1 | LR812316 | Czechia | [23] |
| 75 | 1 | LR812471 | Great Britain | [23] |
| 76 | 1 | LR812364 | Netherlands | [23] |
| 77 | 1 | LR812360 | Netherlands | [23] |
| 78 | 1 | LR812359 | Slovenia | [23] |
| 79 | 3 | LR812358, LR812355, LR812353 | Slovenia | [23] |
| 80 | 1 | LR812368 | Netherlands | [23] |
| 81 | 1 | LR812356 | Slovenia | [23] |
| 82 | 1 | LR812322 | Czechia | [23] |
| 83 | 1 | LR812338 | Belgium | [23] |
| 84 | 1 | LR812308 | Portugal | [23] |
| 85 | 1 | D7 (OL547437) | Poland | |
| 86 | 1 | C6 (OL547438) | Poland | |
| 87 | 1 | B2 (OL547439) | Poland | |
| 88 | 1 | B10 (OL547440) | Poland | |
| 89 | 1 | LR812306 | Portugal | [23] |
| 90 | 2 | LR812302, LR812297 | France | [23] |
| 91 | 1 | LR812295 | Croatia | [23] |
| 92 | 1 | LR812354 | Slovenia | [23] |
| 93 | 1 | LR812461 | Italy | [23] |
| Sampling Site (Farm) | Haplotype Number | ||||
|---|---|---|---|---|---|
| 46 | 85 | 86 | 87 | 88 | |
| A | 100% | - | - | - | - |
| B | 80% | - | - | 10% | 10% |
| C | 83% | - | 17% | - | - |
| D | 90% | 10% | - | - | - |
| E | 100% | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koziatek-Sadłowska, S.; Sokół, R. Genetic Characterization of the Poultry Red Mite (Dermanyssus gallinae) in Poland and a Comparison with European and Asian Isolates. Pathogens 2022, 11, 1301. https://doi.org/10.3390/pathogens11111301
Koziatek-Sadłowska S, Sokół R. Genetic Characterization of the Poultry Red Mite (Dermanyssus gallinae) in Poland and a Comparison with European and Asian Isolates. Pathogens. 2022; 11(11):1301. https://doi.org/10.3390/pathogens11111301
Chicago/Turabian StyleKoziatek-Sadłowska, Sylwia, and Rajmund Sokół. 2022. "Genetic Characterization of the Poultry Red Mite (Dermanyssus gallinae) in Poland and a Comparison with European and Asian Isolates" Pathogens 11, no. 11: 1301. https://doi.org/10.3390/pathogens11111301