
The Prevalence of Virulence Determinants and Antibiotic Resistance Patterns in Methicillin—Resistant Staphylococcus aureus in a Nursing Home in Poland


Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Resistance
2.2. Toxins
2.3. Adhesins
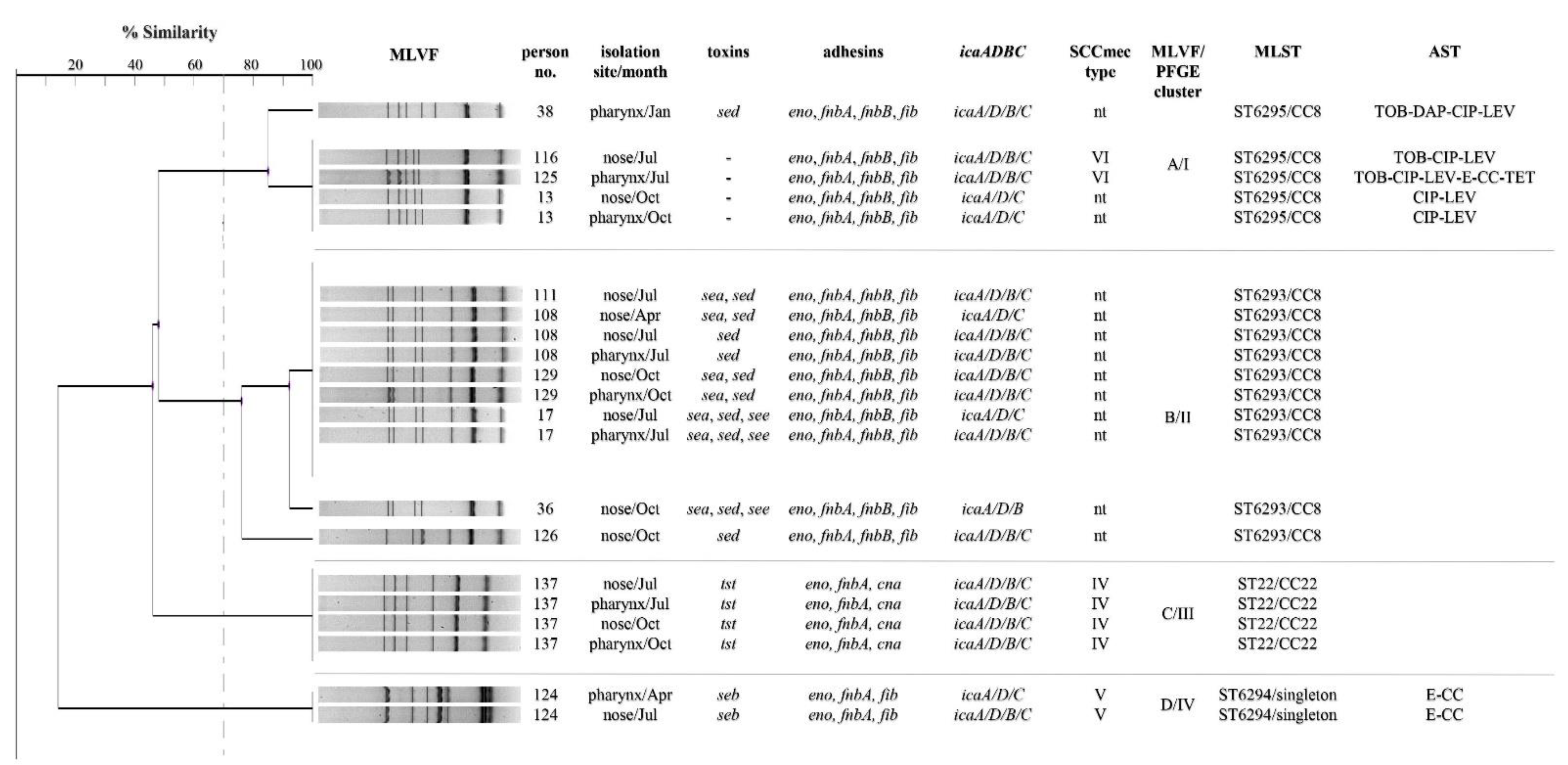
2.4. Genotyping
3. Discussion
4. Materials and Methods
4.1. Collection of Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. Bacterial DNA Isolation
4.4. Detection of Genes Encoding Toxins and Adhesion Factors
4.5. Genotyping of Methicillin-Resistant Staphylococcus aureus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kong, C.; Neoh, H.; Nathan, S. Targeting Staphylococcus aureus toxins: A potential form of anti-virulence therapy. Toxins 2016, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diep, B.A.; Otto, M. The role of virulence determinants in community-associated MRSA pathogenesis. Trends Microbiol. 2008, 16, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Henderson, A.; Nimmo, G.R. Control of healthcare- and community-associated MRSA: Recent progress and persisting challenges. Br. Med. Bull. 2018, 125, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Ruhe, J.J.; Smith, N.; Bradsher, R.W.; Menon, A. Community-onset methicillin-resistant Staphylococcus aureus skin and soft-tissue infections: Impact of antimicrobial therapy on outcome. Clin. Infect. Dis. 2007, 44, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantosti, A.; Sanchini, A.; Monaco, M. Mechanisms of antibiotic resistance in Staphylococcus aureus. Future Microbiol. 2007, 2, 323–334. [Google Scholar] [CrossRef]
- Cho, S.-H.; Strickland, I.; Boguniewicz, M.; Leung, D.Y.M. Fibronectin and fibrinogen contribute to the enhanced binding of Staphylococcus aureus to atopic skin. J. Allergy Clin. Immunol. 2001, 108, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A. Role of Staphylococcus aureus virulence factors in inducing inflammation and vascular permeability in a mouse model of bacterial endophthalmitis. PLoS ONE 2015, 10, e0128423. [Google Scholar] [CrossRef] [Green Version]
- Spaulding, A.R.; Salgado-Pabón, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.M.; Schlievert, P.M. Staphylococcal and streptococcal superantigen exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 1–23. [Google Scholar] [CrossRef]
- Otto, M. MRSA virulence and spread. Cell Microbiol. 2012, 14, 1513–1521. [Google Scholar] [CrossRef] [Green Version]
- Stenehjem, E.; Rimland, D. MRSA nasal colonization burden and risk of MRSA infection. Am. J. Infect. Control. 2013, 41, 405–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastagia, M.; Kleinman, L.C.; Lacerda de la Cruz, E.G.; Jenkins, S.G. Predicting risk for death from MRSA bacteremia. Emerg. Infect. Dis. 2012, 18, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Zhen, X.; Lundborg, C.S.; Zhang, M.; Sun, X.; Li, Y.; Hu, X.; Gu, S.; Gu, Y.; Wei, J.; Dong, H. Clinical and economic impact of methicillin-resistant Staphylococcus aureus: A multicentre study in China. Sci. Rep. 2020, 10, 3900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denis, O.; Jans, B.; Deplano, A.; Nonhoff, C.; Struelens, M.J. Epidemiology of methicillin-resistant Staphylococcus aureus (MRSA) among residents of nursing homes in Belgium. J. Antimicrob. Chemother. 2009, 64, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Gruber, I.; Heudorf, U.; Werner, G.; Pfeifer, Y.; Imirzalioglu, C.; Ackermann, H.; Brandt, C.; Besier, S.; Wichelhaus, T.A. multidrug-resistant bacteria in geriatric clinics, nursing homes, and ambulant care–prevalence and risk factors. Int. J. Med. Microbiol. 2013, 303, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Budimir, A.; Payerl Pal, M.; Bošnjak, Z.; Mareković, I.; Vuković, D.; Roksandić Križan, I.; Milas, J.; Plečko, V.; Kalenić, S. Prevalence and molecular characteristics of methicillin-resistant Staphylococcus aureus strains isolated in a multicenter study of nursing home residents in Croatia. Am. J. Infect. Control. 2014, 42, 1197–1202. [Google Scholar] [CrossRef]
- Ho, P.-L.; Lai, E.L.; Chow, K.-H.; Chow, L.S.M.; Yuen, K.-Y.; Yung, R.W.H. Molecular epidemiology of methicillin-resistant Staphylococcus aureus in residential care homes for the elderly in Hong Kong. Diagn. Microbiol. Infect. Dis. 2008, 61, 135–142. [Google Scholar] [CrossRef]
- Pfingsten-Würzburg, S.; Pieper, D.H.; Bautsch, W.; Probst-Kepper, M. Prevalence and molecular epidemiology of meticillin-resistant Staphylococcus aureus in nursing home residents in northern Germany. J. Hosp. Infect. 2011, 78, 108–112. [Google Scholar] [CrossRef]
- Daeschlein, G.; Assadian, O.; Rangous, I.; Kramer, A. Risk factors for Staphylococcus aureus nasal carriage in residents of three nursing homes in Germany. J. Hosp. Infect. 2006, 63, 216–220. [Google Scholar] [CrossRef]
- Hoogendoorn, M.; Smalbrugge, M.; Stobberingh, E.E.; van Rossum, S.V.; Vlaminckx, B.J.; Thijsen, S.F. Prevalence of antibiotic resistance of the commensal flora in Dutch nursing homes. J. Am. Med. Dir. Assoc. 2013, 14, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Moschou, A. Prevalence and molecular epidemiology of Staphylococcus aureus nasal colonization in four nursing home residents in Crete, Greece. J. Infect. Chemother. 2020, 6, 199–204. [Google Scholar] [CrossRef]
- Rondeau, C.; Chevet, G.; Blanc, D.S.; Gbaguidi-Haore, H.; Decalonne, M.; Dos Santos, S.; Quentin, R.; van der Mee-Marquet, N. Current molecular epidemiology of methicillin-resistant Staphylococcus aureus in elderly French people: Troublesome clones on the horizon. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dalhoff, A. Global fluoroquinolone resistance epidemiology and implications for clinical use. Interdiscip. Perspect Infect. Dis. 2012, 1–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDanel, J.S.; Murphy, C.R.; Diekema, D.J.; Quan, V.; Kim, D.S.; Peterson, E.M.; Evans, K.D.; Tan, G.L.; Hayden, M.K.; Huang, S.S. Chlorhexidine and Mupirocin Susceptibilities of Methicillin-Resistant Staphylococcus aureus from colonized nursing home residents. Antimicrob. Agents Chemother. 2013, 57, 552–558. [Google Scholar] [CrossRef] [Green Version]
- Scerri, J.; Monecke, S.; Borg, M.A. Prevalence and characteristics of community carriage of methicillin-resistant Staphylococcus aureus in Malta. J. Epidemiol. Glob. Health 2013, 3, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef] [Green Version]
- Bergevin, M.; Marion, A.; Farber, D.; Golding, G.R.; Lévesque, S. Severe MRSA enterocolitis caused by a strain harboring enterotoxins D., G., and I. Emerg. Infect. Dis. 2017, 23, 865–867. [Google Scholar] [CrossRef] [Green Version]
- Fooladvand, S.; Sarmadian, H.; Habibi, D.; van Belkum, A.; Ghaznavi-Rad, E. High Prevalence of methicillin resistant and enterotoxin gene-positive Staphylococcus aureus among nasally colonized food handlers in central Iran. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 87–92. [Google Scholar] [CrossRef]
- DeVries, A.S.; Lesher, L.; Schlievert, P.M.; Rogers, T.; Villaume, L.G.; Danila, R.; Lynfield, R. Staphylococcal toxic shock syndrome 2000–2006: Epidemiology, clinical features, and molecular characteristics. PLoS ONE 2011, 6, e22997. [Google Scholar] [CrossRef]
- Mehrotra, M.; Wang, G.; Johnson, W.M. Multiplex PCR for Detection of genes for Staphylococcus aureus enterotoxins, exfoliative toxins, toxic shock syndrome toxin 1, and methicillin resistance. J. Clin. Microbiol. 2000, 38, 1032–1035. [Google Scholar] [CrossRef] [Green Version]
- Asadollahi, P.; Delpisheh, A.; Maleki, M.H.; Azizi Jalilian, F.; Alikhani, M.Y.; Asadollahi, K.; Soroush, S.; Hematian, A.; Emaneini, M.; Taherikalani, M. Enterotoxin and exfoliative toxin genes among methicillin-resistant Staphylococcus aureus isolates recovered from Ilam, Iran. Avicenna J. Clin. Microbiol. Infect. 2014, 1, e20208. [Google Scholar] [CrossRef]
- Chikkala, R.; George, N.O.; Ratnakar, K.S.; Iyer, R.N.; Sritharan, V. Heterogeneity in femA in the Indian isolates of Staphylococcus aureus limits its usefulness as a species specific marker. Adv. Infect. Dis. 2012, 2, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Di Domenico, E.; Farulla, I.; Prignano, G.; Gallo, M.; Vespaziani, M.; Cavallo, I.; Sperduti, I.; Pontone, M.; Bordignon, V.; Cilli, L.; et al. Biofilm is a major virulence determinant in bacterial colonization of chronic skin ulcers independently from the multidrug resistant phenotype. Int. J. Mol. Sci. 2017, 18, 1077. [Google Scholar] [CrossRef]
- Schaffer, A.C.; Solinga, R.M.; Cocchiaro, J.; Portoles, M.; Kiser, K.B.; Risley, A.; Randall, S.M.; Valtulina, V.; Speziale, P.; Walsh, E.; et al. Immunization with Staphylococcus aureus clumping factor B, a major determinant in nasal carriage, reduces nasal colonization in a murine model. Infect. Immun. 2006, 74, 2145–2153. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, E.; Pozzi, C.; Houston, P.; Humphreys, H.; Robinson, D.A.; Loughman, A.; Foster, T.J.; O’Gara, J.P. A Novel Staphylococcus aureus biofilm phenotype mediated by the fibronectin-binding proteins, FnBPA and FnBPB. J. Bacteriol. 2008, 190, 3835–3850. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, A.; Diep, B.A.; Mai, T.T.; Vo, N.H.; Warrener, P.; Suzich, J.; Stover, C.K.; Sellman, B.R. Differential expression and roles of Staphylococcus aureus virulence determinants during colonization and disease. mBio 2015, 6, e02272-14. [Google Scholar] [CrossRef] [Green Version]
- Fluckiger, U.; Ulrich, M.; Steinhuber, A.; Döring, G.; Mack, D.; Landmann, R.; Goerke, C.; Wolz, C. Biofilm formation, icaADBC transcription, and polysaccharide intercellular adhesin synthesis by staphylococci in a device-related infection model. Infect. Immun. 2005, 73, 1811–1819. [Google Scholar] [CrossRef] [Green Version]
- Otter, J.A.; French, G.L. Molecular epidemiology of community-associated meticillin-resistant Staphylococcus aureus in Europe. Lancet Infect. Dis. 2010, 10, 227–239. [Google Scholar] [CrossRef]
- Kasela, M.; Grzegorczyk, A.; Korona-Głowniak, I.; Ossowski, M.; Nowakowicz-Dębek, B.; Malm, A. Transmission and long-term colonization patterns of Staphylococcus aureus in a nursing home. Int. J. Environ. Res. Public Health 2020, 17, 8073. [Google Scholar] [CrossRef]
- McClure, J.-A.; Conly, J.M.; Lau, V.; Elsayed, S.; Louie, T.; Hutchins, W.; Zhang, K. Novel multiplex PCR assay for detection of the staphylococcal virulence marker Panton-Valentine leukocidin genes and simultaneous discrimination of methicillin-susceptible from -resistant staphylococci. J. Clin. Microbiol. 2006, 44, 1141–1144. [Google Scholar] [CrossRef] [Green Version]
- Atshan, S.S.; Nor Shamsudin, M.; Sekawi, Z.; Lung, L.T.T.; Hamat, R.A.; Karunanidhi, A.; Mateg Ali, A.; Ghaznavi-Rad, E.; Ghasemzadeh-Moghaddam, H.; Chong Seng, J.S.; et al. Prevalence of adhesion and regulation of biofilm-related genes in different clones of Staphylococcus aureus. J. Biomed. Biotechnol. 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Sabat, A.; Krzyszton-Russjan, J.; Strzalka, W.; Filipek, R.; Kosowska, K.; Hryniewicz, W.; Travis, J.; Potempa, J. New method for typing Staphylococcus aureus strains: Multiple-locus variable-number tandem repeat analysis of polymorphism and genetic relationships of clinical isolates. J. Clin. Microbiol. 2003, 41, 1801–1804. [Google Scholar] [CrossRef] [Green Version]
- Grzegorczyk, A.; Malm, A. Genotyping of Staphylococcus aureus strains isolated from healthy persistent carriers. Folia Microbiol. 2014, 59, 349–353. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [Green Version]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Category | Number (%) of Resistant MRSA Strains | ||
|---|---|---|---|
| Residents n = 6 | Personnel n = 15 | Total n = 21 | |
| Resistant only to β-lactams | 3 (50) | 11 (73.4) | 14 (66.7) |
| Resistant to 1 other class | 2 (33.3) | 2 (13.3) | 4 (19) |
| Resistant to 2 other classes | 0 | 1 (6.7) | 1 (4.8) |
| Resistant to ≥3 other classes | 1 (16.7) | 1 (6.7) | 2 (9.5) |
| Gene | Primers | Sequence (5′–3′) | Size (bp) | Reference |
|---|---|---|---|---|
| sea | sea-F | GGTTATCAATGTGCGGGTGG | 102 | [31] |
| sea-R | CGGCACTTTTTTCTCTTCGG | |||
| seb | seb-F | GTATGGTGGTGTAACTGAGC | 164 | |
| seb-R | CCAAATAGTGACGAGTTAGG | |||
| sec | sec-F | AGATGAAGTAGTTGATGTGTATGG | 451 | |
| sec-R | CACACTTTTAGAATCAACCG | |||
| sed | sed-F | CCAATAATAGGAGAAAATAAAAG | 278 | |
| sed-R | ATTGGTATTTTTTTTCGTTC | |||
| see | see-F | AGGTTTTTTCACAGGTCATCC | 209 | |
| see-R | CTTTTTTTTCTTCGGTCAATC | |||
| eta | eta-F | GCAGGTGTTGATTTAGCATT | 93 | |
| eta-R | AGATGTCCCTATTTTTGCTG | |||
| etb | etb-F | ACAAGCAAAAGAATACAGCG | 226 | |
| etb-R | GTTTTTGGCTGCTTCTCTTG | |||
| tst | tst-F | ACCCCTGTTCCCTTATCATC | 326 | |
| tst-R | TTTTCAGTATTTGTAACGCC | |||
| lukS-F-PV | luk-PV-F | ATCATTAGGTAAAATGTCTGGACATGATCCA | 433 | [41] |
| luk-PV-R | GCATCAAGTGTATTGGATAGCAAAAGC | |||
| eno | eno-F | ACGTGCAGCAGCTGACT | 301 | [42] |
| eno-R | CAACAGCATCTTCAGTACCTTC | |||
| fnbB | fnbB-F | GTAACAGCTAATGGTCGAATTGATACT | 523 | |
| fnbB-R | CAAGTTCGATAGGAGTACTATGTTC | |||
| cna | cna-F | AAAGCGTTGCCTAGTGGAGA | 192 | |
| cna-R | AGTGCCTTCCCAAACCTTTT | |||
| fnbA | fnbA-F | CATAAATTGGGAGCAGCATCA | 128 | |
| fnbA-R | ATCAGCAGCTGAATTCCCATT | |||
| ebpS | ebpS-F | CATCCAGAACCAATCGAAGAC | 180 | |
| ebpS-R | AGTTACATCATCATGTTTATCTTTTG | |||
| fib | fib-F | CTACAACTACAATTGCGTCAACAG | 405 | |
| fib-R | GCTCTTGTAAGACCATTTTCTTCAC | |||
| bbp | bbp-F | AACTACATCTAGTACTCAACAACAG | 574 | |
| bbp-R | ATGTGCTTGAATAACACCATCATCT | |||
| icaA | icaA-F | ACACTTGCTGGCGCAGTCAA | 188 | |
| icaA-R | TCTGGAACCAACATCCAACA | |||
| icaD | icaD-F | ATGGTCAAGCCCAGACAGAG | 198 | |
| icaD-R | AGTATTTTCAATGTTTAAAGCAA | |||
| icaB | icaB-F | AGAATCGTGAAGTATAGAAAATT | 900 | |
| icaB-R | TCTAATCTTTTTCATGGAATCCGT | |||
| icaC | icaC-F | ATGGGACGGATTCCATGAAAAAGA | 1100 | |
| icaC-R | TAATAAGCATTAATGTTCAATT | |||
| femA | femA-F | AAAAAAGCACATAACAAGCG | 132 | [31] |
| femA-R | GATAAAGAAGAAACCAGCAG | |||
| mecA | mecA-F | ACTGCTATCCACCCTCAAAC | 163 | |
| mecA-R | CTGGTGAAGTTGTAATCTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasela, M.; Grzegorczyk, A.; Nowakowicz-Dębek, B.; Malm, A. The Prevalence of Virulence Determinants and Antibiotic Resistance Patterns in Methicillin—Resistant Staphylococcus aureus in a Nursing Home in Poland. Pathogens 2021, 10, 427. https://doi.org/10.3390/pathogens10040427
Kasela M, Grzegorczyk A, Nowakowicz-Dębek B, Malm A. The Prevalence of Virulence Determinants and Antibiotic Resistance Patterns in Methicillin—Resistant Staphylococcus aureus in a Nursing Home in Poland. Pathogens. 2021; 10(4):427. https://doi.org/10.3390/pathogens10040427
Chicago/Turabian StyleKasela, Martyna, Agnieszka Grzegorczyk, Bożena Nowakowicz-Dębek, and Anna Malm. 2021. "The Prevalence of Virulence Determinants and Antibiotic Resistance Patterns in Methicillin—Resistant Staphylococcus aureus in a Nursing Home in Poland" Pathogens 10, no. 4: 427. https://doi.org/10.3390/pathogens10040427