
Wild Small Mammals and Ticks in Zoos—Reservoir of Agents with Zoonotic Potential?


Abstract
:1. Introduction
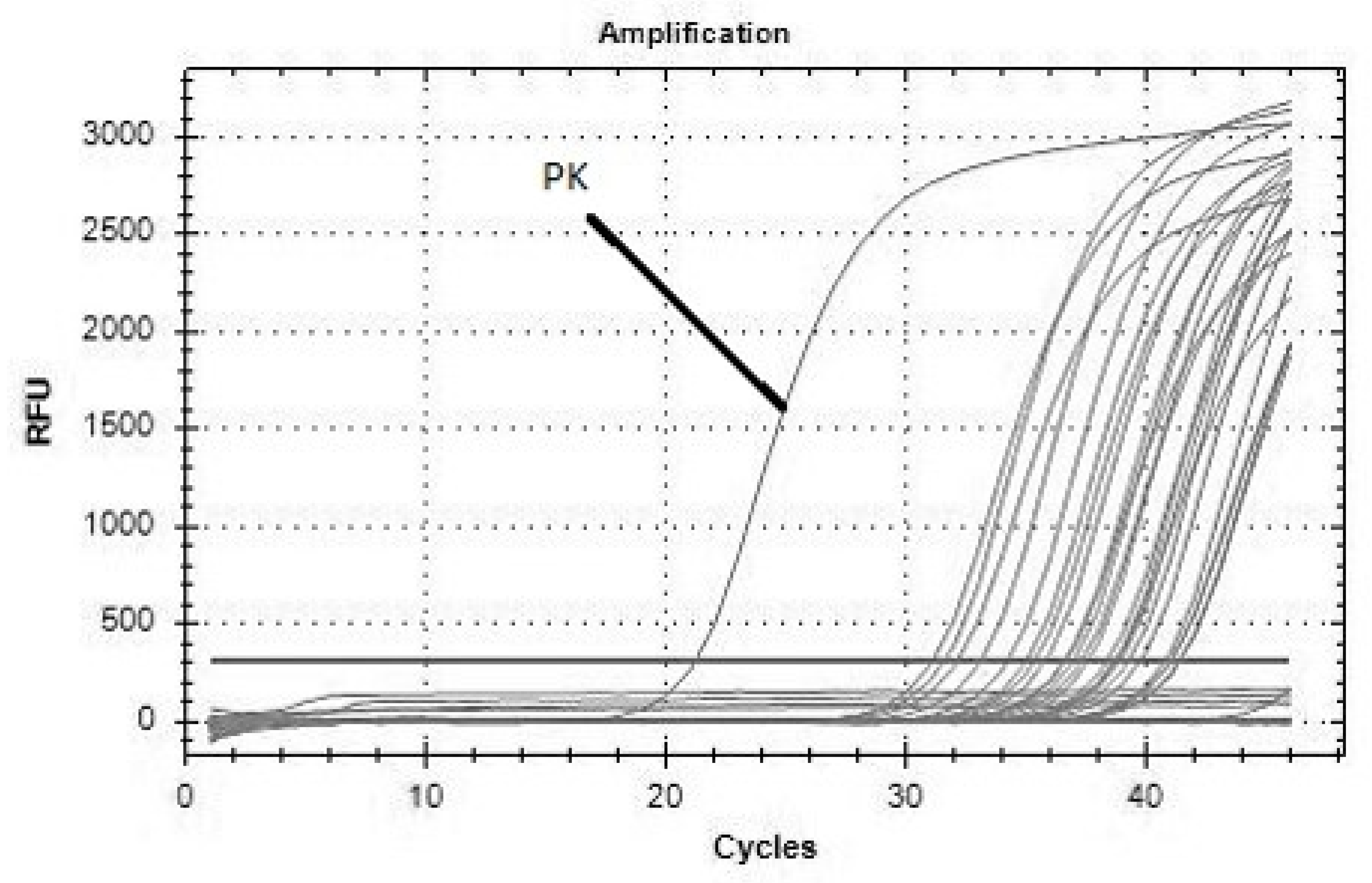
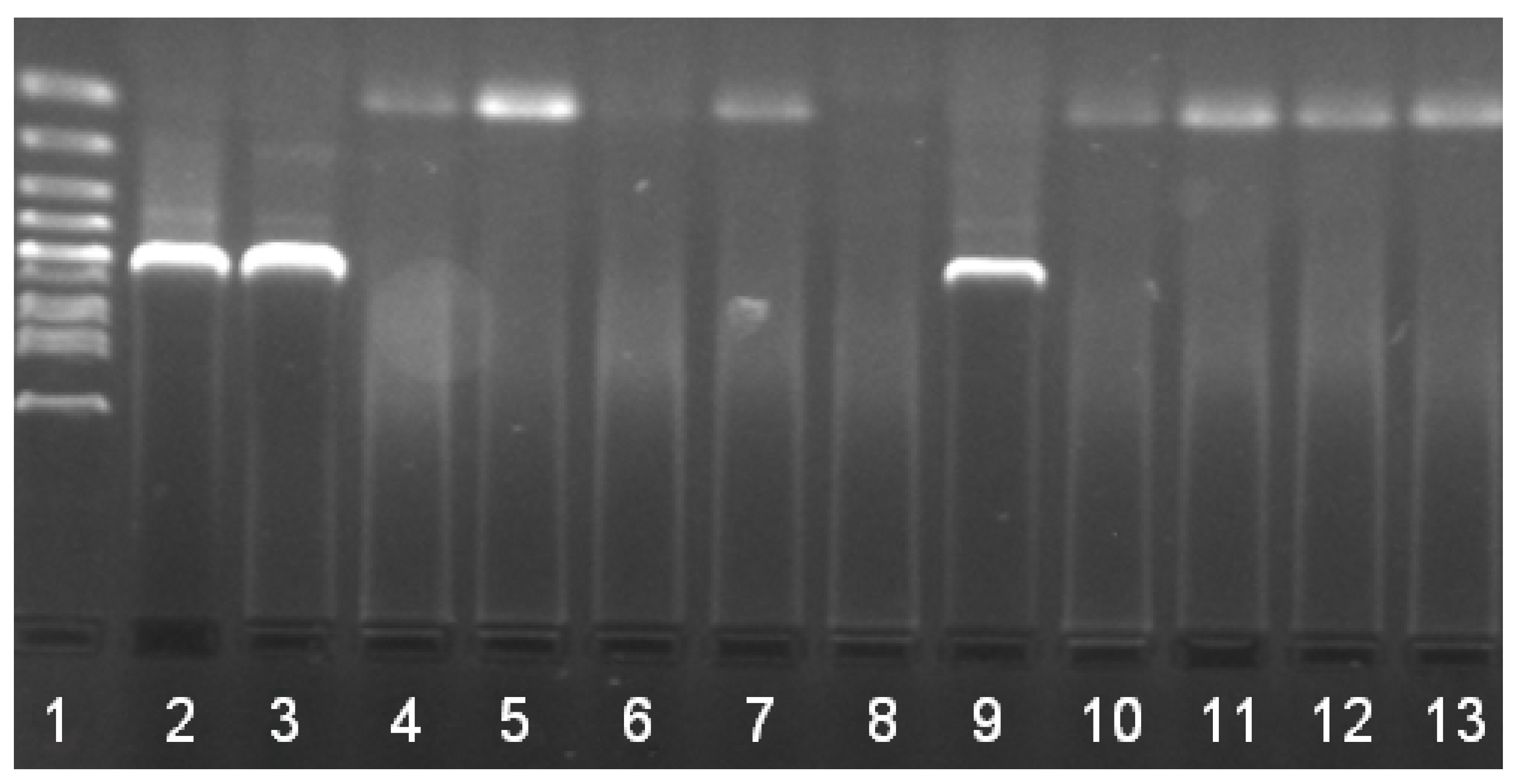
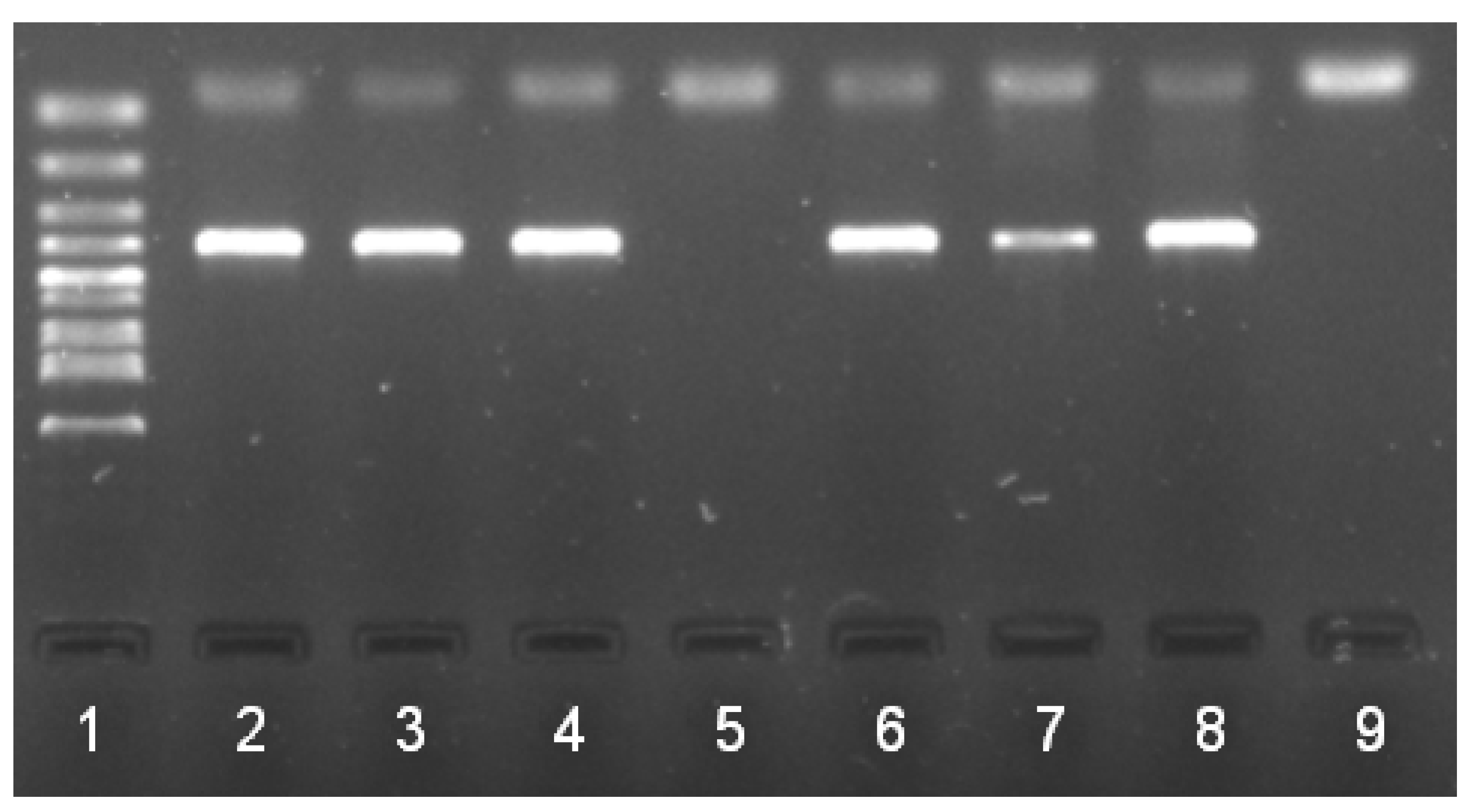
2. Results
3. Discussion
4. Materials and Methods
4.1. Sampling (Wild Small Mammals, Ticks)
4.2. Serology
4.3. Molecular Methods
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gürtler, L.; Bauerfeind, U.; Blümel, J.; Burger, R.; Drosten, C.; Gröner, A.; Heiden, M.; Hildebrandt, M.; Jansen, B.; Offergeld, R.; et al. Coxiella burnetiid—pathogenic agent of Q (Query) fever. Transfus. Med. Hemoth. 2014, 41, 60–72. [Google Scholar]
- Maurin, M.; Raoult, D. Q fever. Clin. Microbiol. Rev. 1999, 12, 518–553. [Google Scholar] [CrossRef] [Green Version]
- Hennebique, A.; Boisset, S.; Maurin, M. Tularemia as a waterborne disease: A review. Emerg. Microbes Infect. 2019, 8, 1027–1042. [Google Scholar] [CrossRef]
- Britannica, The Editors of Encyclopaedia. Tularemia. Encyclopedia Britannica, 6 August 2020. Available online: https://www.britannica.com/science/tularemia (accessed on 3 June 2021).
- Bertasio, C.; Papetti, A.; Scaltriti, E.; Tagliabue, S.; D’Incau, M.; Boniotti, M.B. Serological survey and molecular typing reveal new Leptospira serogroup pomona strains among pigs of Northern Italy. Pathogens 2020, 9, 332. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Peru-United States Leptospirosis Consortium. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Grennan, D. Leptospirosis. JAMA. 2019, 321, 812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawczyk, A.I.; van Duijvendijk, G.L.A.; Swart, A.; Heylen, D.; Jaarsma, R.I.; Jacobs, F.H.H.; Fonville, M.; Sprong, H.; Takken, W. Effect of rodent density on tick and tick-borne pathogen populations: Consequences for infectious disease risk. Parasites Vectors 2020, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Lernout, T.; De Regge, N.; Tersago, K.; Fonville, M.; Suin, V.; Sprong, H. Prevalence of pathogens in ticks collected from humans through citizen science in Belgium. Parasites Vectors 2019, 12, 550. [Google Scholar] [CrossRef] [PubMed]
- Gross, L. A new view on Lyme disease: Rodents hold the key to annual risk. PLoS Biol. 2006, 4, 182. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Strle, F.; Wormser, G.P.; Hu, L.T.; Branda, J.A.; Hovius, J.W.R.; Li, X.; Mead, P.S. Lyme borreliosis. Nat. Rev. Dis. Primers. 2016, 2, 16090. [Google Scholar] [CrossRef]
- Borawski, K.; Dunaj, J.; Czupryna, P.; Pancewicz, S.; Świerzbińska, R.; Żebrowska, A.; Moniuszko-Malinowska, A. Prevalence of spotted fever group rickettsia in North-Eastern Poland. Infect. Dis. 2019, 51, 810–814. [Google Scholar] [CrossRef]
- Raoult, D.; Roux, V. Rickettsioses as paradigms of new or emerging infectious diseases. Clin Microbiol Rev. 1997, 10, 694–719. [Google Scholar] [CrossRef]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasites Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Ismail, N.; Bloch, K.C.; McBride, J.W. Human ehrlichiosis and anaplasmosis. Clin. Lab. Med. 2010, 30, 261–292. [Google Scholar] [CrossRef] [Green Version]
- Bártová, E.; Kučerová, H.L.; Žákovská, A.; Budíková, M.; Nejezchlebová, H. Coxiella burnetii and Francisella tularensis in wild small mammals from the Czech Republic. Ticks Tick-Borne Dis. 2020, 11, 101350. [Google Scholar] [CrossRef]
- Meredith, A.L.; Cleaveland, S.C.; Denwood, M.J.; Brown, J.K.; Shaw, D.J. Coxiella burnetii (Q Fever) seroprevalence in prey and predators in the United Kingdom: Evaluation of infection in wild rodents, foxes and domestic cats using a modified ELISA. Transbound. Emerg. Dis. 2015, 62, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Bielawska-Drózd, A.; Cieślik, P.; Żakowska, D.; Głowacka, P.; Wlizło-Skowronek, B.; Ziȩba, P.; Zdun, A. Detection of Coxiella burnetii and Francisella tularensis in tissues of wild-living animals and in ticks of North-West Poland. Pol. J. Microbiol. 2018, 67, 529–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorko, E.; Rimarova, K.; Pilipinec, E.; Travnicek, M. Prevalence of Coxiella burnetii antibodies in wild ruminants in Kavecany ZOO, Kosice, eastern Slovakia. Ann. Agr. Env. Med. 2009, 16, 321–324. [Google Scholar]
- Otto, P.; Chaignat, V.; Klimpel, D.; Diller, R.; Melzer, F.; Muller, W.; Tomaso, H. Serological investigation of wild boars (Sus scrofa) and red foxes (Vulpes vulpes) asindicator animals for circulation of Francisella tularensis in Germany. Vector-Borne Zoonotic Dis. 2014, 14, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossow, H.; Sissonen, S.; Koskela, K.A.; Kinnunen, P.M.; Hemmila, H.; Niemimaa, J.; Huitu, O.; Kuusi, M.; Vapalahti, O.; Henttonen, H.; et al. Detection of Francisella tularensis in voles in Finland. Vector Borne Zoonotic Dis. 2014, 14, 193–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascucci, L.; Di Domenico, M.; Dall’Acqua, F.; Sozio, G.; Cammà, C. Detection of lyme disease and Q fever agents in wild rodents in central Italy. Vector-Borne Zoonotic 2015, 15, 404–411. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.; Essbauer, S.S.; Mayer-Scholl, A.; Poppert, S.; Schmidt-Chanasit, J.; Klempa, B.; Henning, K.; Schares, G.; Groschup, M.; Spitzenberger, F.; et al. Multiple infections of rodents with zoonotic pathogens in Austria. Vector-Borne Zoonotic 2014, 14, 467–475. [Google Scholar] [CrossRef] [Green Version]
- State Health Institute Data. Available online: www.uzis.cz/en/about-us (accessed on 20 April 2021).
- Vostal, K.; Žákovská, A. Two-year study of examination of blood from wild rodents for the presence of antiborrelian antibodies. Ann. Agric. Environ. Med. 2003, 10, 203–206. [Google Scholar]
- Kybicová, K.; Kurzová, Z.; Hulínská, D. Molecular and serological evidence of Borrelia burgdorferi sensu lato in wild rodents in the Czech Republic. Vector-Borne Zoonotic 2008, 8, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Kučerová, H.L.; Žákovská, A.; Marková, J.; Bártová, E. Detection of antibodies to Borrelia burgdorferi s.l. in wild small mammals and sensitivity of PCR and cultivation. Vet. Microbiol. 2019, 230, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Štefančíková, A.; Bhide, M.; Peťko, B.; Stanko, M.; Mošanský, L.; Fričova, J.; Derdáková, M.; Trávniček, M. Anti-Borrelia antibodies in rodents: Important hosts in ecology of Lyme disease. Ann. Agric. Environ. Med. 2004, 11, 209–213. [Google Scholar] [PubMed]
- Pangrácová, L.; Derdáková, M.; Pekárik, L.; Hviščová, I.; Víchová, B.; Stanko, M.; Hlavatá, H.; Peťko, B. Ixodes ricinus abundance and its infection with the tick-borne pathogens in urban and suburban areas of Eastern Slovakia. Parasites Vectors 2013, 6, 238. [Google Scholar] [CrossRef] [Green Version]
- Širmarová, J.; Tichá, L.; Golovchenko, M.; Salát, J.; Grubhoffer, L.; Rudenko, N.; Nowotny, N.; Růžek, D. Seroprevalence of Borrelia burgdorferi sensu lato and tick-borne encephalitis virus in zoo animal species in the Czech Republic. Ticks Tick-Borne Dis. 2014, 5, 523–527. [Google Scholar] [CrossRef]
- Stoebel, K.; Schoenberg, A.; Streich, W.J. The seroepidemiology of Lyme borreliosis in zoo animals in Germany. Epidemiol. Infect. 2003, 131, 975–983. [Google Scholar] [CrossRef]
- Stanko, M.; Prokopcáková, H.; Fricová, J.; Pet’ko, B. Leptospira antibodies in small mammals in Eastern Slovakia. Vet. Med. 1996, 41, 373–377. [Google Scholar]
- Boey, K.; Shiokawa, K.; Rajeev, S. Leptospira infection in rats: A literature review of global prevalence and distribution. PLoS Negl. Trop. Dis. 2019, 13, e0007499. [Google Scholar] [CrossRef]
- Beest, J.T.; Cushing, A.; McClean, M.; Hsu, W.; Bildfell, R. Disease surveillance of California Ground Squirrels (Spermophilus beecheyi) in a Drive-through Zoo in Oregon, USA. J. Wildl. Dis. 2017, 53, 667–670. [Google Scholar] [CrossRef] [Green Version]
- Himsworth, C.G.; Jardine, C.M.; Parsons, K.L.; Feng, A.Y.T.; Patrick, D.M. The characteristics of wild rat (Rattus spp.) populations from an inner-city neighborhood with a focus on factors critical to the understanding of rat-associated zoonoses. PLoS ONE 2014. [Google Scholar] [CrossRef]
- Derdáková, M.; Beati, L.; Pet’ko, B.; Stanko, M.; Fish, D. Genetic variability within Borrelia burgdorferi sensu lato genospecies established by PCR-single-strand conformation polymorphism analysis of the rrfA-rrlB intergenic spacer in Ixodes ricinus ticks from the Czech Republic. Appl. Environ. Microb. 2003, 69, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Hönig, V.; Svec, P.; Halas, P.; Vavruskova, Z.; Tykalova, H.; Kilian, P.; Vetiskova, V.; Dornakova, V.; Sterbova, J.; Simonova, Z.; et al. Ticks and tick-borne pathogens in South Bohemia (Czech Republic)—Spatial variability in Ixodes ricinus abundance, Borrelia burgdorferi and tick-borne encephalitis virus prevalence. Ticks Tick-Borne Dis. 2015, 6, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Žákovská, A.; Nejezchlebová, H.; Bartoňková, N.; Rašovská, T.; Kučerová, H.L.; Norek, A.; Ovesná, P. Activity of the tick Ixodes ricinus monitored in a suburban park in Brno, Czech Republic, in association with the evaluation of selected repellents. J. Vector. Ecol. 2013, 38, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Venclikova, K.; Rudolf, I.; Mendel, J.; Betasova, L.; Hubalek, Z. Rickettsiae in questing Ixodes ricinus ticks in the Czech Republic. Ticks Tick-Borne Dis. 2014, 5, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Hönig, V.; Carolan, H.E.; Vavruskova, Z.; Massire, C.; Mosel, M.R.; Crowder, C.D.; Rounds, M.A.; Ecker, D.J.; Ruzek, D.; Grubhoffer, L.; et al. Broad-range survey of vector-borne pathogens and tick host identification of Ixodes ricinus from Southern Czech Republic. FEMS Microbiol. Ecol. 2017, 93, fix129. [Google Scholar] [CrossRef] [PubMed]
- Svitálková, Z.; Haruštiaková, D.; Mahríková, L.; Berthová, L.; Slovák, M.; Kocianová, E.; Kazimírová, M. Anaplasma phagocytophilum prevalence in ticks and rodents in an urban and natural habitat in South-Western Slovakia. Parasites Vectors. 2015, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žákovská, A.; Rusňáková, H.; Vostal, K. Host response to Borrelia afzelii in BALB/c mice tested by immunoblotting. Ann. Agric. Environ. Med. 2013, 20, 823–825. [Google Scholar]
- Shropshire, S.B.; Veir, J.K.; Morris, A.K.; Lappin, M.R. Evaluation of Leptospira species microscopic agglutination test in experimentally vaccinated cats and Leptospira species seropositivity in aged azotemic client-owned cats. J. Feline Med. Surg. 2016, 18, 768–772. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, M.; Traweger, A.; Lusa, L.; Stupica, D.; Maraspin, V.; Barrett, P.N.; Strle, F.; Livey, I. Quantitative detection of Borrelia burgdorferi sensu lato in erythema migrans skin lesions using internally controlled duplex real time PCR. PLoS ONE 2013, 8, e63968. [Google Scholar] [CrossRef]
- Labruna, M.B.; Whitworth, T.; Horta, M.C.; Bouyer, D.H.; McBride, J.W.; Pinter, A.; Popov, V.; Gennari, S.M.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of São Paulo, Brazil, where Brazilian spotted fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massung, R.F.; Slater, K.; Owens, J.H.; Nicholson, W.L.; Mather, T.N.; Solberg, V.B.; Olson, J.G. Nested PCR assay for detection of granulocytic Ehrlichiae. J. Clin. Microbiol. 1998, 36, 1090–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- STATISTICA (Data Analysis Software System), Version 12; Statsoft Inc.: Tulsa, OK, USA, 2013. Available online: www.statsoft.com (accessed on 6 June 2000).




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Total | Serology | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C. burnetii | F. tularensis | B. burgdorferi s.l. | Leptospira spp. | ||||||||
| Total | IgM | IgG | Total | IgM | IgG | Total | IgM | IgG | Total | ||
| Species | |||||||||||
| Apodemus flavicollis | 44 | 6/37 (16%) | 6 (16%) | 0 (0%) | 2/37 (5%) | 2 (5%) | 0 (0%) | 3/37 (8%) | 0 (0%) | 0 (0%) | 1/17 (6%) |
| Apodemus sylvaticus | 20 | 4/10 (40%) | 4 (40%) | 0 (0%) | 1/10 (10%) | 1 (10%) | 0 (0%) | 2/10 (20%) | 0 (0%) | 1 (10%) | 0/1 |
| Mus musculus | 40 | 7/40 (18%) | 5 (13%) | 2 (5%) | 1/40 (3%) | 0 (0%) | 1 (3%) | 10/40 (25%) | 2 (5%) | 6 (15%) | 2/35 (6%) |
| Myodes glareolus | 4 | 0/3 | 0 | 0 | 0/3 | 0 | 0 | 0/3 | 0 | 0 | 0/2 |
| Rattus norvegicus var. alba | 6 | 2/6 (33%) | 2 (33%) | 0 (0%) | 0/6 (0%) | 0 (0%) | 0 (0%) | 0/6 (0%) | 0 (0%) | 0 (0%) | 1/6 (17%) |
| Sorex araneus | 3 | 0/3 | 0 | 0 | 0/3 | 0 | 0 | 0/3 | 0 | 0 | 0/2 |
| Sex | |||||||||||
| Female | 58 | 8/52 (15%) | 8 (15%) | 0 (0%) | 3/52 (6%) | 2 (4%) | 1 (2%) | 7/52 (13%) | 0 (0%) | 3 (6%) | 1/38 (3%) |
| Male | 59 | 11/47 (23%) | 9 (19%) | 2 (4%) | 1/47 (2%) | 1 (2%) | 0 (0%) | 8/47 (17%) | 2 (4%) | 4 (9%) | 3/25 (12%) |
| Age | |||||||||||
| Adult | 60 | 9/51 (18%) | 7 (14%) | 2 (4%) | 2/51 (4%) | 2 (4%) | 0 (0%) | 8/51 (16%) | 2 (4%) | 4 (8%) | 0/32 (0%) |
| Juvenile | 57 | 10/48 (21%) | 10 (21%) | 0 (0%) | 2/48 (4%) | 1 (2%) | 1 (2%) | 7/48 (15%) | 0 (0%) | 3 (6%) | 4/31 (13%) |
| Year of trapping * | |||||||||||
| 2016 | 29 | 6/29 (21%) | 6 (21%) | 0 (0%) | 2/29 (7%) | 2 (7%) | 0 (0%) | 2/29 (7%) | 0 (0%) | 0 (0%) | 1/10 (10%) |
| 2017 | 35 | 7/17 (41%) | 7 (41%) | 0 (0%) | 1/17 (6%) | 1 (6%) | 0 (0%) | 3/17 (18%) | 0 (0%) | 2 (12%) | 0/0 |
| 2018 | 53 | 6/53 (11%) | 4 (8%) | 2 (4%) | 1/53 (2%) | 0 (0%) | 1 (2%) | 10/53 (19%) | 2 (4%) | 5 (9%) | 3/53 (6%) |
| Total | 117 | 19/99 (19%) | 17 (17%) | 2 (2%) | 4/99 (4%) | 3 (3%) | 1 (1%) | 15/99 (15%) | 2 (2%) | 7 (7%) | 4/63 (6%) |
| Characteristic | Total | PCR | ||
|---|---|---|---|---|
| B. burgdorferi s.l. | Rickettsia sp. | A. phagocytophilum | ||
| Life stage | ||||
| Nymph | 24 | 6/24 (25%) | 3/24 (12.5%) | 0/24 |
| Adult | 46 | 5/46 (10.9%) | 1/46 (2.2%) | 1/46 (2.2%) |
| Sex | ||||
| Female | 21 | 2/21 (9.5%) | 0/21 | 1/21 (4.8%) |
| Male | 25 | 3/25 (12%) | 1/25 (4%) | 0/25 |
| Total | 70 | 11/70 (16%) | 4/70 (6%) | 1/70 (1%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pittermannová, P.; Žákovská, A.; Váňa, P.; Marková, J.; Treml, F.; Černíková, L.; Budíková, M.; Bártová, E. Wild Small Mammals and Ticks in Zoos—Reservoir of Agents with Zoonotic Potential? Pathogens 2021, 10, 777. https://doi.org/10.3390/pathogens10060777
Pittermannová P, Žákovská A, Váňa P, Marková J, Treml F, Černíková L, Budíková M, Bártová E. Wild Small Mammals and Ticks in Zoos—Reservoir of Agents with Zoonotic Potential? Pathogens. 2021; 10(6):777. https://doi.org/10.3390/pathogens10060777
Chicago/Turabian StylePittermannová, Pavlína, Alena Žákovská, Petr Váňa, Jiřina Marková, František Treml, Lenka Černíková, Marie Budíková, and Eva Bártová. 2021. "Wild Small Mammals and Ticks in Zoos—Reservoir of Agents with Zoonotic Potential?" Pathogens 10, no. 6: 777. https://doi.org/10.3390/pathogens10060777