

Immunoproteomic Analysis of Trichinella britovi Proteins Recognized by IgG Antibodies from Meat Juice of Carnivores Naturally Infected with T. britovi

Abstract
:1. Introduction
2. Results
2.1. ELISA and Immunoblot Results
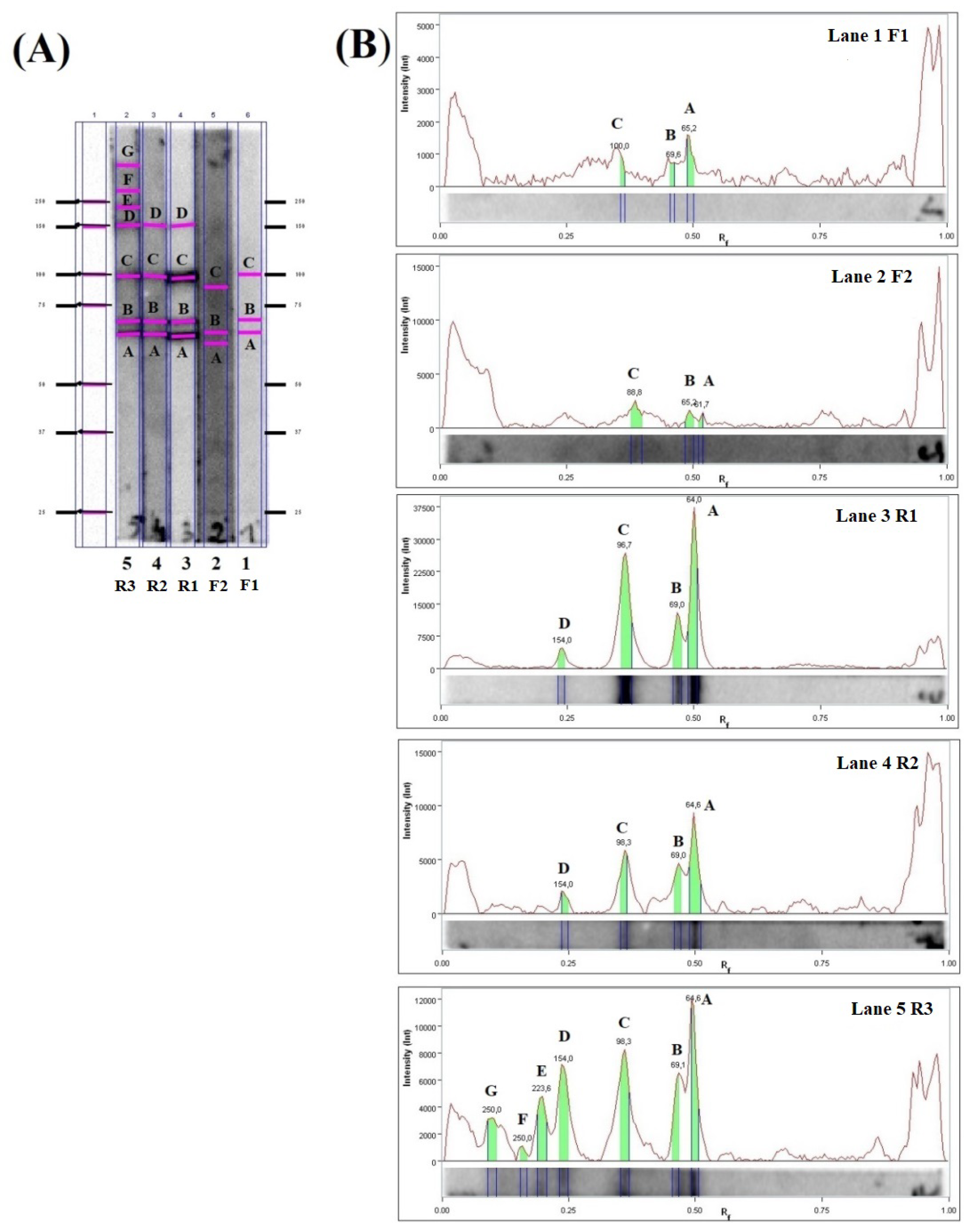
2.2. Immunoblot Analysis
2.3. LC-MS/MS Analysis
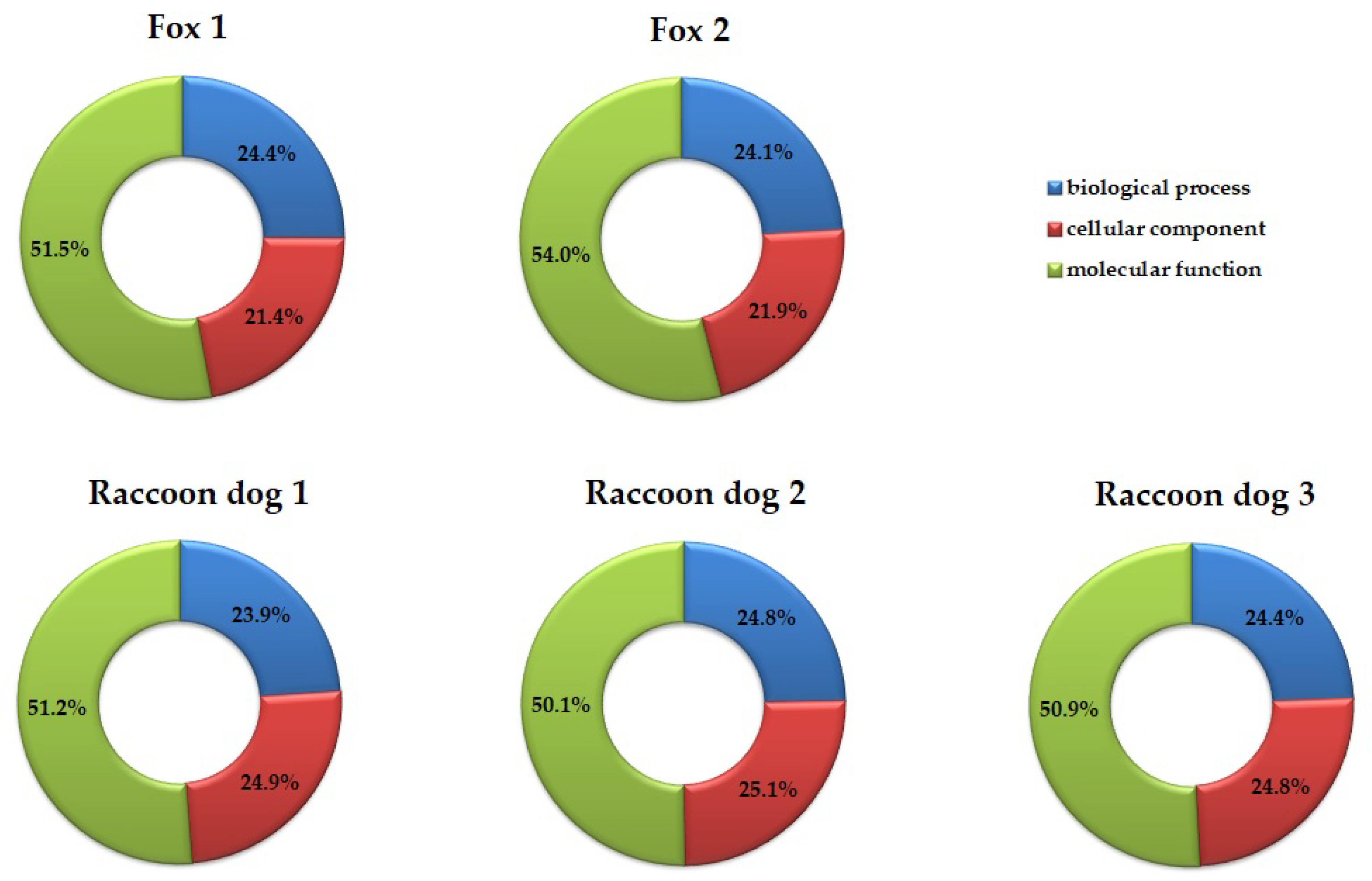
2.4. Gene Ontology (GO) Analysis
3. Discussion
4. Materials and Methods
4.1. Collection of Material and Research Scheme
4.2. ELISA
4.3. Preparation of Crude Antigens from Muscle Larvae of T. britovi
4.4. SDS–Polyacrylamide Gel Electrophoresis (SDS–PAGE) and Immunoblot
4.5. Proteomic Analyses—Mass Spectrometry (MS) and Protein Identification
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gottstein, B.; Pozio, E.; Nöckler, K. Epidemiology, diagnosis, treatment, and control of trichinellosis. Clin. Microbiol. Rev. 2009, 22, 127–145. [Google Scholar] [CrossRef] [Green Version]
- Blaga, R.; Gherman, C.; Cozma, V.; Zocevic, A.; Pozio, E.; Boireau, P. Trichinella species circulating among wild and domestic animals in Romania. Vet. Parasitol. 2009, 159, 218–221. [Google Scholar] [CrossRef]
- Pozio, E.; Rinaldi, L.; Marucci, G.; Musella, V.; Galati, F.; Cringoli, G.; Boireau, P.; La Rosa, G. Hosts and habitats of Trichinella spiralis and Trichinella britovi in Europe. Int. J. Parasitol. 2009, 39, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Pozio, E.; Hoberg, E.; La Rosa, G.; Zarlenga, D.S. Molecular taxonomy, phylogeny and biogeography of nematodes belonging to the Trichinella genus. Infect. Genet. Evol. 2009, 9, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Chmurzyńska, E.; Różycki, M.; Bilska-Zając, E.; Nöckler, K.; Mayer-Scholl, A.; Pozio, E.; Cencek, T.; Karamon, J. Trichinella nativa in red foxes (Vulpes vulpes) of Germany and Poland: Possible different origins. Vet. Parasitol. 2013, 198, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Bień, J.; Moskwa, B.; Goździk, K.; Cybulska, A.; Kornacka, A.; Welc, M.; Popiołek, M.; Cabaj, W. The occurrence of nematodes of the genus Trichinella in wolves (Canis lupus) from the Bieszczady Mountains and Augustowska Forest in Poland. Vet. Parasitol. 2016, 231, 115–117. [Google Scholar] [CrossRef]
- Deksne, G.; Segliņa, Z.; Jahundoviča, I.; Esīte, Z.; Bakasejevs, E.; Bagrade, G.; Keidāne, D.; Interisano, M.; Marucci, G.; Tonanzi, D.; et al. High prevalence of Trichinella spp. in sylvatic carnivore mammals of Latvia. Vet. Parasitol. 2016, 231, 118–123. [Google Scholar] [CrossRef]
- Kirjušina, M.; Bakasejevs, E.; Pezzotti, P.; Pozio, E. Trichinella britovi biomass in naturally infected pine martens (Martes martes) of Latvia. Vet. Parasitol. 2016, 231, 110–114. [Google Scholar] [CrossRef]
- Dmitric, M.; Vidanovic, D.; Vaskovic, N.; Matovic, K.; Sekler, M.; Debeljak, Z.; Karabasil, N. Trichinella infections in red foxes (Vulpes vulpes) and golden jackals (Canis aureus) in six districts of Serbia. J. Zoo Wildl. Med. 2017, 48, 703–707. [Google Scholar] [CrossRef]
- Kärssin, A.; Häkkinen, L.; Niin, E.; Peik, K.; Vilem, A.; Jokelainen, P.; Lassen, B. Trichinella spp. biomass has increased in raccoon dogs (Nyctereutes procyonoides) and red foxes (Vulpes vulpes) in Estonia. Parasite Vectors 2017, 10, 609. [Google Scholar] [CrossRef]
- Cybulska, A.; Skopek, R.; Kornacka, A.; Popiołek, M.; Piróg, A.; Laskowski, Z.; Moskwa, B. First detection of Trichinella pseudospiralis infection in raccoon (Procyon lotor) in Central Europe. Vet. Parasitol. 2018, 254, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Cybulska, A.; Kornacka, A.; Moskwa, B. The occurrence and muscle distribution of Trichinella britovi in raccoon dogs (Nyctereutes procyonoides) in wildlife in the Głęboki Bród Forest District, Poland. Int. J. Parasitol. Parasites Wildl. 2019, 9, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Gherman, C.M.; Boros, Z.; Băieș, M.H.; Cozma-Petruț, A.; Cozma, V. A review of Trichinella species infection in wild animals in Romania. Food Waterborne Parasitol. 2022, 28, e00178. [Google Scholar] [CrossRef] [PubMed]
- Grzelak, S.; Moskwa, B.; Bień, J. Trichinella britovi muscle larvae and adult worms: Stage-specific and common antigens detected by two-dimensional gel electrophoresis-based immunoblotting. Parasites Vectors 2018, 11, 584. [Google Scholar] [CrossRef]
- Miller, I.; Wait, R.; Sipos, W.; Gemeiner, M. A proteomic reference map for pig serum proteins as a prerequisite for diagnostic applications. Res. Vet. Sci. 2009, 86, 362–367. [Google Scholar] [CrossRef]
- Gondek, M.; Herosimczyk, A.; Knysz, P.; Ożgo, M.; Lepczyński, A.; Szkucik, K. Comparative Proteomic Analysis of Serum from Pigs Experimentally Infected with Trichinella spiralis, Trichinella britovi, and Trichinella pseudospiralis. Pathogens 2020, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Wallander, C.; Frössling, J.; Vågsholm, I.; Burrells, A.; Lundén, A. “Meat juice” is not a homogeneous serological matrix. Foodborne Pathog. Dis. 2015, 12, 280–288. [Google Scholar] [CrossRef]
- Berger-Schoch, A.E.; Bernet, D.; Doherr, M.G.; Gottstein, B.; Frey, C.F. Toxoplasma gondii in Switzerland: A serosurvey based on meat juice analysis of slaughtered pigs, wild boar, sheep and cattle. Zoonoses Public Health 2011, 58, 472–478. [Google Scholar] [CrossRef]
- Kornacka, A.; Cybulska, A.; Bień, J.; Goździk, K.; Moskwa, B. The usefulness of direct agglutination test, enzyme-linked immunosorbent assay and polymerase chain reaction for the detection of Toxoplasma gondii in wild animals. Vet. Parasitol. 2016, 228, 85–89. [Google Scholar] [CrossRef]
- Kornacka, A.; Cybulska, A.; Popiołek, M.; Kuśmierek, N.; Moskwa, B. Survey of Toxoplasma gondii and Neospora caninum in raccoons (Procyon lotor) from the Czech Republic, Germany and Poland. Vet. Parasitol. 2018, 262, 47–50. [Google Scholar] [CrossRef]
- Vico, J.P.; Mainar-Jaime, R.C. The use of meat juice or blood serum for the diagnosis of Salmonella infection in pigs and its possible implications on Salmonella control programs. J. Vet. Diagn. Investig. 2011, 23, 528–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szulowski, K.; Pilaszek, J.; Iwaniak, W. Application of meat juice in diagnosis of brucellosis in hares and wild boars by ELISA. Bull. Vet. Inst. Pulawy. 2000, 44, 45–52. [Google Scholar]
- Yonemitsu, K.; Minami, S.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Maeda, K. Detection of anti-viral antibodies from meat juice of wild boars. J. Vet. Med. Sci. 2019, 81, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Cybulska, A.; Kornacka, A.; Popiołek, M.; Bień-Kalinowska, J.; Moskwa, B. Use of meat juice from racoons (Procyon lotor) collected from Central Europe for immunological detection of Trichinella spp. Vet. Parasitol. 2021, 297, 109066. [Google Scholar] [CrossRef] [PubMed]
- Winter, M.; Abate, S.D.; Pasqualetti, M.I.; Fariña, F.A.; Ercole, M.E.; Pardini, L.; Moré, G.; Venturini, M.C.; Perera, N.; Corominas, M.J.; et al. Toxoplasma gondii and Trichinella infections in wild boars (Sus scrofa) from Northeastern Patagonia, Argentina. Prev. Vet. Med. 2019, 168, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Kärssin, A.; Velström, K.; Gómez-Morales, M.A.; Saar, T.; Jokelainen, P.; Lassen, B. Cross-Sectional Study of Anti-Trichinella Antibody Prevalence in Domestic Pigs and Hunted Wild Boars in Estonia. Vector Borne Zoonotic Dis. 2016, 16, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Morales, M.A.; Ludovisi, A.; Amati, M.; Bandino, E.; Capelli, G.; Corrias, F.; Gelmini, L.; Nardi, A.; Sacchi, C.; Cherchi, S.; et al. Indirect versus direct detection methods of Trichinella spp. infection in wild boar (Sus scrofa). Parasite Vectors 2014, 7, 171. [Google Scholar] [CrossRef] [Green Version]
- Nöckler, K.; Hamidi, A.; Fries, R.; Heidrich, J.; Beck, R.; Marinculic, A. Influence of methods for Trichinella detection in pigs from endemic and non-endemic European region. J. Vet. Med. B Infect Dis. Vet. Public Health 2004, 51, 297–301. [Google Scholar] [CrossRef]
- Bessi, C.; Ercole, M.E.; Fariña, F.A.; Ribicich, M.M.; Montalvo, F.; Acerbo, M.; Krivokapich, S.J.; Pasqualetti, M.I. Study of Trichinella patagoniensis in wild boars. Vet. Parasitol. 2021, 297, 109166. [Google Scholar] [CrossRef]
- Grzelak, S.; Stachyra, A.; Bień-Kalinowska, J. The first analysis of Trichinella spiralis and Trichinella britovi adult worm excretory-secretory proteins by two-dimensional electrophoresis coupled with LC-MS/MS. Vet. Parasitol. 2021, 297, 109096. [Google Scholar] [CrossRef]
- Cybulska, A.; Kornacka, A.; Skopek, R.; Moskwa, B. Trichinella britovi infection and muscle distribution in free-living martens (Martes spp.) from the Głęboki Bród Forest District, Poland. Int. J. Parasitol. Parasites Wildl. 2020, 12, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Antolová, D.; Fecková, M.; Valentová, D.; Hurníková, Z.; Miklisová, D.; Avdičová, M.; Halánová, M. Trichinellosis in Slovakia–epidemiological situation in humans and animals (2009-2018). Ann. Agric. Environ. Med. 2020, 27, 361–367. [Google Scholar] [CrossRef]
- Rostami, A.; Gamble, H.R.; Dupouy-Camet, J.; Khazan, H.; Bruschi, F. Meat sources of infection for outbreaks of human trichinellosis. Food Microbiol. 2017, 64, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Turiac, I.A.; Cappelli, M.G.; Olivieri, R.; Angelillis, R.; Martinelli, D.; Prato, R.; Fortunato, F. Trichinellosis outbreak due to wild boar meat consumption in southern Italy. Parasite Vectors 2017, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavic, S.; Andric, A.; Sofronic-Milosavljevic, L.J.; Gnjatovic, M.; Mitić, I.; Vasilev, S.; Sparic, R.; Pavic, A. Trichinella britovi outbreak: Epidemiological, clinical, and biological features. Med. Mal. Infect. 2020, 50, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Bień, J.; Näreaho, A.; Varmanen, P.; Goździk, K.; Moskwa, B.; Cabaj, W.; Nyman, T.A.; Savijoki, K. Comparative analysis of excretory-secretory antigens of Trichinella spiralis and Trichinella britovi muscle larvae by two-dimensional difference gel electrophoresis and immunoblotting. Proteome Sci. 2012, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Gondek, M.; Knysz, P.; Pomorska-Mól, M.; Ziomek, M.; Bień-Kalinowska, J. Acute phase protein pattern and antibody response in pigs experimentally infected with a moderate dose of Trichinella spiralis, T. britovi, and T. pseudospiralis. Vet. Parasitol. 2020, 288, 109277. [Google Scholar] [CrossRef]
- Grzelak, S.; Stachyra, A.; Moskwa, B.; Bień-Kalinowska, J. Exploiting the potential of 2D DIGE and 2DE immunoblotting for comparative analysis of crude extract of Trichinella britovi and Trichinella spiralis muscle larvae proteomes. Vet. Parasitol. 2021, 289, 109323. [Google Scholar] [CrossRef]
- Gondek, M.; Grzelak, S.; Pyz-Łukasik, R.; Knysz, P.; Ziomek, M.; Bień-Kalinowska, J. Insight into Trichinella britovi Infection in Pigs: Effect of Various Infectious Doses on Larvae Density and Spatial Larvae Distribution in Carcasses and Comparison of the Detection of Anti-T. britovi IgG of Three Different Commercial ELISA Tests and Immunoblot Assay. Pathogens 2022, 11, 735. [Google Scholar]
- Sukura, A.; Näreaho, A.; Mikkonen, T.; Niemi, M.; Oivanen, L. Trichinella nativa and T. spiralis induce distinguishable histopathologic and humoral responses in the raccoon dog (Nyctereutes procyonoides). Vet. Pathol. 2002, 39, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Davidson, R.K.; Ørpetveit, I.; Møller, L.; Kapel, C.M. Serological detection of anti-Trichinella antibodies in wild foxes and experimentally infected farmed foxes in Norway. Vet. Parasitol. 2009, 163, 93–100. [Google Scholar] [CrossRef]
- Gómez-Morales, M.A.; Selmi, M.; Ludovisi, A.; Amati, M.; Fiorentino, E.; Breviglieri, L.; Poglayen, G.; Pozio, E. Hunting dogs as sentinel animals for monitoring infections with Trichinella spp. in wildlife. Parasite Vectors 2016, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Miterpáková, M.; Antolová, D.; Hurníková, Z.; Březinová, N.; Čabanová, V.; Reiterová, K. Seroprevalence of Trichinella infections in domestic dogs from Slovakia. J. Helminthol. 2017, 91, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Grzybek, M.; Cybulska, A.; Tołkacz, K.; Alsarraf, M.; Behnke-Borowczyk, J.; Szczepaniak, K.; Strachecka, A.; Paleolog, J.; Moskwa, B.; Behnke, J.M.; et al. Seroprevalence of Trichinella spp. infection in bank voles (Myodes glareolus)–A long term study. Int. J. Parasitol. Parasites Wildl. 2019, 9, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Boros, Z.; Vallée, I.; Panait, L.C.; Gherman, C.M.; Chevillot, A.; Boireau, P.; Cozma, V. Seroprevalance of Trichinella Spp. in Wild Boars (Sus Scrofa) from Bihor County, Western Romania. Helminthologia 2020, 57, 235–240. [Google Scholar] [CrossRef]
- Pilfold, N.W.; Richardson, E.S.; Ellis, J.; Jenkins, E.; Scandrett, W.B.; Hernández-Ortiz, A.; Buhler, K.; McGeachy, D.; Al-Adhami, B.; Konecsni, K.; et al. Long-term increases in pathogen seroprevalence in polar bears (Ursus maritimus) influenced by climate change. Glob. Chang. Biol. 2021, 27, 4481–4497. [Google Scholar] [CrossRef]
- Gondek, M.; Bień, J.; Nowakowski, Z. Use of ELISA and Western blot for serological detection of antibodies to E-S antigens of Trichinella spiralis muscle larvae in sera of swine experimentally infected with Trichinella spiralis. Vet. Immunol. Immunopathol. 2018, 203, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Grzelak, S.; Stachyra, A.; Stefaniak, J.; Mrówka, K.; Moskwa, B.; Bień-Kalinowska, J. Immunoproteomic analysis of Trichinella spiralis and Trichinella britovi excretory-secretory muscle larvae proteins recognized by sera from humans infected with Trichinella. PLoS ONE 2020, 15, e0241918. [Google Scholar]
- Wang, Z.Q.; Liu, R.D.; Sun, G.G.; Song, Y.Y.; Jiang, P.; Zhang, X.; Cui, J. Proteomic Analysis of Trichinella spiralis Adult Worm Excretory-Secretory Proteins Recognized by Sera of Patients with Early Trichinellosis. Front. Microbiol. 2017, 8, 986. [Google Scholar] [CrossRef] [Green Version]
- Bień, J.; Cabaj, W.; Moskwa, B. Recognition of antigens of three different stages of the Trichinella spiralis by antibodies from pigs infected with T. spiralis. Exp. Parasitol. 2013, 134, 129–137. [Google Scholar] [CrossRef]
- Wang, L.; Cui, J.; Hu, D.D.; Liu, R.D.; Wang, Z.Q. Identification of early diagnostic antigens from major excretory-secretory proteins of Trichinella spiralis muscle larvae using immunoproteomics. Parasite Vectors 2014, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Liu, R.D.; Wang, L.; Zhang, X.; Jiang, P.; Liu, M.Y.; Wang, Z.Q. Proteomic analysis of surface proteins of Trichinella spiralis muscle larvae by two-dimensional gel electrophoresis and mass spectrometry. Parasite Vectors 2013, 6, 355. [Google Scholar] [CrossRef] [PubMed]
- Thawornkuno, C.; Nogrado, K.; Adisakwattana, P.; Thiangtrongjit, T.; Reamtong, O. Identification and profiling of Trichinella spiralis circulating antigens and proteins in sera of mice with trichinellosis. PLoS ONE 2022, 17, e0265013. [Google Scholar] [CrossRef] [PubMed]
- Troiano, G.; Nante, N. Human Trichinellosis in Italy: An epidemiological review since 1989. J. Prev. Med. Hyg. 2019, 60, E71–E75. [Google Scholar] [PubMed]
- Krivokapich, S.J.; Gatti, G.M.; Gonzalez Prous, C.L.; Degese, M.F.; Arbusti, P.A.; Ayesa, G.E.; Bello, G.V.; Salomón, M.C. Detection of Trichinella britovi in pork sausage suspected to be implicated in a human outbreak in Mendoza, Argentina. Parasitol. Int. 2019, 71, 53–55. [Google Scholar] [CrossRef]
- Dimzas, D.; Diakou, A.; Koutras, C.; Gómez Morales, M.A.; Psalla, D.; Keryttopoulos, P.; Deligianni, D.; Kontotasios, K.; Pozio, E. Human trichinellosis caused by Trichinella britovi in Greece, and literature review. J. Helminthol. 2019, 94, e33. [Google Scholar] [CrossRef]
- Stroffolini, G.; Rossi, L.; Lupia, T.; Faraoni, S.; Paltrinieri, G.; Lipani, F.; Calcagno, A.; Bonora, S.; Di Perri, G.; Calleri, G. Trichinella britovi outbreak in Piedmont, North-West Italy, 2019-2020: Clinical and epidemiological insights in the one health perspective. Travel Med. Infect. Dis. 2022, 47, 102308. [Google Scholar] [CrossRef]
- Liao, C.; Liu, M.; Bai, X.; Liu, P.; Wang, X.; Li, T.; Tang, B.; Gao, H.; Sun, Q.; Liu, X.; et al. Characterisation of a plancitoxin-1-like DNase II gene in Trichinella spiralis. PLoS Negl. Trop. Dis. 2014, 8, e3097. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Yue, X.; Han, Y.; Jiang, P.; Yang, F.; Lei, J.J.; Liu, R.D.; Zhang, X.; Wang, Z.Q.; Cui, J. Characterization of Two Trichinella spiralis Adult-Specific DNase II and Their Capacity to Induce Protective Immunity. Front. Microbiol. 2018, 9, 2504. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Stamenović, D. Contribution of intermediate filaments to cell stiffness, stiffening, and growth. Am. J. Physiol. Cell Physiol. 2000, 279, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yang, J.; Wei, J.; Yang, Y.; Chen, X.; Zhao, X.; Gu, Y.; Cui, S.; Zhu, X. Trichinella spiralis paramyosin binds to C8 and C9 and protects the tissue-dwelling nematode from being attacked by host complement. PLoS Negl. Trop. Dis. 2011, 5, e1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gretes, M.C.; Poole, L.B.; Karplus, P.A. Peroxiredoxins in parasites. Antioxid. Redox Signal 2012, 17, 608–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Liu, M.; Wang, L.; Yu, S.; Shi, H.; Boireau, P.; Cozma, V.; Wu, X.; Liu, X. Characterisation of a high-frequency gene encoding a strongly antigenic cystatin-like protein from Trichinella spiralis at its early invasion stage. Parasite Vectors 2015, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Stachyra, A.; Zawistowska-Deniziak, A.; Basałaj, K.; Grzelak, S.; Gondek, M.; Bień –Kalinowska, J. The Immunological Properties of Recombinant Multi-Cystatin-Like Domain Protein from Trichinella Britovi Produced in Yeast. Front. Immunol. 2019, 10, 2420. [Google Scholar] [CrossRef] [Green Version]
- Wakelinn, D. Immunity to Parasites: How Animals Control Parasitic Infections, London, UK, 1st ed.; Edoward Arnold: Baltimore, MD, USA, 1984. [Google Scholar]
- Gómez-Morales, M.A.; Ludovisi, A.; Amati, M.; Cherchi, S.; Tonanzi, D.; Pozio, E. Differentiation of Trichinella species (Trichinella spiralis/Trichinella britovi versus Trichinella pseudospiralis) using western blot. Parasite Vectors 2018, 11, 631. [Google Scholar] [CrossRef] [Green Version]
- Pozio, E.; Varese, P.; Morales, M.A.; Croppo, G.P.; Pelliccia, D.; Bruschi, F. Comparison of human trichinellosis caused by Trichinella spiralis and by Trichinella britovi. Am. J. Trop. Med. Hyg. 1993, 48, 568–575. [Google Scholar] [CrossRef]
- European Commission. Regulation (EC) No. 2075/2005 of the European Parliament and of the Council of 5 December 2005 laying down specific rules on official controls for Trichinella in meat. Off. J. Eur. Union 2005, L338, 60–82. [Google Scholar]
- Zarlenga, D.S.; Chute, M.B.; Martin, A.; Kapel, C.M. A multiplex PCR for unequivocal differentiation of all encapsulated and non-encapsulated genotypes of Trichinella. Int. J. Parasitol. 1999, 29, 1859–1867. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Protein | Access to Genbank | Protein | Access to Genbank |
|---|---|---|---|
| 1,5-anhydro-D-fructose reductase | KRY43899.1 | Lysosomal aspartic protease, partial | KRY49432.1 |
| 2,3-bisphosphoglycerate-independent phosphoglycerate mutase | KRY48325.1 | Malate dehydrogenase, cytoplasmic | KRY57329.1 |
| 26S protease regulatory subunit 4, partial | KRY52921.1 | Mannose-6-phosphate isomerase, partial | KRY55073.1 |
| 26S proteasome non-ATPase regulatory subunit 6, partial | KRY50659.1 | Mediator of RNA polymerase II transcription subunit 23 | KRY54660.1 |
| 32 kDa beta-galactoside-binding lectin | KRY59236.1 | Medium-chain specific acyl-CoA dehydrogenase, mitochondrial, partial | KRY52707.1 |
| 3-ketoacyl-CoA thiolase, mitochondrial | KRY51064.1 | Mitochondrial succinyl-CoA ligase subunit beta-like protein | KRY60682.1 |
| 4-hydroxybutyrate coenzyme A transferase, partial | KRY56869.1 | Mitochondrial-processing peptidase subunit alpha | KRY50357.1 |
| 60 kDa heat shock protein, mitochondrial | KRY51676.1 | Mitochondrial-processing peptidase subunit beta | KRY53080.1 |
| 60S acidic ribosomal protein P0 | KRY54528.1 | Mitochondrial isocitrate dehydrogenase, partial | KRY60163.1 |
| 60S ribosomal protein L4 | KRY61057.1 | Myophilin | KRY53866.1 |
| Actin-related protein 2/3 complex subunit 2 | KRY56438.1 | NADP-dependent malic enzyme, mitochondrial, partial | KRY59664.1 |
| Adducin-related protein 1 | KRY47021.1 | Obg-like ATPase 1 | KRY54762.1 |
| Adenylosuccinate lyase | KRY47589.1 | Paramyosin, partial | KRY49321.1 |
| Adenylosuccinate synthetase, partial | KRY50054.1 | Peroxiredoxin-2, partial | KRY49990.1 |
| ADP-ribose pyrophosphatase, mitochondrial | KRY48334.1 | Phosphate carrier protein, mitochondrial, partial | KRY57035.1 |
| Alpha-L-fucosidase | KRY54685.1 | Phosphoglucomutase-1 | KRY58984.1 |
| Aspartate aminotransferase, mitochondrial, partial | KRY49869.1 | Phosphoinositide 3-kinase regulatory subunit 4, partial | KRY53638.1 |
| Aspartate--tRNA ligase, cytoplasmic | KRY49144.1 | Polyubiquitin | KRY46475.1 |
| ATP synthase subunit alpha, mitochondrial, partial | KRY49394.1 | Polyubiquitin-like protein, partial | KRY24237.1 |
| Calpain clp-1 | KRY60775.1 | Polyubiquitin-B, partial | KRY44570.1 |
| Calreticulin, partial | KRY53845.1 | Polyubiquitin-C | KRY55776.1 |
| cAMP-dependent protein kinase regulatory subunit | KRY58719.1 | Proliferation-associated protein 2G4, partial | KRY53054.1 |
| Chymotrypsin-like elastase family member 2A, partial | KRY59723.1 | Propionyl-CoA carboxylase alpha chain, mitochondrial | KRY51819.1 |
| Cleavage and polyadenylation specificity factor subunit 2 | KRY53654.1 | Propionyl-CoA carboxylase beta chain, mitochondrial, partial | KRY52668.1 |
| Cytochrome b-c1 complex subunit 2, mitochondrial, partial | KRY49815.1 | Protein arginine N-methyltransferase 1, partial | KRY51927.1 |
| Cytosol aminopeptidase | KRY54953.1 | Protein ERGIC-53, partial | KRY52865.1 |
| Deoxyribonuclease-2-alpha | KRY53116.1 | putative [pyruvate dehydrogenase (acetyl-transferring)] kinase, mitochondrial, partial | KRY57184.1 |
| Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial | KRY46167.1 | putative aconitate hydratase, mitochondrial | KRY56796.1 |
| Dihydropyrimidinase-related protein 3 | KRY48683.1 | putative aminopeptidase W07G4.4 | KRY54333.1 |
| Diphthine--ammonia ligase, partial | KRY47980.1 | putative arginine kinase F46H5.3 | KRY58392.1 |
| DnaJ-like protein dnj-20 | KRY45252.1 | putative oxidoreductase-like protein | KRY46863.1 |
| Dynein heavy chain, cytoplasmic | KRY59544.1 | putative phosphoglycerate kinase | KRY46311.1 |
| Elongation factor G, mitochondrial | KRY55228.1 | putative pyruvate dehydrogenase E1 component subunit alpha, mitochondrial | KRY58181.1 |
| Endoplasmic reticulum resident protein 44 | KRY50742.1 | Rab GDP dissociation inhibitor alpha | KRY54650.1 |
| Endoplasmin, partial | KRY54651.1 | secretion antigen precursor | CAD86782.1 |
| Eukaryotic initiation factor 4A, partial | KRY53637.1 | Serine hydroxymethyltransferase | KRY56265.1 |
| Eukaryotic initiation factor 4A-III | KRY49326.1 | Serine protease 30 | KRY58838.1 |
| Far upstream element-binding protein 2 | KRY57083.1 | Snake venom 5’-nucleotidase | KRY49427.1 |
| Fructose-bisphosphate aldolase 1, partial | KRY52953.1 | Solute carrier family 2, facilitated glucose transporter member 3 | KRY47169.1 |
| Glucose-6-phosphate isomerase | KRY50483.1 | Splicing factor 3A subunit 3 | KRY56136.1 |
| Glyceraldehyde-3-phosphate dehydrogenase 1 | KRY59635.1 | Stress-induced-phosphoprotein 1 | KRY50814.1 |
| Glycogen phosphorylase, partial | KRY47089.1 | Succinate dehydrogenase [ubiquinone] flavoprotein subunit, mitochondrial | KRY54543.1 |
| Glycogenin-1 | KRY61196.1 | Succinate-semialdehyde dehydrogenase, mitochondrial | KRY54971.1 |
| Heat shock cognate 71 kDa protein, partial | KRY58599.1 | Sulfide:quinone oxidoreductase, mitochondrial | KRY46292.1 |
| Heat shock protein 83, partial | KRY48983.1 | T-complex protein 1 subunit alpha | KRY49588.1 |
| Histone-lysine N-methyltransferase EHMT2, partial | KRY52481.1 | T-complex protein 1 subunit beta | KRY52365.1 |
| hypothetical protein T03_13827 | KRY56487.1 | T-complex protein 1 subunit epsilon | KRY50385.1 |
| hypothetical protein T03_17187 | KRY50177.1 | T-complex protein 1 subunit eta | KRY48081.1 |
| hypothetical protein T03_539 | KRY55455.1 | T-complex protein 1 subunit gamma | KRY47292.1 |
| hypothetical protein T03_7459 | KRY60639.1 | T-complex protein 1 subunit theta | KRY50910.1 |
| hypothetical protein T03_8666, partial | KRY58917.1 | Transketolase | KRY48205.1 |
| hypothetical protein T03_8694 | KRY46541.1 | Transmembrane protease serine 5 | KRY50806.1 |
| hypothetical protein T03_9489 | KRY46491.1 | Transmembrane protease serine 9 | KRY58843.1 |
| Ig-like and fibronectin type-III domain-containing protein C25G4.10 | KRY58616.1 | Trifunctional enzyme subunit beta, mitochondrial | KRY48050.1 |
| Intermediate filament protein B, partial | KRY59373.1 | Tubulin alpha-3 chain | KRY50534.1 |
| Intermediate filament protein ifa-1 | KRY45949.1 | Type I inositol 1,4,5-trisphosphate 5-phosphatase, partial | KRY50866.1 |
| Kynurenine--oxoglutarate transaminase 3 | KRY54139.1 | Ubiquitin carboxyl-terminal hydrolase 14, partial | KRY58645.1 |
| L-2-hydroxyglutarate dehydrogenase, mitochondrial | KRY56323.1 | Uncharacterized protein T03_9851 | KRY58607.1 |
| Leukocyte elastase inhibitor C, partial | KRY55578.1 |
| Protein | Animal | GenBank Accession Number | Reference |
|---|---|---|---|
| Hypothetical protein T03_17187 | fox | KRY50178.1 | this study |
| raccoon dog | this study | ||
| human | [48] | ||
| pig | [14] | ||
| Deoxyribonuclease-2-alpha | fox | KRY53318.1 | this study |
| raccoon dog | this study | ||
| human | KRX47308.1 | [48] | |
| pig | [14] | ||
| Transmembrane protease serine 9 | fox | KRY59262.1 | this study |
| raccoon dog | this study | ||
| human | KRY20911.1 | [48] | |
| human | KRY21930.1 | [48] | |
| pig | KRY58843.1 | [38] | |
| Intermediate filament protein B, partial | fox | KRY59373.1 | this study |
| raccoon dog | this study | ||
| pig | KRY11608.1 | [38] | |
| pig | [14] | ||
| Intermediate filament protein ifa-1 | fox | KRY45949.1 | this study |
| raccoon dog | this study | ||
| pig | KRY09282.1 | [38] | |
| pig | [14] | ||
| Serine protease 30 | fox | KRY58841.1 | this study |
| raccoon dog | this study | ||
| pig | KRX47705.1 | [38] | |
| pig | [14] | ||
| Hypothetical protein T03_8694 | fox | KRY46541.1 | this study |
| raccoon dog | this study | ||
| pig | [38] | ||
| Cuticlin-1 | fox | KRY58751.1 | this study |
| raccoon dog | this study | ||
| Cuticlin-1, partial | pig | KRY01407.1 | [14] |
| pig | KRY57444.1 | [38] | |
| Paramyosin, partial | fox | KRY49321.1 | this study |
| raccoon dog | this study | ||
| pig | KRY49322.1 | [38] | |
| Myosin-4, partial | fox | KRY50587.1 | this study |
| raccoon dog | this study | ||
| pig | [38] | ||
| 26S protease regulatory subunit 10B | fox | KRY59566.1 | this study |
| raccoon dog | this study | ||
| pig | [38] | ||
| Adenylosuccinate lyase | fox | KRY47589.1 | this study |
| raccoon dog | this study | ||
| pig | [38] | ||
| 32 kDa beta-galactoside-binding lectin | fox | KRY59236.1 | this study |
| raccoon dog | this study | ||
| pig | [38] | ||
| Peroxiredoxin-2, partial | fox | KRY49990.1 | this study |
| raccoon dog | this study | ||
| pig | KRX46812.1 | [14] | |
| Poly-cysteine and histidine-tailed protein, partial | raccoon dog | KRY58369.1 | this study |
| pig | KRY11984.1 | [14] | |
| Calponin-like protein OV9M, partial | fox | KRY56610.1 | this study |
| raccoon dog | this study | ||
| pig | KRX28313.1 | [14] | |
| Propionyl-CoA carboxylase alpha chain, mitochondrial | fox | KRY51819.1 | this study |
| raccoon dog | this study | ||
| pig | KRZ50222.1 | [14] | |
| 1,5-anhydro-D-fructose reductase | fox | KRY43899.1 | this study |
| raccoon dog | this study | ||
| pig | KRY07641.1 | [14] | |
| Mitochondrial-processing peptidase subunit beta | fox | KRY53080.1 | this study |
| raccoon dog | this study | ||
| Mitochondrial-processing peptidase subunit beta, partial | pig | KRZ09733.1 | [14] |
| putative histone-binding protein Caf1 | fox | KRY53239.1 | this study |
| raccoon dog | this study | ||
| pig | KRZ17128.1 | [14] | |
| Rab GDP dissociation inhibitor alpha | fox | KRY54650.1 | this study |
| raccoon dog | this study | ||
| pig | KRY13378.1 | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cybulska, A. Immunoproteomic Analysis of Trichinella britovi Proteins Recognized by IgG Antibodies from Meat Juice of Carnivores Naturally Infected with T. britovi. Pathogens 2022, 11, 1155. https://doi.org/10.3390/pathogens11101155
Cybulska A. Immunoproteomic Analysis of Trichinella britovi Proteins Recognized by IgG Antibodies from Meat Juice of Carnivores Naturally Infected with T. britovi. Pathogens. 2022; 11(10):1155. https://doi.org/10.3390/pathogens11101155
Chicago/Turabian StyleCybulska, Aleksandra. 2022. "Immunoproteomic Analysis of Trichinella britovi Proteins Recognized by IgG Antibodies from Meat Juice of Carnivores Naturally Infected with T. britovi" Pathogens 11, no. 10: 1155. https://doi.org/10.3390/pathogens11101155