
Staphylococcus saccharolyticus Associated with Prosthetic Joint Infections: Clinical Features and Genomic Characteristics


Abstract
:1. Introduction
2. Results
2.1. Clinical Data
2.2. Antibiotic Susceptibility Testing
2.3. Genome Sequencing of Clinical S. saccharolyticus Isolates
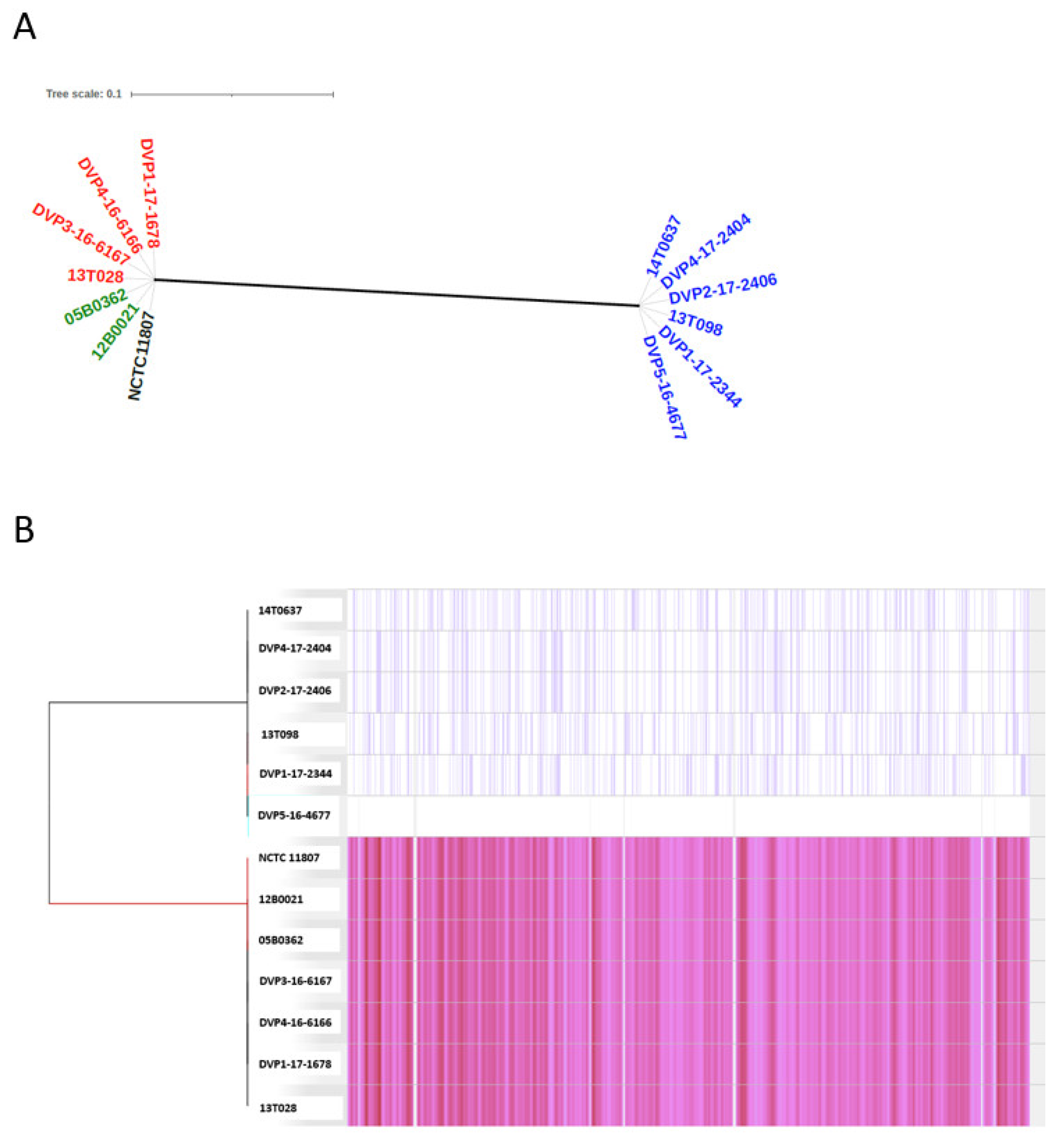
2.3.1. Population Structure Based on Core-Genome Comparison
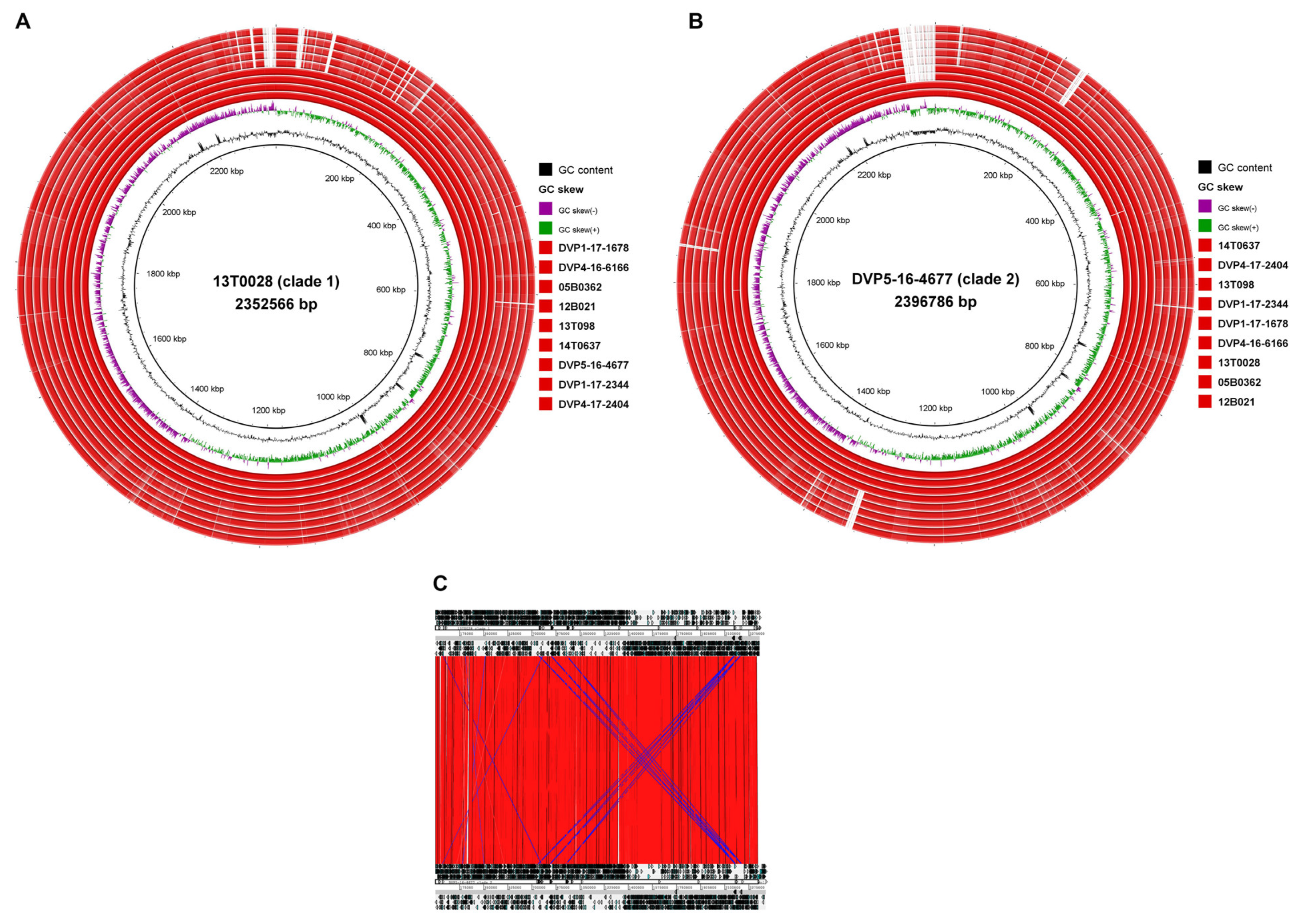
2.3.2. Genome-Wide Synteny with a Small Accessory Genome
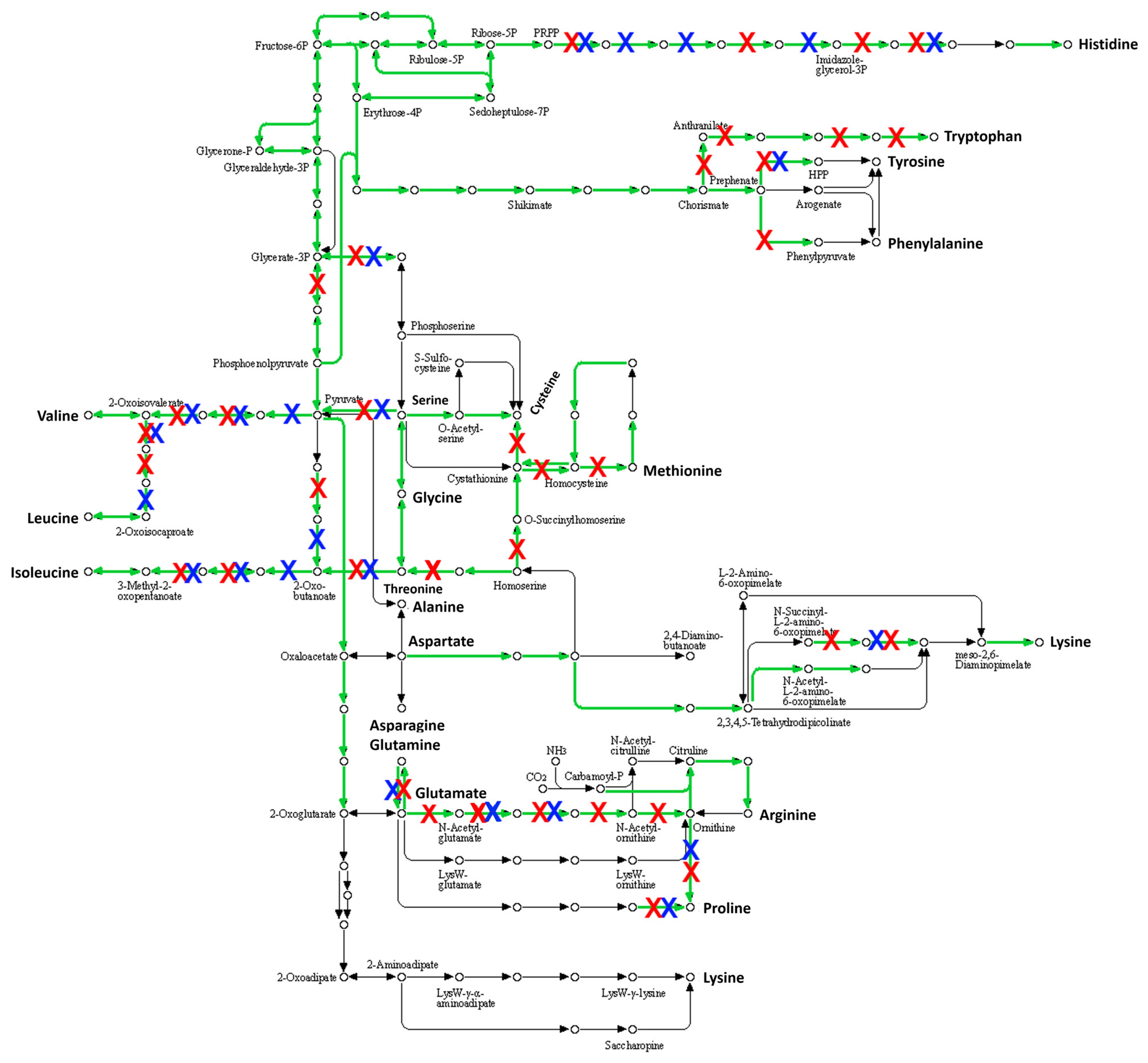
2.3.3. Many Pseudogenes in the Genome of S. saccharolyticus
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Bacterial Isolates
4.3. Antibiotic Susceptibility Testing
4.4. Clinical Data
4.5. Genome Sequencing of S. saccharolyticus Isolates
4.6. Bioinformatics and Phylogenetic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evans, C.A.; Mattern, K.L.; Hallam, S.L. Isolation and identification of Peptococcus saccharolyticus from human skin. J. Clin. Microbiol. 1978, 7, 261–264. [Google Scholar] [PubMed]
- Ahle, C.M.; Stødkilde, K.; Afshar, M.; Poehlein, A.; Ogilvie, L.A.; Söderquist, B.; Hüpeden, J.; Brüggemann, H. Staphylococcus saccharolyticus: An overlooked human skin colonizer. Microorganisms 2020, 8, 1105. [Google Scholar] [CrossRef] [PubMed]
- Brüggemann, H.; Poehlein, A.; Brzuszkiewicz, E.; Scavenius, C.; Enghild, J.J.; Al-Zeer, M.A.; Brinkmann, V.; Jensen, A.; Söderquist, B. Staphylococcus saccharolyticus isolated from blood cultures and prosthetic joint infections exhibits excessive genome decay. Front. Microbiol. 2019, 10, 478. [Google Scholar] [CrossRef]
- Godreuil, S.; Jean-Pierre, H.; Morel, J.; Darbas, H.; Jumas-Bilak, E.; Bañuls, A.L.; Marchandin, H. Unusual case of spondylodiscitis due to Staphylococcus saccharolyticus. Jt. Bone Spine 2005, 72, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Mikhael, M.M.; Bach, H.G.; Huddleston, P.M.; Maus, T.P.; Berbari, E.F. Multilevel diskitis and vertebral osteomyelitis after diskography. Orthopedics 2009, 32, 60. [Google Scholar] [CrossRef] [PubMed]
- Trojani, M.C.; Lamy, B.; Ruimy, R.; Amoretti, N.; Risso, K.; Roux, C. An unusual Staphylococcus saccharolyticus spondylodiscitis post kyphoplasty: A case report. BMC Infect. Dis. 2020, 20, 539. [Google Scholar] [CrossRef] [PubMed]
- Westblom, T.U.; Gorse, G.J.; Milligan, T.W.; Schindzielorz, A.H. Anaerobic endocarditis caused by Staphylococcus saccharolyticus. J. Clin. Microbiol. 1990, 28, 2818–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, S.; Haglund, L.; Ashfaq, A.; Leist, P.; Roat, T. Prosthetic valve endocarditis due to Staphylococcus saccharolyticus. Clin. Infect. Dis. 1996, 22, 722–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, N.; Bhally, H. Bilateral neck pyomyositis caused by Staphylococcus capitis and Staphylococcus saccharolyticus in a diabetic adult. Case Rep. Infect. Dis. 2017, 2017, 3713212. [Google Scholar] [PubMed] [Green Version]
- Wu, X.; Yu, C.; Wang, X. A case of Staphylococcus saccharolyticus pneumonia. Int. J. Infect. Dis. 2009, 13, e43–e46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Liu, Y.; Xu, Y.; Xu, Z. Staphylococcus saccharolyticus infection: Case series with a PRISMA-compliant systemic review. Medicine 2020, 26, e20686. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.J.; Sun, B.; Guo, J.; Liu, C.J.; Sun, B.; Guo, J. A case of bone marrow infection by Staphylococcus saccharolyticus. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1161–1163. [Google Scholar] [PubMed]
- Steinbrueckner, B.; Singh, S.; Freney, J.; Kuhnert, P.; Pelz, K.; Aufenanger, J. Facing a mysterious hospital outbreak of bacteraemia due to Staphylococcus saccharolyticus. J. Hosp. Infect. 2001, 49, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, A.G.; Yian, E.; Steens, W. Injection-induced low-grade infection of the shoulder joint: Preliminary results. Arch. Orthop. Trauma Surg. 2012, 132, 1387–1392. [Google Scholar] [CrossRef]
- Shah, N.B.; Tande, A.J.; Patel, R.; Berbari, E.F. Anaerobic prosthetic joint infection. Anaerobe 2015, 36, 1–8. [Google Scholar] [CrossRef]
- Osmon, D.R.; Berbari, E.F.; Berendt, A.R.; Lew, D.; Zimmerli, W.; Steckelberg, J.M.; Rao, N.; Hanssen, A.; Wilson, W.R.; Infectious Diseases Society of America. Diagnosis and management of prosthetic joint infection: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, e1–e25. [Google Scholar] [CrossRef] [Green Version]
- Afshar, M.; Poehlein, A.; Söderquist, B.; Brüggemann, H. Complete genome sequences of Two Staphylococcus saccharolyticus strains isolated from prosthetic joint infections. Microbiol. Resour. Announc. 2021, 10, e00157-21. [Google Scholar] [CrossRef]
- Flurin, L.; Greenwood-Quaintance, K.E.; Patel, R. Microbiology of polymicrobial prosthetic joint infection. Diagn. Microbiol. Infect. Dis. 2019, 94, 255–259. [Google Scholar] [CrossRef]
- Falk-Brynhildsen, K.; Friberg, Ö.; Söderquist, B.; Nilsson, U. Bacterial recolonisation of the skin and wound contamination during cardiac surgery: A randomized controlled trial of the use of plastic adhesive drape compared with bare skin. J. Hosp. Infect. 2013, 84, 151–158. [Google Scholar] [CrossRef]
- Fernandez, R.C.; Weiss, A.A. Cloning and sequencing of a Bordetella pertussis serum resistance locus. Infect. Immun. 1994, 62, 4727–4738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Carver, T.J.; Rutherford, K.M.; Berriman, M.; Rajandream, M.A.; Barrell, B.G.; Parkhill, J. ACT: The Artemis Comparison Tool. Bioinformatics 2005, 21, 3422–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pat | Patient ID Sex, Age | Isolate ID | Affected Joint | Characteristics (Diagnosis, Surgical Procedure) | Time to Diagnosis (from Primary Surgery) | Duration of Symptoms | Type of Infection (Number of Tissue Biopsies with Growth) | Antimicrobial Treatment, Duration | Surgical Procedure and Follow-Up |
|---|---|---|---|---|---|---|---|---|---|
| 1 | ♂, 76 | DVP2-17- 2406 | Hip | Osteoarthritis. Primary arthroplasty surgery 2002 (Exeter). Loosening of the cup in 2015. | 13 years | Pain 2 years, affected functional status | Monomicrobial Staphylococcus saccharolyticus, 2/5 positive cultures | No antibiotics administered | One-stage exchange procedure of cup and femoral head due to loosening. Two years later good functional status, no walking aid. |
| 2 | ♂, 65 | DVP5-16-4677 | Hip | Osteoarthritis. Primary arthroplasty surgery May 2014 (Stryker). Loosening of the cup in July 2015. | 14 Months | Pain 1 year, affected functional status | Monomicrobial Staphylococcus saccharolyticus, 3/5 positive cultures | Amoxicillin, 4 months | One-stage exchange procedure of cup and femoral head due to loosening. One year later good functional status, no walking aid. |
| 3 | ♂, 76 | 14T637 | Hip | Osteoarthritis. Primary arthroplasty surgery 1998 (Exeter). Loosening of the cup in 2014. | 16 years | Pain ca 6 months, affected functional status | Polymicrobial Cutibacterium acnes 4/5, Staphylococcus saccharolyticus 1/5 | Amoxicillin, 3 months | One-stage exchange procedure of cup due to loosening. One year later acceptable functional status, walking aids sporadically. |
| 4 | ♂, 69 | 13T098 | Hip | Osteoarthritis. Primary arthroplasty surgery 2005 (Exeter). Loosening of the cup in 2012. | 7 years | Pain ca 1 year | Monomicrobial Staphylococcus saccharolyticus, 3/5 positive cultures | No antibiotics administered | One-stage exchange procedure of cup and stem due to loosening. One year later excellent functional status, no walking aid. |
| 5 | ♂, 75 | DVP1-17-2344 | Hip | Osteoarthritis. Primary arthroplasty surgery 2001 (Lubinus). Loosening of the cup in 2016. | 15 years | Pain ca 6 moths | Polymicrobial Staphylococcus saccharolyticus 3/5, Cutibacterium acnes 2/5, | Amoxicillin, 3 months | One-stage exchange procedure of all prosthetic devices due to loosening. One year later excellent functional status, no walking aid. |
| 6 | ♂, 72 | 13T028 | Shoulder | Traumatic fracture of proximal humerus. Global FX Shoulder Prosthesis 2012. Early post-operative infection. | 21 days | Pain, swelling, local inflammation 17 days post-opertively | Polymicrobial Staphylococcus saccharolyticus 5/5, Cutibacterium acnes 4/5 | Penicillin V, 4 months | Revision and lavage 1 month post-operatively. One year later poor functional status, ROM 30°. |
| 7 | ♂, 49 | DVP4-16-6166, DVP1-17-1678 | Shoulder | Secondary osteoarthritis due to Staphylococcus aureus osteomyelitis. Tissue biopsies at primary arthroplasty surgery (Global Unite) showed growth of Staphylococcus saccharolyticus 2/6, Cutibacterium acnes 1/6 | 5 months | Pain, swelling, local inflammation post-operatively | Polymicrobial Staphylococcus saccharolyticus 3/5, Cutibacterium acnes 1/5 | Penicillin V, 3 months | Two-stage exchange procedure. Antibiotic treatment; 14 months follow-up shows good functional status. |
| Patient Lab ID | 1 DVP2-17-2406 | 2 DVP5-16-4677 | 3 14T637 | 4 13T098 | 5 DVP1-17-2344 | 6 13T028 | 7 DVP4-16-6166 |
|---|---|---|---|---|---|---|---|
| Benzyl-penicillin | 0.002 | <0.002 | 0.004 | 0.003 | <0.002 | 0.003 | 0.002 |
| Clindamycin | 0.016 | 0.032 | 0.032 | 0.032 | 0.032 | 0.047 | 0.125 |
| Metronidazole | >256 | >256 | >256 | >256 | >256 | >256 | >256 |
| Amoxicillin | <0.016 | <0.016 | <0.016 | <0.016 | <0.016 | <0.016 | <0.016 |
| Imipenem | <0.008 | <0.004 | <0.008 | <0.016 | <0.008 | <0.004 | <0.004 |
| Meropenem | <0.002 | <0.002 | <0.002 | <0.002 | <0.002 | <0.002 | <0.002 |
| Vancomycin | 0.5 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Chloramphenicol | 4 | 1 | 1 | 2 | 2 | 0.125 | 0.16 |
| Strain | Source | Genome Size (kb) | G+C (%) | Contigs | Coverage | N50 (kb) | Clade | CDS | Pseudogenes | Accession Number |
|---|---|---|---|---|---|---|---|---|---|---|
| DVP2-17-2406 | Patient 1, hip | 2373 | 32.00 | 11 | 222 | 1222 | 2 | 1740 | 502 | QHKD00000000 |
| DVP5-16-4677 | Patient 2, hip | 2396 | 32.08 | 2 | 188 | - | 2 | 1726 | 493 | Chromosome: CP068031.1 Plasmid: CP068032.1 |
| 14T637 | Patient 3, hip | 2375 | 32.00 | 13 | 257 | 521 | 2 | 1732 | 487 | JAENGT000000000 |
| 13T098 | Patient 4, hip | 2321 | 32.10 | 10 | 225 | 1224 | 2 | 1704 | 485 | JAENGW000000000 |
| DVP1-17-2344 | Patient 5, hip | 2321 | 32.10 | 10 | 254 | 1223 | 2 | 1711 | 485 | JAENGU000000000 |
| 13T028 | Patient 6, shoulder | 2352 | 32.16 | 2 | 205 | - | 1 | 1750 | 469 | Chromosome: CP068029.1 Plasmid: CP068030.1 |
| DVP4-16-6166 | Patient 7, shoulder | 2349 | 32.00 | 13 | 291 | 749 | 1 | 1778 | 470 | JAENGX000000000 |
| DVP1-17-1678 | Patient 7, shoulder | 2349 | 32.00 | 10 | 200 | 768 | 1 | 1775 | 471 | JAENGV000000000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Söderquist, B.; Afshar, M.; Poehlein, A.; Brüggemann, H. Staphylococcus saccharolyticus Associated with Prosthetic Joint Infections: Clinical Features and Genomic Characteristics. Pathogens 2021, 10, 397. https://doi.org/10.3390/pathogens10040397
Söderquist B, Afshar M, Poehlein A, Brüggemann H. Staphylococcus saccharolyticus Associated with Prosthetic Joint Infections: Clinical Features and Genomic Characteristics. Pathogens. 2021; 10(4):397. https://doi.org/10.3390/pathogens10040397
Chicago/Turabian StyleSöderquist, Bo, Mastaneh Afshar, Anja Poehlein, and Holger Brüggemann. 2021. "Staphylococcus saccharolyticus Associated with Prosthetic Joint Infections: Clinical Features and Genomic Characteristics" Pathogens 10, no. 4: 397. https://doi.org/10.3390/pathogens10040397