
Development of a Recombinant Pichinde Virus-Vectored Vaccine against Turkey Arthritis Reovirus and Its Immunological Response Characterization in Vaccinated Animals


 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Pichinde Virus Plasmids
2.3. Preparation of Vectors and Gene Inserts
2.4. Cloning and Transfection
2.5. Detection of Reovirus Antigenic Proteins
2.6. Vaccination Experiment
2.7. Serum Neutralization Assay
3. Results
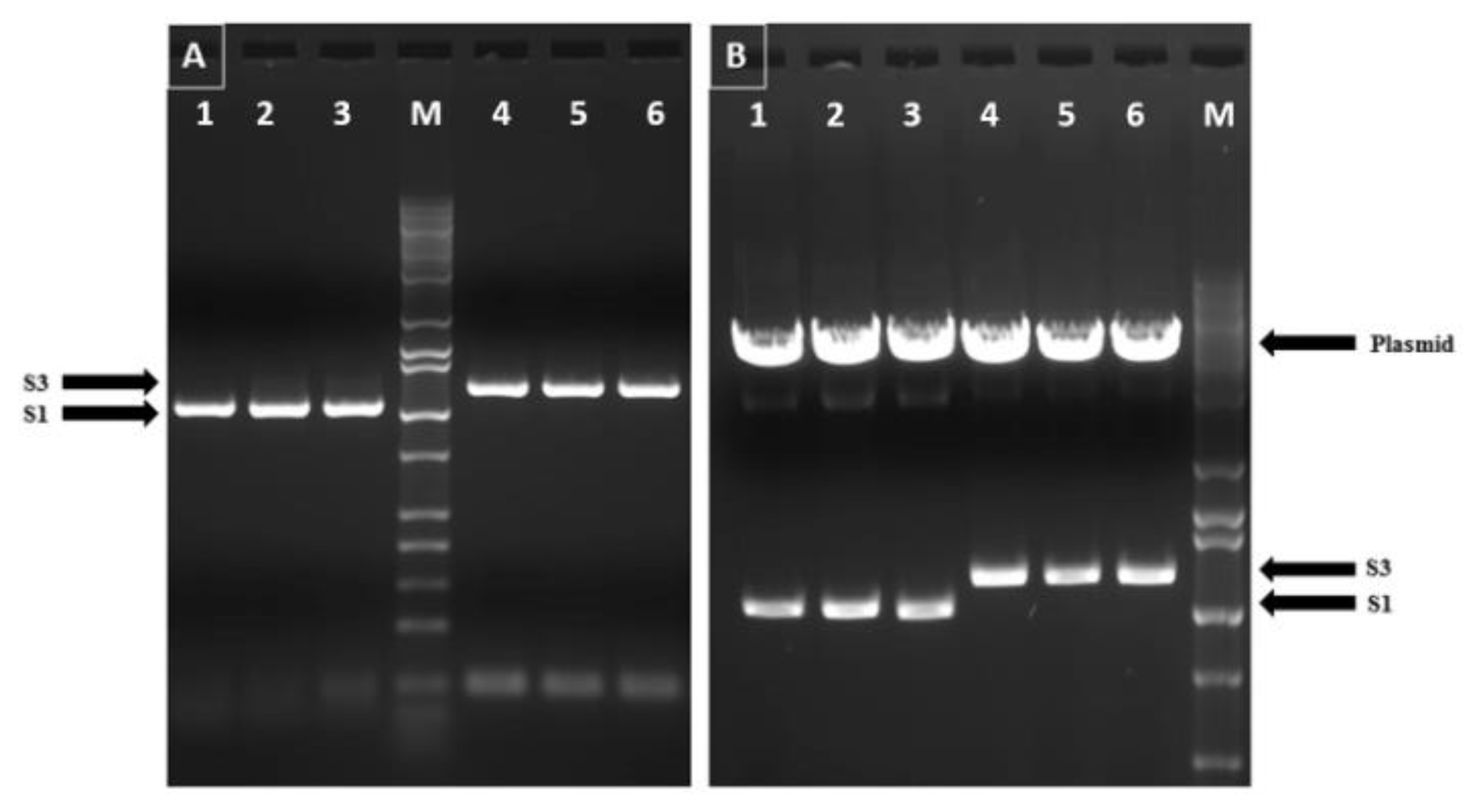
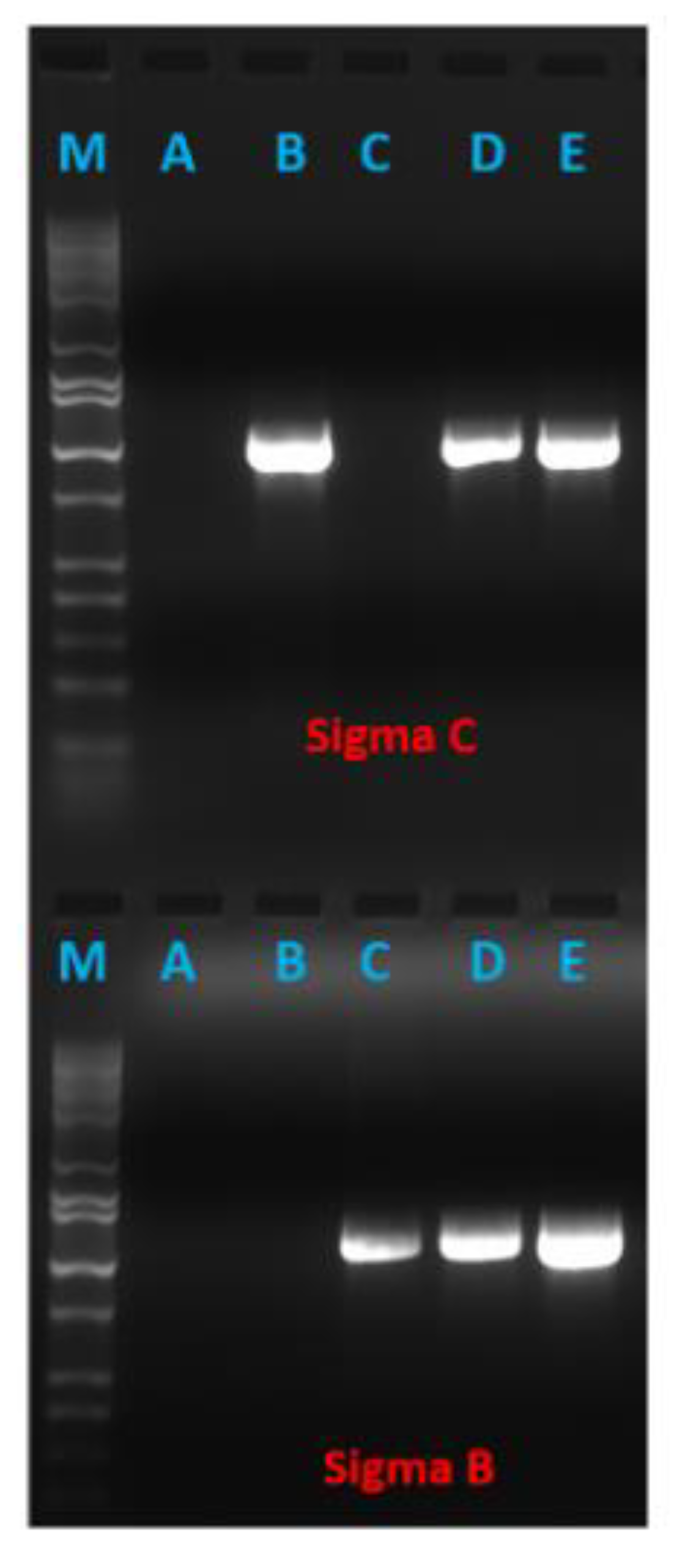
3.1. Cloning of Reovirus Genes into PICV Plasmids
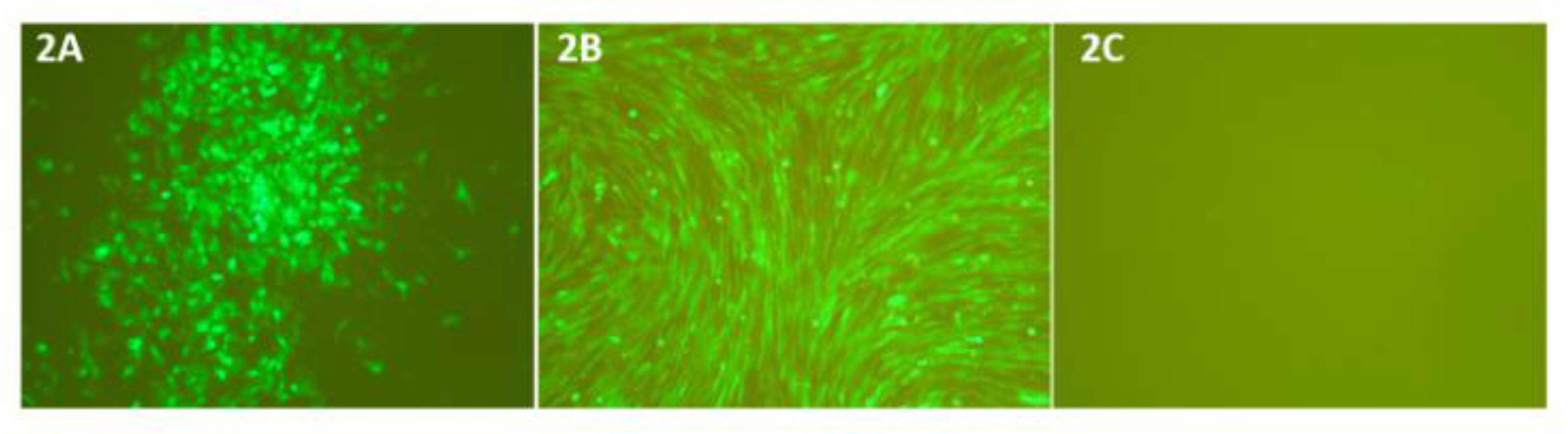
3.2. Plasmid Transfection and Virus Rescue
3.3. Recombinant PICVs Expressing Reovirus Antigens
3.4. Clinical Signs and Necropsy
3.5. Serum Neutralization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mor, S.K.; Sharafeldin, T.A.; Porter, R.E.; Ziegler, A.; Patnayak, D.P.; Goyal, S.M. Isolation and characterization of a turkey arthritis reovirus. Avian Dis. 2013, 57, 97–103. [Google Scholar] [CrossRef]
- Sharafeldin, T.A.; Mor, S.K.; Bekele, A.Z.; Verma, H.; Goyal, S.M.; Porter, R.E. The role of avian reoviruses in turkey tenosynovitis/arthritis. Avian Pathol. 2014, 43, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharafeldin, T.A.; Mor, S.K.; Bekele, A.Z.; Verma, H.; Noll, S.L.; Goyal, S.M.; Porter, R.E. Experimentally induced lameness in turkeys inoculated with a newly emergent turkey reovirus. Vet. Res. 2015, 46, 11. [Google Scholar] [CrossRef] [Green Version]
- Benavente, J.; Martinez-Costas, J. Avian reovirus: Structure and biology. Virus Res. 2007, 123, 105–119. [Google Scholar] [CrossRef]
- Varela, R.; Benavente, J. Protein coding assignment of avian reovirus strain S1133. J. Virol. 1994, 68, 6775–6777. [Google Scholar] [CrossRef] [Green Version]
- Grande, A.; Costas, C.; Benavente, J. Subunit composition and conformational stability of the oligomeric form of the avian reovirus cell-attachment protein σC. J. Gen. Virol. 2002, 83, 131–139. [Google Scholar] [CrossRef]
- Huang, W.R.; Wang, Y.C.; Chi, P.I.; Wang, L.; Wang, C.Y.; Lin, C.H.; Liu, H.J. Cell entry of avian reovirus follows a caveolin-1-mediated and dynamin-2-dependent endocytic pathway that requires activation of p38 mitogen-activated protein kinase (MAPK) and Src signaling pathways as well as microtubules and small GTPase Rab5 protein. J. Biol. Chem. 2011, 286, 30780–30794. [Google Scholar] [CrossRef] [Green Version]
- Shih, W.L.; Hsu, H.W.; Liao, M.H.; Lee, L.H.; Liu, H.J. Avian reovirus σC protein induces apoptosis in cultured cells. Virology 2004, 321, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.G.; Fox, G.C.; Parrado, X.L.H.; Saiz, A.L.L.; Costas, C.; Costas, J.M.; Benavente, J.; Raaij, M.J.V. Structure of the carboxy-terminal receptor-binding domain of avian reovirus fibre sigmaC. J. Mol. Biol. 2005, 354, 137–149. [Google Scholar] [CrossRef]
- Martinez-Costas, J.; Grande, A.; Varela, R.; Garcia-Martinez, C.; Benavente, J. Protein architecture of avian reovirus S1133 and identification of the cell attachment protein. J. Virol. 1997, 71, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, R.; Meanger, J.; Enriquez, C.E.; Wilcox, G.E. Avian reovirus proteins associated with neutralization of virus infectivity. Virology 1993, 194, 688–696. [Google Scholar] [CrossRef]
- Goldenberg, D.; Lublin, A.; Rosenbluth, E.; Heller, E.D. Optimized polypeptide for a subunit vaccine against avian reovirus. Vaccine 2016, 34, 3178–3183. [Google Scholar] [CrossRef]
- Wu, H.; Williams, Y.; Gunn, K.S.; Singh, N.K.; Locy, R.D.; Giambrone, J.J. Yeast-derived sigma C protein-induced immunity against avian reovirus. Avian Dis. 2005, 49, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lee, L.H.; Shih, W.L.; Hu, Y.C.; Liu, H.J. Baculovirus surface display of σC and σB proteins of avian reovirus and immunogenicity of the displayed proteins in a mouse model. Vaccine 2008, 26, 6361–6367. [Google Scholar] [CrossRef]
- Bi, Z.; Zhu, Y.; Chen, Z.; Li, C.; Wang, Y.; Wang, G.; Liu, G. Induction of a robust immunity response against novel duck reovirus in ducklings using a subunit vaccine of sigma C protein. Sci. Rep. 2016, 6, 39092. [Google Scholar] [CrossRef] [Green Version]
- Dhanwani, R.; Zhou, Y.; Huang, Q.; Verma, V.; Dileepan, M.; Ly, H.; Lianga, Y. A novel live Pichinde virus-based vaccine vector induces enhanced humoral and cellular immunity upon a booster dose. J. Virol. 2016, 90, 2551–2560. [Google Scholar] [CrossRef] [Green Version]
- Trapido, H.; Sanmartin, C. Pichinde virus, a new virus of the Tacaribe group from Colombia. Am. J. Trop. Med. Hyg. 1971, 20, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Emonet, S.F.; Garidou, L.; McGavern, D.B.; de la Torre, J.C. Generation of recombinant lymphocytic choriomeningitis viruses with trisegmented genomes stably expressing two additional genes of interest. Proc. Natl. Acad. Sci. USA 2009, 106, 3473–3478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flatz, L.; Hegazy, A.N.; Bergthaler, A.; Verschoor, A.; Claus, C.; Fernandez, M.; Gattinoni, L.; Johnson, S.; Kreppel, F.; Kochanek, S.; et al. Development of replication-defective lymphocytic choriomeningitis virus vectors for the induction of potent CD8+ T cell immunity. Nat. Med. 2010, 16, 339–345. [Google Scholar] [CrossRef]
- Popkin, D.L.; Teijaro, J.R.; Lee, A.M.; Lewicki, H.; Emonet, S.; de la Torre, J.C.; Oldstone, M. Expanded potential for recombinant trisegmented lymphocytic choriomeningitis viruses: Protein production, antibody production, and in vivo assessment of biological function of genes of interest. J. Virol. 2011, 85, 7928–7932. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Riano, E.; Cheng, B.Y.; Carlos de la Torre, J.; Martinez-Sobrido, L. Arenavirus reverse genetics for vaccine development. J. Gen. Virol. 2013, 94, 1175–1188. [Google Scholar] [CrossRef]
- Jones, R.C.; Georgiou, K. Reovims-induced tenosynovitis in chickens: The influence of age at infection. Avian Pathol. 1984, 13, 441–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roessler, D.E.; Rosenberger, J.K. In vitro and in vivo characterisation of avian reovirus. III. Host factors affecting virulence and persistence. Avian Dis. 1989, 33, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.C. Avian reovirus infections. Rev. Sci. Tech. Off. Int. Epizoot. 2000, 19, 614–619. [Google Scholar] [CrossRef]
- Giambrone, J.J.; Hathcock, T.L. Efficacy of coarse-spray administration of a reovirus vaccine in young chicks. Avian Dis. 1991, 35, 204–209. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme 5. | Name | Sequence (5’ to 3’) | Number of Bases | Position of Primer on Viral Template (ORFs) | Expected PCR Product Size (Including Primer Sequences) | RE Site | Tag |
|---|---|---|---|---|---|---|---|
| 1 | TARV-S1 F | CGATgctagcGCCACCATGGCCGCTCTAACTCCGTC | 36 | 1–20 | 1031 bp | NheI | - |
| 2 | TARV-S1 R | ATCGctcgagTTACTTGTCGTCATCGTCTTTGTAGTCGGTGTCGATGCCCGTACGCA | 57 | 978–959 | XhoI | FLAG | |
| 3 | TARV-S3 F | CGATgctagcGCCACCATGGAGGTACGTGTGCCAAACTTTC | 41 | 1–25 | 1157 bp | NheI | - |
| 4 | TARV-S3 R | ATCGctcgagTTAAGCGTAATCTGGAACATCGTATGGGTACCAACCACACTCCATAAAAGTCAG | 64 | 1101–1078 | XhoI | HA |
| Serial Number of Recombinant PICV-Based TARV Vaccines | Strains of Turkey Arthritis Reovirus | Recombinant PICV-Based TARV Vaccine Type | Insert in Plasmid 1 pP18S1-GPC/MCS | Insert in Plasmid 2 pP18S2-MCS/NP |
|---|---|---|---|---|
| 1 | SKM73 * | Monovalent | GFP a | S1 + wild-type |
| 2 | Monovalent | S3 + wild-type | GFP | |
| 3 | Bivalent | S3 wild-type | S1 wild-type | |
| 4 | SKM95 * | Monovalent | GFP | S1 wild-type |
| 5 | Monovalent | S3 wild-type | GFP | |
| 6 | Bivalent | S3 wild-type | S1 wild-type | |
| 7 | SKM121 * | Monovalent | GFP | S1 wild-type |
| 8 | Monovalent | S3 wild-type | GFP | |
| 9 | Bivalent | S3 wild-type | S1 wild-type | |
| 10 | SKM121 * | Monovalent | GFP | S1 codon-optimized |
| 11 | Monovalent | S3 codon-optimized | GFP | |
| 12 | Bivalent | S3 codon-optimized | S1 codon-optimized | |
| 13 | Control | GFP | GFP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, P.; Sharafeldin, T.A.; Kumar, R.; Huang, Q.; Liang, Y.; Goyal, S.M.; Porter, R.E.; Ly, H.; Mor, S.K. Development of a Recombinant Pichinde Virus-Vectored Vaccine against Turkey Arthritis Reovirus and Its Immunological Response Characterization in Vaccinated Animals. Pathogens 2021, 10, 197. https://doi.org/10.3390/pathogens10020197
Kumar P, Sharafeldin TA, Kumar R, Huang Q, Liang Y, Goyal SM, Porter RE, Ly H, Mor SK. Development of a Recombinant Pichinde Virus-Vectored Vaccine against Turkey Arthritis Reovirus and Its Immunological Response Characterization in Vaccinated Animals. Pathogens. 2021; 10(2):197. https://doi.org/10.3390/pathogens10020197
Chicago/Turabian StyleKumar, Pawan, Tamer A. Sharafeldin, Rahul Kumar, Qinfeng Huang, Yuying Liang, Sagar M. Goyal, Robert E. Porter, Hinh Ly, and Sunil K. Mor. 2021. "Development of a Recombinant Pichinde Virus-Vectored Vaccine against Turkey Arthritis Reovirus and Its Immunological Response Characterization in Vaccinated Animals" Pathogens 10, no. 2: 197. https://doi.org/10.3390/pathogens10020197