
Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland


Abstract
:1. Introduction
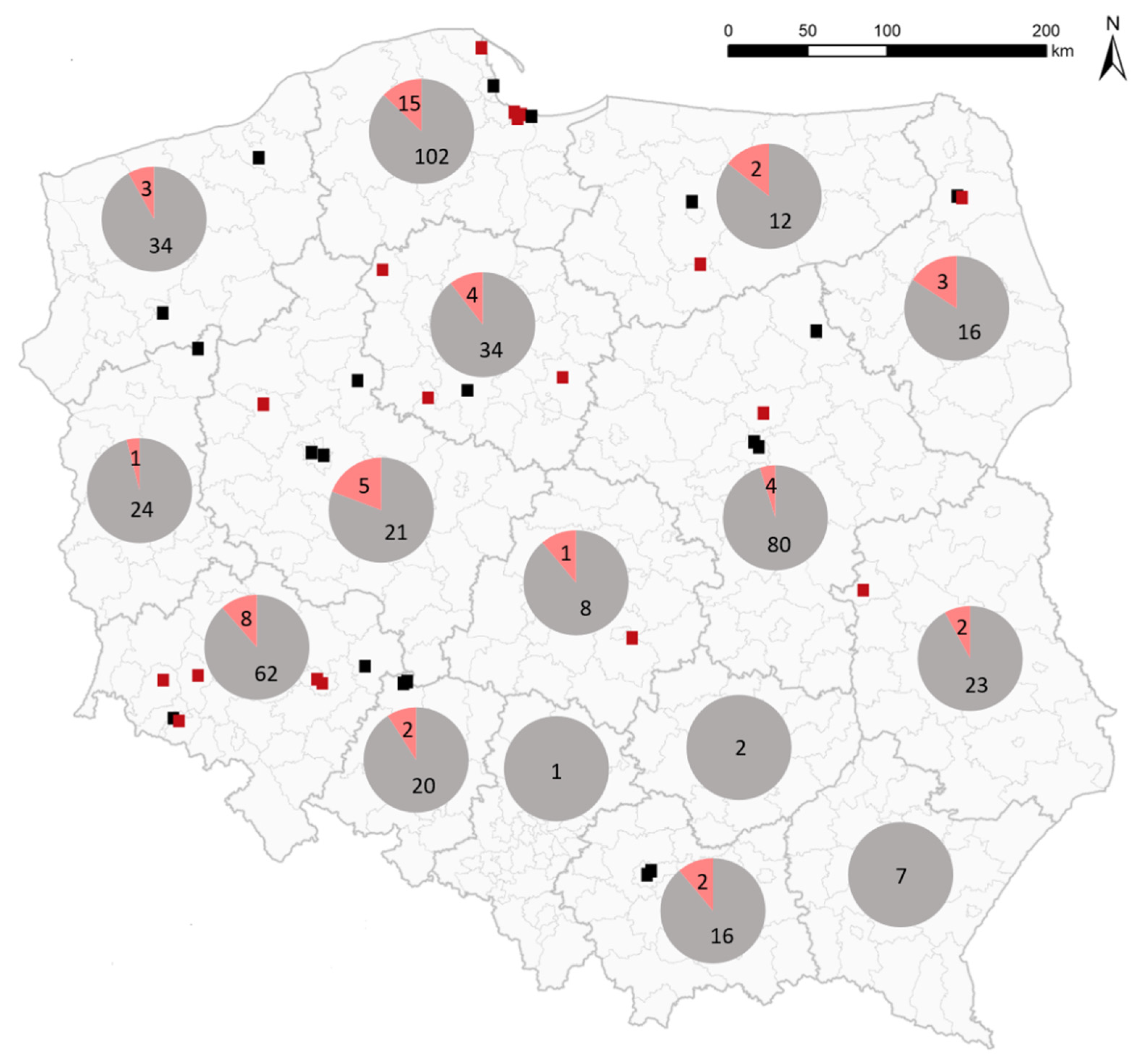
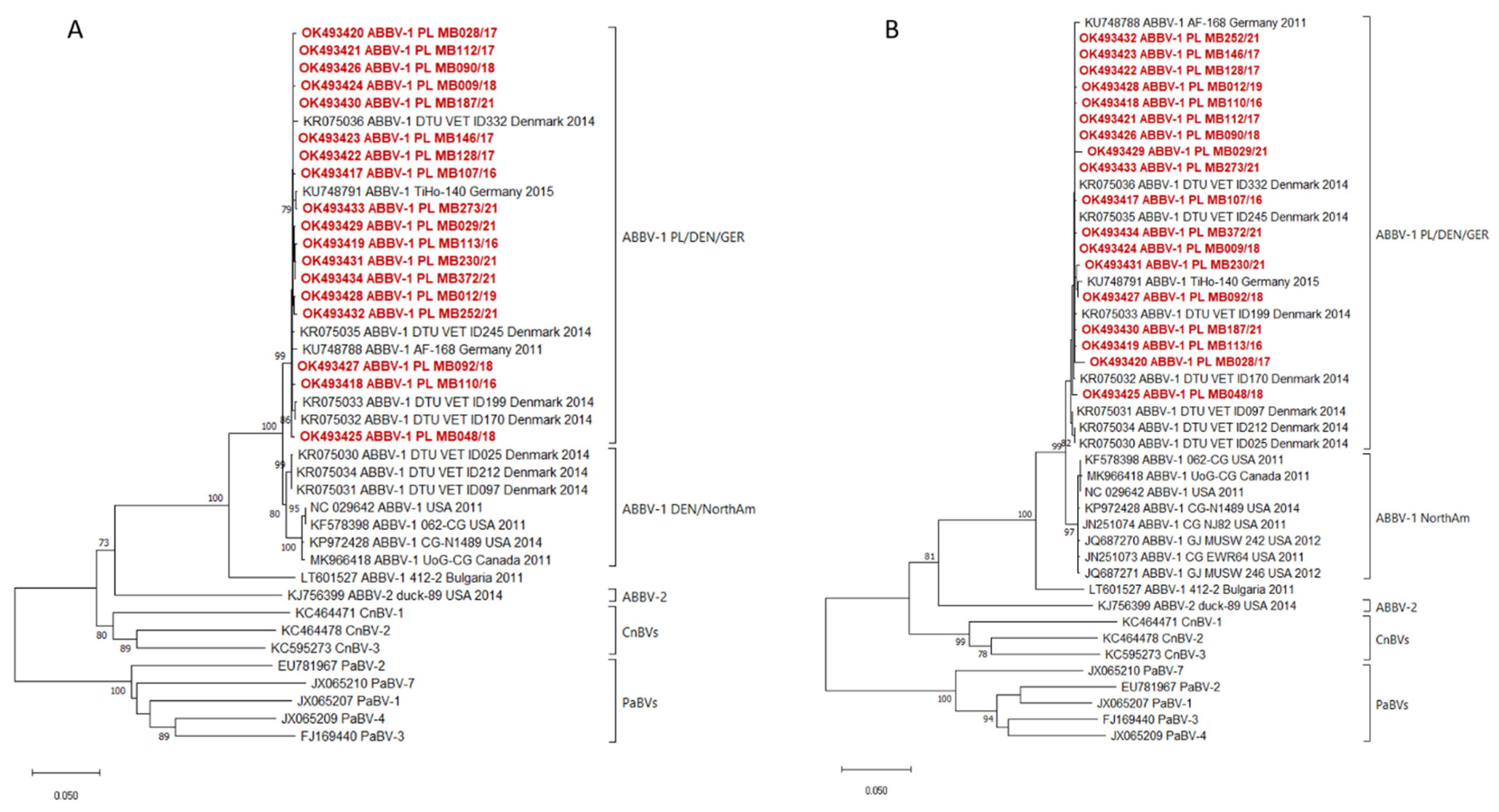
2. Results
3. Discussion
4. Materials and Methods
4.1. Samples from Wild Birds
4.2. Detection of ABBVs RNA with Real Time RT-PCR
4.3. Sequencing and Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rubbenstroth, D.; Briese, T.; Dürrwald, R.; Horie, M.; Hyndman, T.H.; Kuhn, J.H.; Nowotny, N.; Payne, S.; Stenglein, M.D.; Tomonaga, K.; et al. ICTV Virus Taxonomy Profile: Bornaviridae. J. Gen. Virol. 2021, 102, 001613. [Google Scholar] [CrossRef] [PubMed]
- Gancz, A.Y.; Clubb, S.; Shivaprasad, H.L. Advanced diagnostic approaches and current management of proventricular dilatation disease. Vet. Clin. N. Am. Exot. Anim. Pract. 2010, 13, 471–494. [Google Scholar] [CrossRef] [PubMed]
- Staeheli, P.; Rinder, M.; Kaspers, B. Avian bornavirus associated with fatal disease in psittacine birds. J. Virol. 2010, 84, 6269–6275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissenböck, H.; Sekulin, K.; Bakonyi, T.; Högler, S.; Nowotny, N. Novel avian bornavirus in a nonpsittacine species (Canary; Serinus canaria) with enteric ganglioneuritis and encephalitis. J. Virol. 2009, 83, 11367–11371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delnatte, P.; Berkvens, C.; Kummrow, M.; Smith, D.A.; Campbell, D.; Crawshaw, G.; Ojkic, D.; DeLay, J. New genotype of avian bornavirus in wild geese and trumpeter swans in Canada. Vet. Rec. 2011, 169, 108. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Covaleda, L.; Heatley, J.J.; Baroch, J.A.; Tizard, I.; Payne, S.L. Widespread avian bornavirus infection in mute swans in the Northeast United States. Vet. Med. 2012, 3, 49–52. [Google Scholar]
- Delnatte, P.; Ojkic, D.; DeLay, J.; Campbell, D.; Crawshaw, G.; Smith, D.A. Pathology and diagnosis of avian bornavirus infection in wild Canada geese (Branta canadensis), trumpeter swans (Cygnus buccinator) and mute swans (Cygnus olor) in Canada: A retrospective study. Avian. Pathol. 2013, 42, 114–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, S.; Covaleda, L.; Jianhua, G.; Swafford, S.; Baroch, J.; Ferro, P.J.; Lupiani, B.; Heatley, J.; Tizard, I. Detection and characterization of a distinct bornavirus lineage from healthy Canada geese (Branta canadensis). J. Virol. 2011, 85, 12053–12066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Tizard, I.; Baroch, J.; Shivaprasad, H.L.; Payne, S.L. Avian Bornaviruses in North American Gulls. J. Wildl. Dis. 2015, 51, 754–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, M.; Guo, J.; Tizard, I.; Jennings, S.; Shivaprasad, H.L.; Payne, S.; Ellis, J.C.; Van Wettere, A.J.; O’Brien, K.M. Aquatic Bird Bornavirus-Associated Disease in Free-Living Canada Geese (Branta canadensis) in the Northeastern USA. J. Wildl. Dis. 2017, 53, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Shivaprasad, H.L.; Rech, R.R.; Heatley, J.J.; Tizard, I.; Payne, S. Characterization of a new genotype of avian bornavirus from wild ducks. Virol. J. 2014, 11, 197. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, A.F.; Nielsen, J.B.; Hjulsager, C.K.; Chriél, M.; Smith, D.A.; Bertelsen, M.F. Aquatic Bird Bornavirus 1 in Wild Geese, Denmark. Emerg. Infect. Dis. 2015, 21, 2201–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubbenstroth, D.; Schmidt, V.; Rinder, M.; Legler, M.; Twietmeyer, S.; Schwemmer, P.; Corman, V.M. Phylogenetic Analysis Supports Horizontal Transmission as a Driving Force of the Spread of Avian Bornaviruses. PLoS ONE 2016, 11, e0160936. [Google Scholar] [CrossRef] [PubMed]
- Rubbenstroth, D.; Schlottau, K.; Schwemmle, M.; Rissland, J.; Beer, M. Human bornavirus research: Back on track! PLoS Pathog. 2019, 15, e1007873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinder, M.; Ackermann, A.; Kempf, H.; Kaspers, B.; Korbel, R.; Staeheli, P. Broad tissue and cell tropism of avian bornavirus in parrots with proventricular dilatation disease. J. Virol. 2009, 83, 5401–5407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delnatte, P.; Mak, M.; Ojkic, D.; Raghav, R.; DeLay, J.; Smith, D.A. Detection of Avian bornavirus in multiple tissues of infected psittacine birds using real-time reverse transcription polymerase chain reaction. J. Vet. Diagn. Invest. 2014, 26, 266–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delnatte, P.; Nagy, É.; Ojkic, D.; Leishman, D.; Crawshaw, G.; Elias, K.; Smith, D.A. Avian bornavirus in free-ranging waterfowl: Prevalence of antibodies and cloacal shedding of viral RNA. J. Wildl. Dis. 2014, 50, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Sigrist, B.; Geers, J.; Albini, S.; Rubbenstroth, D.; Wolfrum, N. A New Multiplex Real-Time RT-PCR for Simultaneous Detection and Differentiation of Avian Bornaviruses. Viruses 2021, 13, 1358. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Number of Tested Birds | Number of ABBV-Positive Birds |
|---|---|---|
| mute swan (Cygnus olor) | 220 | 36 |
| whooper swan (Cygnus cygnus) | 2 | 1 |
| black swan (Cygnus atratus) | 9 | 0 |
| swan (Cygnus sp.) 1 | 170 | 15 |
| mallard (Anas platyrhynchos) | 48 | 0 |
| long-tailed duck (Clangula hyemalis) | 11 | 0 |
| garganey (Spatula querquedula) | 3 | 0 |
| wild duck 2 | 31 | 0 |
| other ducks 3 | 3 | 0 |
| bean goose (Anser fabalis) | 7 | 0 |
| tundra bean goose (Anser serrirostris) | 3 | 0 |
| greylag goose (Anser anser) | 3 | 0 |
| white-fronted goose (Anser albifrons) | 1 | 0 |
| wild goose 2 | 3 | 0 |
| Total | 514 | 52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Świętoń, E.; Dziadek, K.; Śmietanka, K. Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland. Pathogens 2022, 11, 98. https://doi.org/10.3390/pathogens11010098
Świętoń E, Dziadek K, Śmietanka K. Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland. Pathogens. 2022; 11(1):98. https://doi.org/10.3390/pathogens11010098
Chicago/Turabian StyleŚwiętoń, Edyta, Kamila Dziadek, and Krzysztof Śmietanka. 2022. "Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland" Pathogens 11, no. 1: 98. https://doi.org/10.3390/pathogens11010098