
Role of Extracellular Mycobacteria in Blood-Retinal Barrier Invasion in a Zebrafish Model of Ocular TB


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
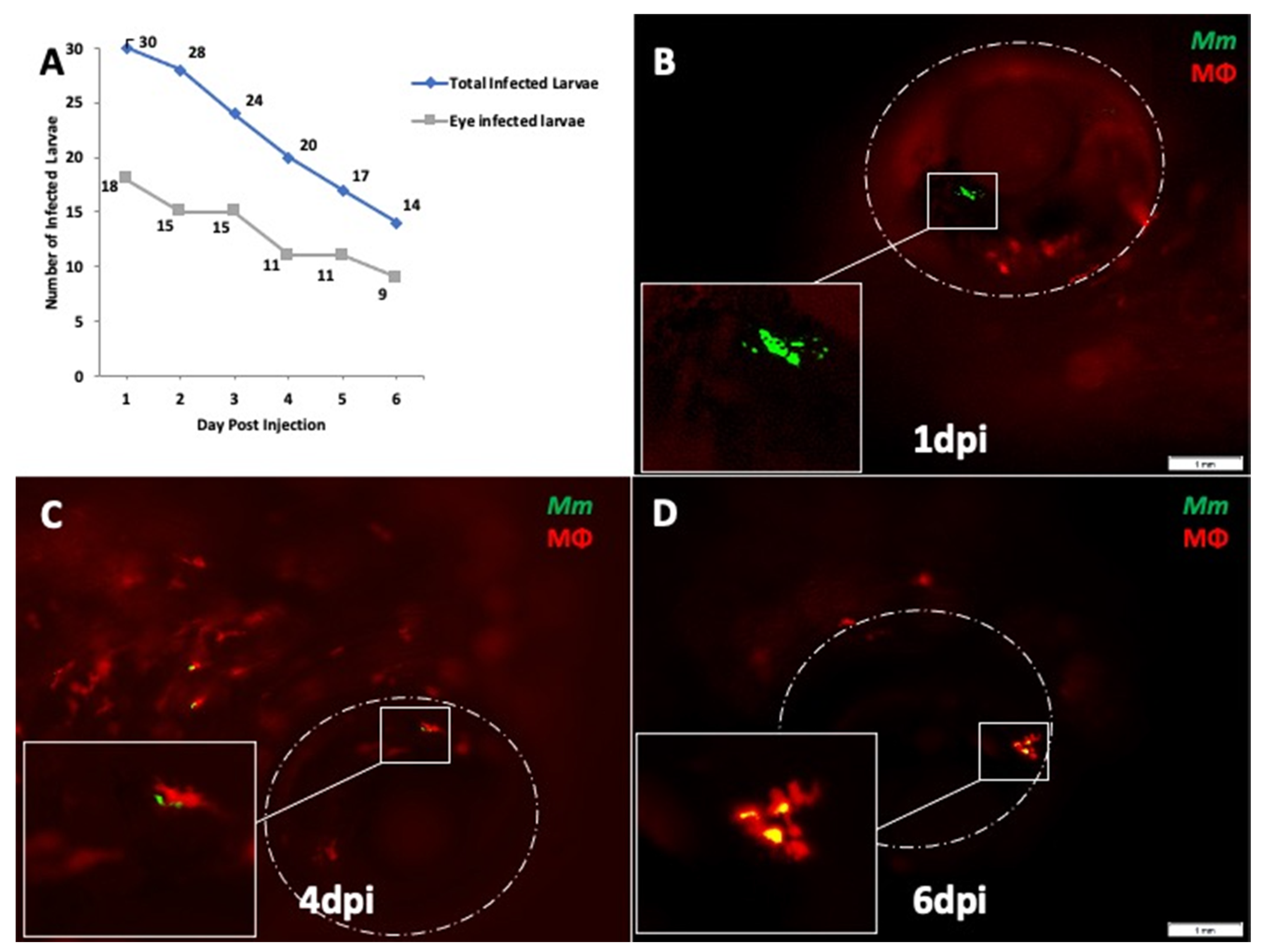
2.1. Mm Causes High Rate of Ocular Infection Even with Low Inoculum of Systemic Infection
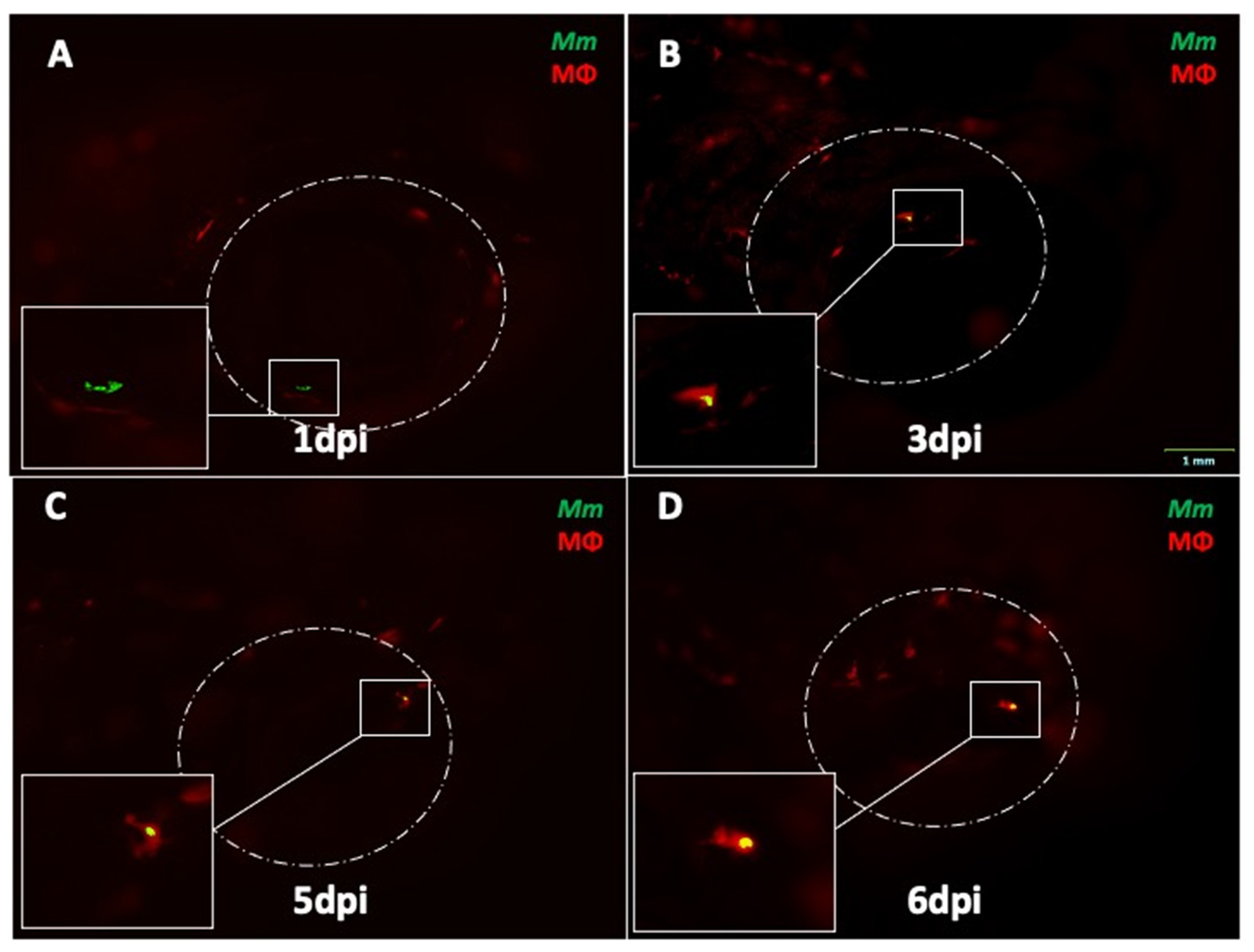
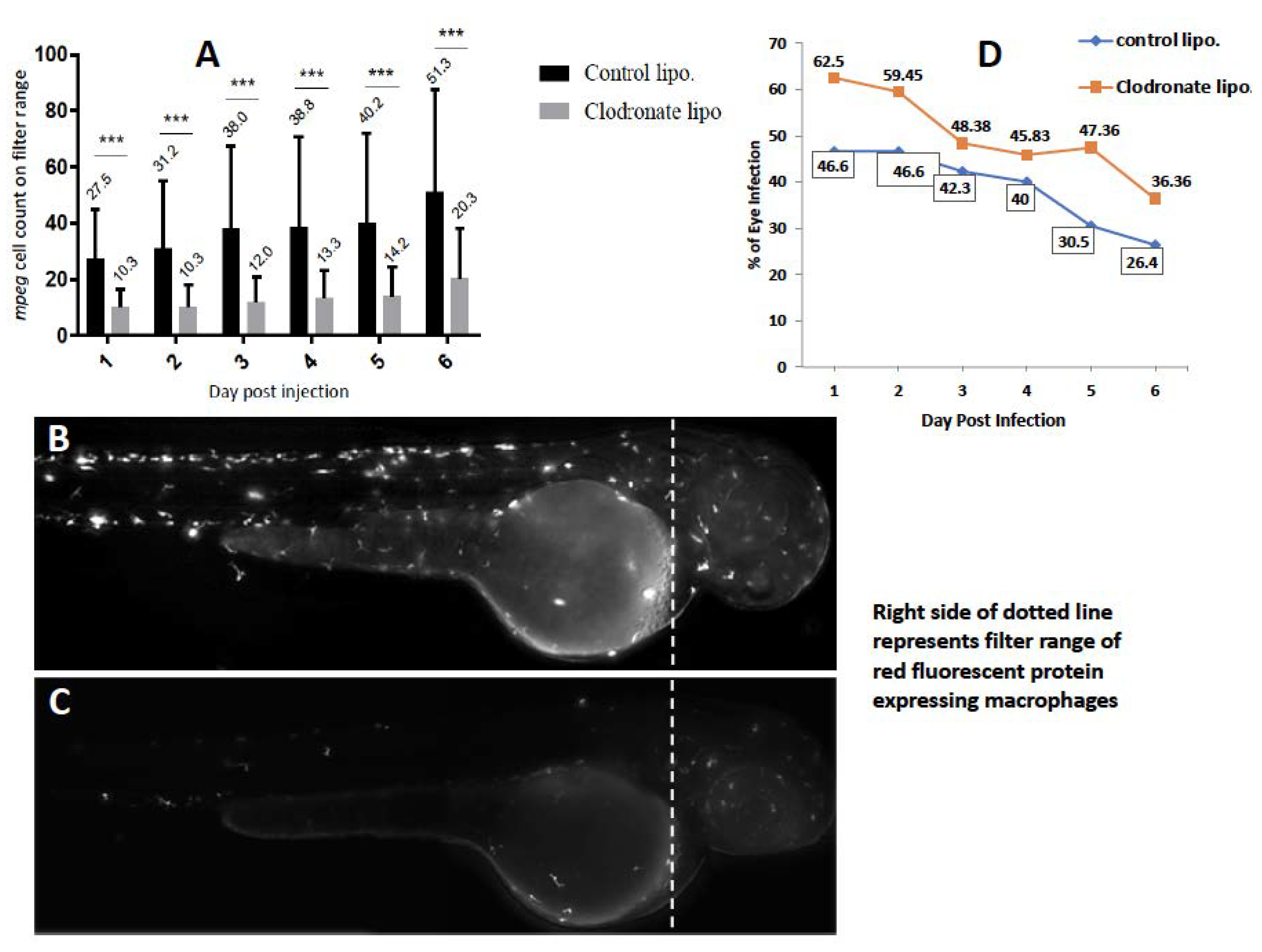
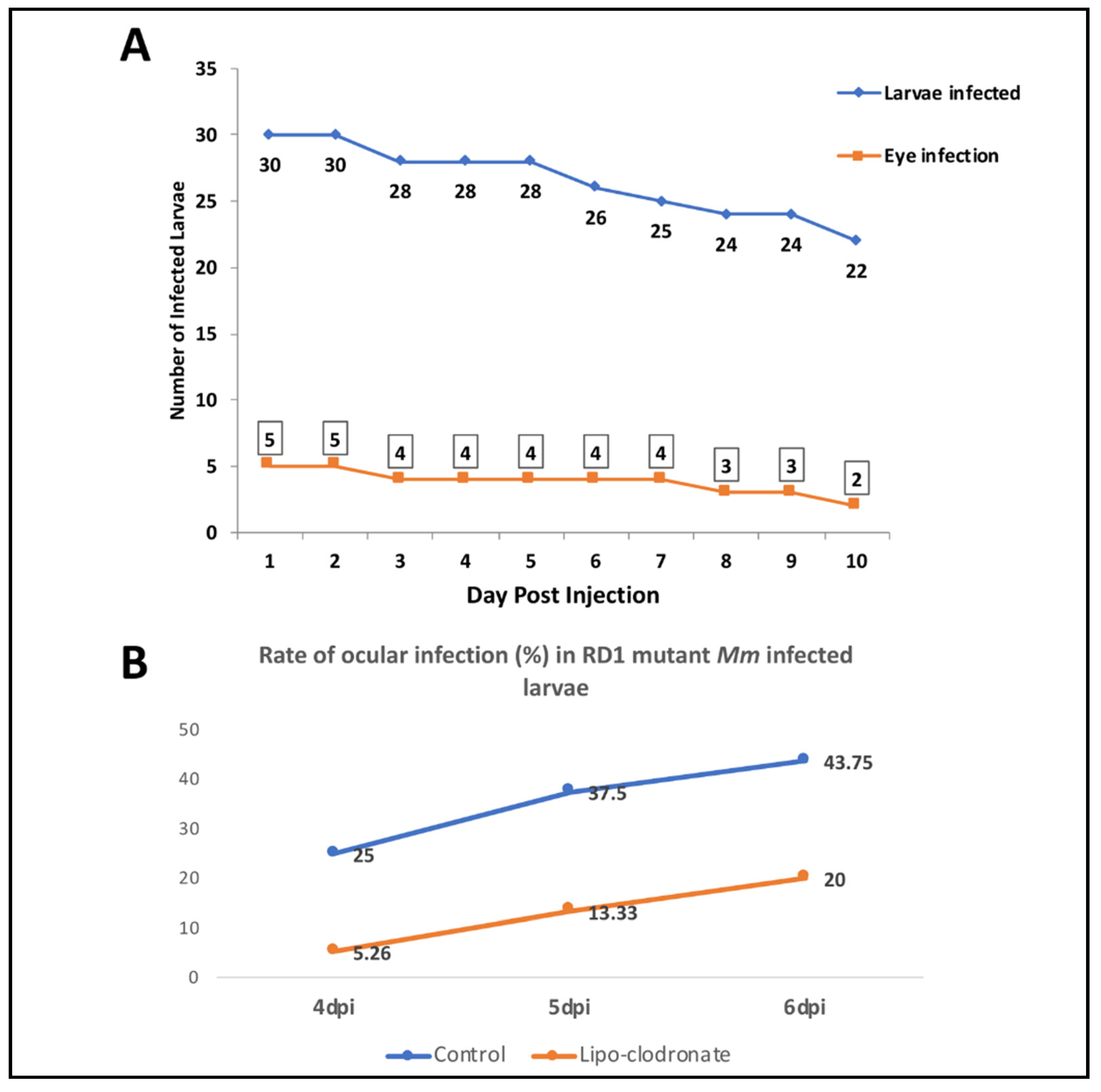
2.2. Depletion of Circulating Monocytes Increases Rate of Ocular Infection
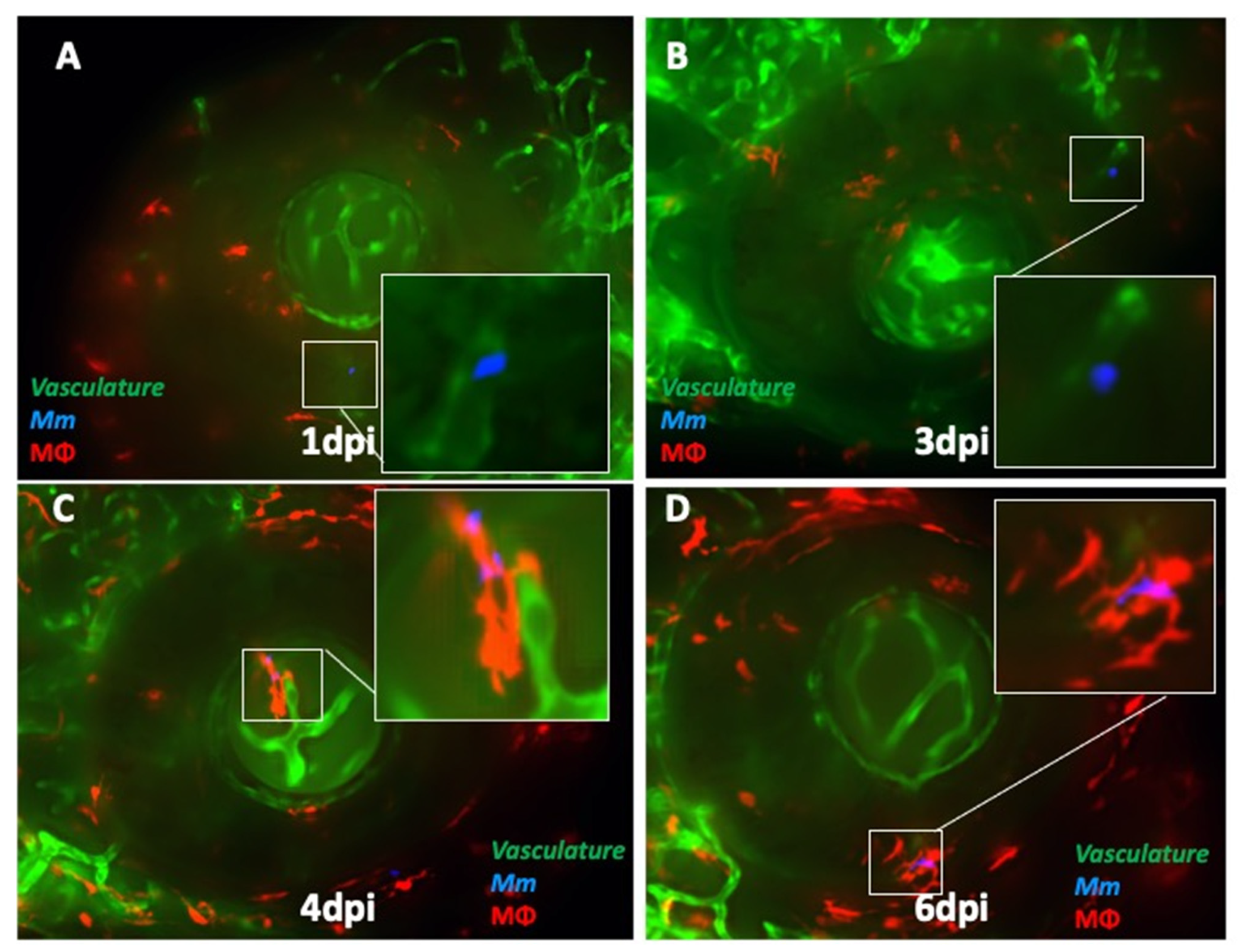
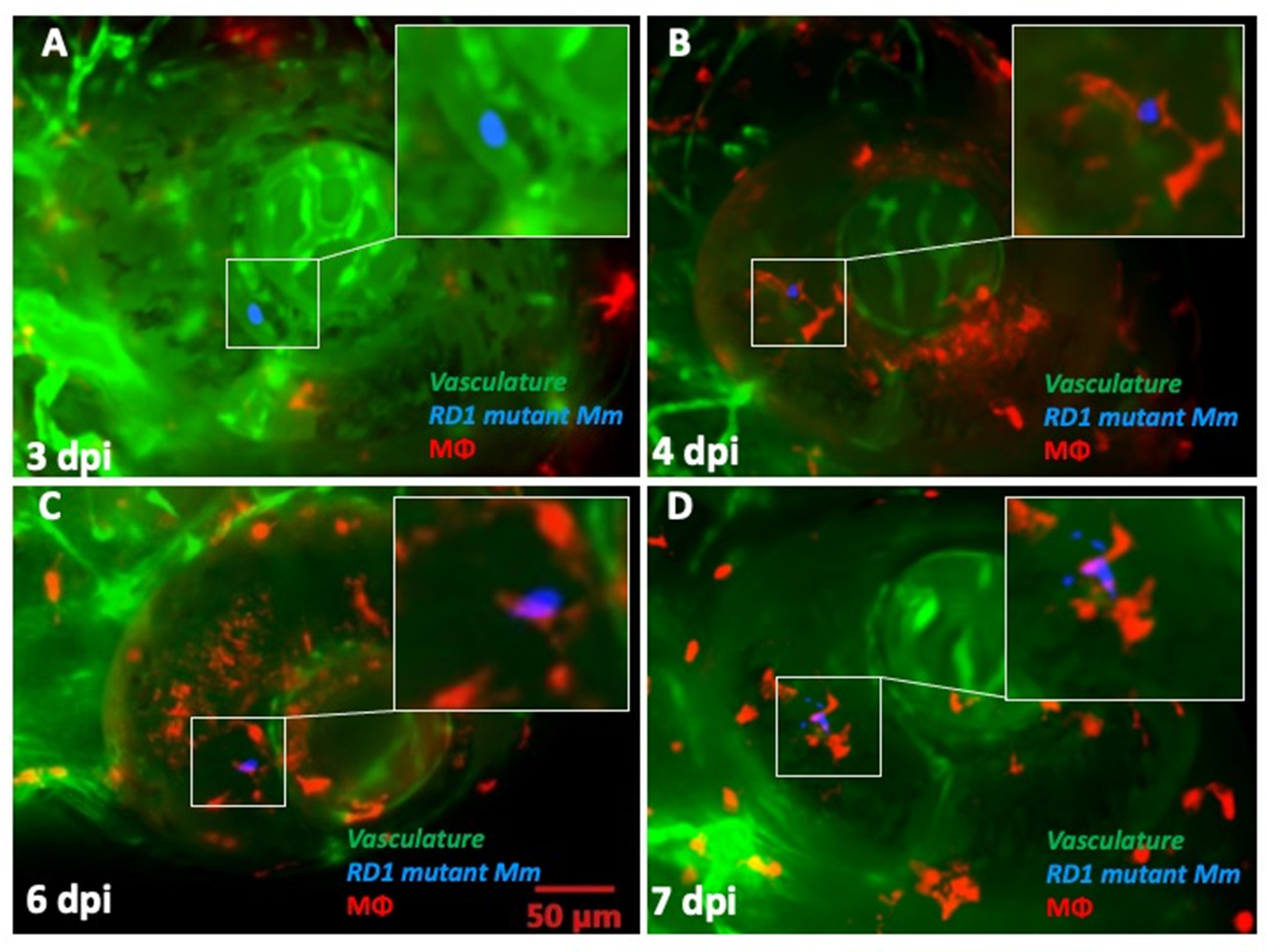
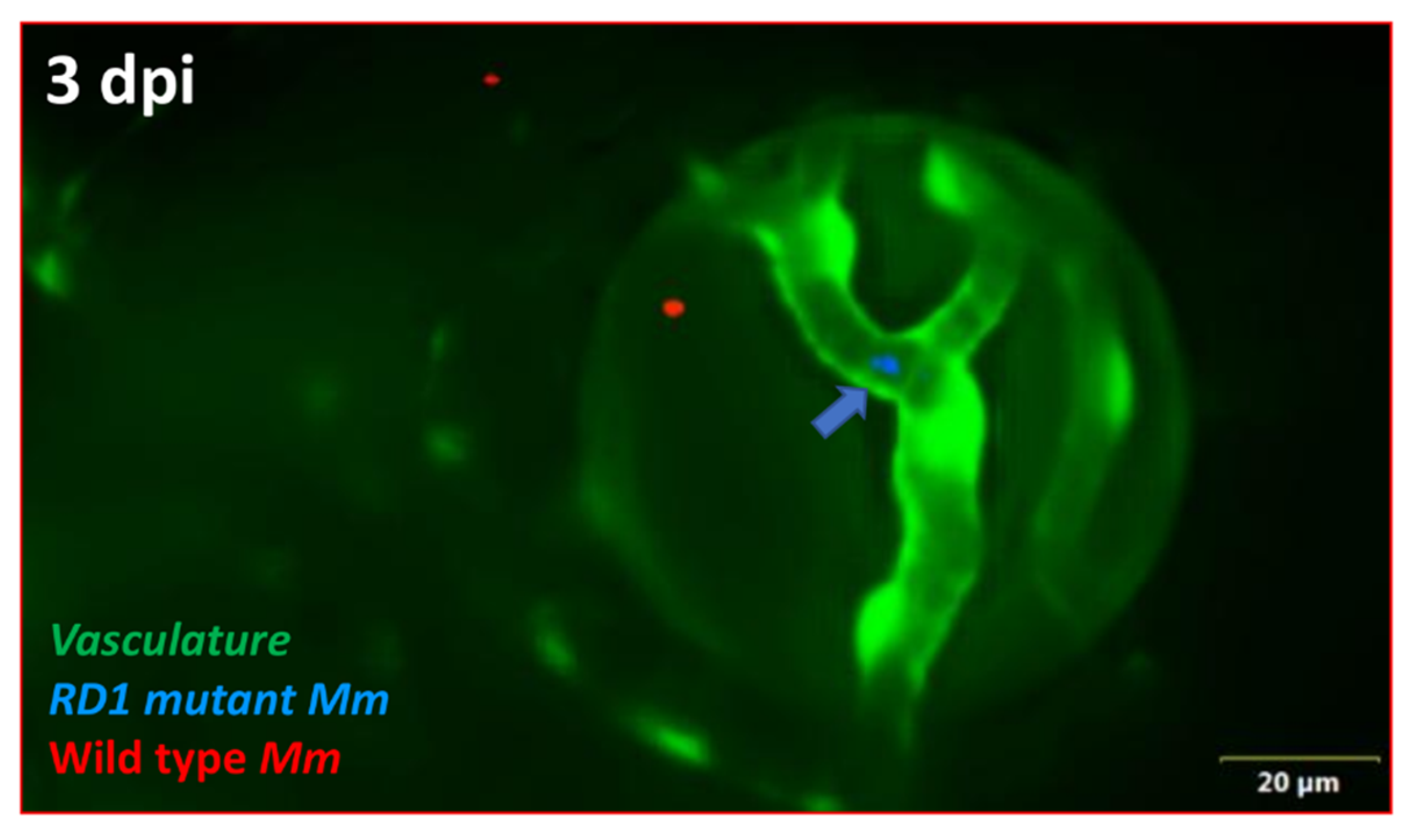
2.3. ESX-1 Secretion System Is Required for BRB Traversal and Granuloma Formation
3. Discussion
4. Methods
4.1. Zebrafish Husbandry
4.2. Bacterial Strains
4.3. Caudal Vein Injections
4.4. Microscopy
4.5. Depletion of Circulating Monocytes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dogra, M.; Singh, R.; Agarwal, A.; Sharma, A.; Singh, S.R.; Gautam, N.; Yangzes, S.; Samanta, R.; Sharma, M.; Aggarwal, K.; et al. Epidemiology of Uveitis in a Tertiary-care Referral Institute in North India. Ocul. Immunol. Inflamm. 2017, 25, S46–S53. [Google Scholar] [CrossRef]
- Gan, W.L.; Jones, N.P. Serpiginous-like choroiditis as a marker for tuberculosis in a non-endemic area. Br. J. Ophthalmol. 2013, 97, 644–647. [Google Scholar] [CrossRef]
- Basu, S.; Wakefield, D.; Biswas, J.; Rao, N.A. Pathogenesis and Pathology of Intraocular Tuberculosis. Ocul. Immunol. Inflamm. 2015, 23, 353–357. [Google Scholar] [CrossRef]
- Biswas, J.; Madhavan, H.N.; Gopal, L.; Badrinath, S.S. Intraocular tuberculosis. Clinicopathologic study of five cases. Retina 1995, 15, 461–468. [Google Scholar] [CrossRef]
- Wroblewski, K.J.; Hidayat, A.A.; Neafie, R.C.; Rao, N.A.; Zapor, M. Ocular Tuberculosis: A Clinicopathologic and Molecular Study. Ophthalmology 2011, 118, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Cadena, A.M.; Flynn, J.L.; Fortune, S.M. The Importance of First Impressions: Early Events in Mycobacterium tuberculosis Infection Influence Outcome. mBio 2016, 7, e00342-16. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.; Clay, H.; Lewis, J.L.; Ghori, N.; Herbomel, P.; Ramakrishnan, L. Real-Time Visualization of Mycobacterium-Macrophage Interactions Leading to Initiation of Granuloma Formation in Zebrafish Embryos. Immunity 2002, 17, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Takaki, K.; Davis, J.M.; Winglee, K.; Ramakrishnan, L. Evaluation of the pathogenesis and treatment of Mycobacterium marinum infection in zebrafish. Nat. Protoc. 2013, 8, 1114–1124. [Google Scholar] [CrossRef] [Green Version]
- Cronan, M.R.; Tobin, D.M. Fit for consumption: Zebrafish as a model for tuberculosis. Dis. Model. Mech. 2014, 7, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, L. The Zebrafish Guide to Tuberculosis Immunity and Treatment. Cold Spring Harb. Symp. Quant. Biol. 2013, 78, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, L.M.; Van Der Kuip, M.; Youssef, S.A.; De Bruin, A.; Bitter, W.; Van Furth, A.M.; Van Der Sar, A.M. Modeling tuberculous meningitis in zebrafish using Mycobacterium marinum. Dis. Model. Mech. 2014, 7, 1111–1122. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, L.M.; Boot, M.; Kuijl, C.; Picavet, D.I.; van Stempvoort, G.; van der Pol, S.M.; de Vries, H.E.; van der Wel, N.N.; van der Kuip, M.; van Furth, A.M.; et al. Mycobacteria employ two different mechanisms to cross the blood–brain barrier. Cell. Microbiol. 2018, 20, e12858. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Shao, X.-Y.; Wang, C.; Hua, M.-H.; Wang, C.-N.; Wang, X.; Wang, Q.-J.; Yao, J.-Y.; Fan, Y.-H.; Qin, Y.-W. Mycobacterium marinum Infection in Zebrafish and Microglia Imitates the Early Stage of Tuberculous Meningitis. J. Mol. Neurosci. 2018, 64, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Tenor, J.L.; Oehlers, S.H.; Yang, J.L.; Tobin, D.M.; Perfect, J.R. Live Imaging of Host-Parasite Interactions in a Zebrafish Infection Model Reveals Cryptococcal Determinants of Virulence and Central Nervous System Invasion. mBio 2015, 6, e01425-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaki, K.; Ramakrishnan, L.; Basu, S. A zebrafish model for ocular tuberculosis. PLoS ONE 2018, 13, e0194982. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Farage, E.; Sugimoto, M.; Anand-Apte, B. A novel transgenic zebrafish model for blood-brain and blood-retinal barrier development. BMC Dev. Biol. 2010, 10, 76. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.-Y.; Kwon, H.-B.; Ahn, J.-C.; Kang, D.; Kwon, S.-H.; Park, J.A.; Kim, K.-W. Functional and developmental analysis of the blood–brain barrier in zebrafish. Brain Res. Bull. 2008, 75, 619–628. [Google Scholar] [CrossRef]
- Santiago-Tirado, F.H.; Onken, M.D.; Cooper, J.A.; Klein, R.S.; Doering, T.L. Trojan Horse Transit Contributes to Blood-Brain Barrier Crossing of a Eukaryotic Pathogen. mBio 2017, 8, e02183-16. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.K.; Paul-Satyaseela, M.; Lamichhane, G.; Kim, K.S.; Bishai, W.R. Mycobacterium tuberculosisInvasion and Traversal across an In Vitro Human Blood-Brain Barrier as a Pathogenic Mechanism for Central Nervous System Tuberculosis. J. Infect. Dis. 2006, 193, 1287–1295. [Google Scholar] [CrossRef] [Green Version]
- Dobos, K.M.; Spotts, E.A.; Quinn, F.D.; King, C.H. Necrosis of Lung Epithelial Cells during Infection withMycobacterium tuberculosis Is Preceded by Cell Permeation. Infect. Immun. 2000, 68, 2315–2322. [Google Scholar] [CrossRef]
- Ryndak, M.B.; Singh, K.K.; Peng, Z.; Laal, S. Transcriptional Profile of Mycobacterium tuberculosis Replicating in Type II Alveolar Epithelial Cells. PLoS ONE 2015, 10, e0123745. [Google Scholar] [CrossRef]
- Ryndak, M.B.; Laal, S. Mycobacterium tuberculosis Primary Infection and Dissemination: A Critical Role for Alveolar Epithelial Cells. Front. Cell. Infect. Microbiol. 2019, 9, 299. [Google Scholar] [CrossRef]
- McDonald, L.C.; Archibald, L.K.; Rheanpumikankit, S.; Tansuphaswadikul, S.; Eampokalap, B.; Nwanyanwu, O.; Kazembe, P.; Dobbie, H.; Reller, L.B.; Jarvis, W.R. Unrecognised Mycobacterium tuberculosis bacteraemia among hospital inpatients in less developed countries. Lancet 1999, 354, 1159–1163. [Google Scholar] [CrossRef]
- Lerner, T.R.; Carvalho-Wodarz, C.D.S.; Repnik, U.; Russell, M.R.; Borel, S.; Diedrich, C.R.; Rohde, M.; Wainwright, H.; Collinson, L.M.; Wilkinson, R.J.; et al. Lymphatic endothelial cells are a replicative niche for Mycobacterium tuberculosis. J. Clin. Investig. 2016, 126, 1093–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkington, P.T.; Friedland, J.S. Permutations of time and place in tuberculosis. Lancet Infect. Dis. 2015, 15, 1357–1360. [Google Scholar] [CrossRef] [Green Version]
- Clay, H.; Davis, J.M.; Beery, D.; Huttenlocher, A.; Lyons, S.E.; Ramakrishnan, L. Dichotomous Role of the Macrophage in Early Mycobacterium marinum Infection of the Zebrafish. Cell Host Microbe 2007, 2, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Dannenberg, A.M., Jr. Immunopathogenesis of Pulmonary Tuberculosis. Hosp. Pract. 1993, 28, 51–58. [Google Scholar] [CrossRef]
- Davis, J.M.; Ramakrishnan, L. The Role of the Granuloma in Expansion and Dissemination of Early Tuberculous Infection. Cell 2009, 136, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Elkington, P.; Rao, N.A. Pathogenesis of ocular tuberculosis: New observations and future directions. Tuberculosis 2020, 124, 101961. [Google Scholar] [CrossRef]
- Abhishek, S.; Saikia, U.N.; Gupta, A.; Bansal, R.; Gupta, V.; Singh, N.; Laal, S.; Verma, I. Transcriptional Profile of Mycobacterium tuberculosis in an in vitro Model of Intraocular Tuberculosis. Front. Cell. Infect. Microbiol. 2018, 8, 330. [Google Scholar] [CrossRef] [PubMed]
- Nazari, H.; Karakousis, P.C.; Rao, N.A. Replication of Mycobacterium Tuberculosis in Retinal Pigment Epithelium. JAMA Ophthalmol. 2014, 132, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.T.; Cambier, C.J.; Davis, J.M.; Hall, C.J.; Crosier, P.S.; Ramakrishnan, L. Neutrophils exert protection in the early tuberculous gran-uloma by oxidative killing of mycobacteria phagocytosed from infected macrophages. Cell Host Microbe 2012, 12, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojarczuk, A.; Miller, K.A.; Hotham, R.; Lewis, A.; Ogryzko, N.V.; Kamuyango, A.A.; Frost, H.; Gibson, R.H.; Stillman, E.; May, R.C.; et al. Cryptococcus neoformans Intracellular Proliferation and Capsule Size Determines Early Macrophage Control of Infection. Sci. Rep. 2016, 6, 21489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernut, A.; Herrmann, J.L.; Kissa, K.; Dubremetz, J.-F.; Gaillard, J.-L.; Lutfalla, G.; Kremer, L. Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation. Proc. Natl. Acad. Sci. USA 2014, 111, E943–E952. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damera, S.K.; Panigrahi, R.K.; Mitra, S.; Basu, S. Role of Extracellular Mycobacteria in Blood-Retinal Barrier Invasion in a Zebrafish Model of Ocular TB. Pathogens 2021, 10, 333. https://doi.org/10.3390/pathogens10030333
Damera SK, Panigrahi RK, Mitra S, Basu S. Role of Extracellular Mycobacteria in Blood-Retinal Barrier Invasion in a Zebrafish Model of Ocular TB. Pathogens. 2021; 10(3):333. https://doi.org/10.3390/pathogens10030333
Chicago/Turabian StyleDamera, Santhosh Kumar, Ranjan Kumar Panigrahi, Sanchita Mitra, and Soumyava Basu. 2021. "Role of Extracellular Mycobacteria in Blood-Retinal Barrier Invasion in a Zebrafish Model of Ocular TB" Pathogens 10, no. 3: 333. https://doi.org/10.3390/pathogens10030333