Identification and Characterization of Plant-Interacting Targets of Tomato Spotted Wilt Virus Silencing Suppressor



Abstract
:1. Introduction
2. Results
2.1. Affinity Purification—Mass Spectrometry Analysis Reveals NSs-Interacting Proteins in N. benthamiana
2.2. Gene Ontology Overrepresentation/Enrichment Tests of NSs-Interacting Proteins
2.3. The Protein-Protein Interaction Network of NSs-Interacting Proteins
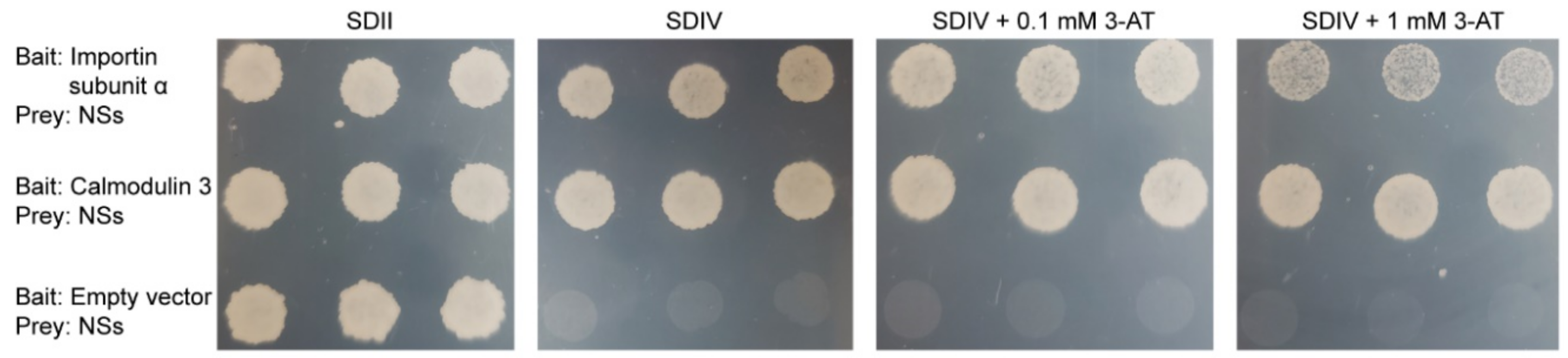
2.4. Importin Subunit α and Calmodulin 3 Interact with NSs in Targeted Yeast Two-Hybrid Assays
3. Discussion
4. Materials and Methods
4.1. Plasmids and Gene Cloning
4.2. Affinity Purification—Mass Spectrometry Analysis of NSs-Interacting Proteins
4.3. Refinement of the NSs-Interacting Protein Candidate List
4.4. Verification of NSs-Interacting Proteins by Targeted Yeast Two-Hybrid Assay
4.5. Gene Ontology Analysis of Inferred Tobacco, Tomato and Arabidopsis Homologs
4.6. Protein-Protein Interaction Network Analysis of Inferred Arabidopsis Homologs
4.7. Mass Spectrometry Data Deposit
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Siddell, S.G.; Walker, P.J.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; et al. Additional changes to taxonomy ratified in a special vote by the International Committee on Taxonomy of Viruses (October 2018). Arch. Virol. 2019, 164, 943–946. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; van Grinsven, I.L.; Kormelink, R.; Tao, X. Paving the way to tospovirus infection: Multilined interplays with plant innate immunity. Annu. Rev. Phytopathol. 2019, 57, 41–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adkins, S. Tomato spotted wilt virus—Positive steps towards negative success. Mol. Plant Pathol. 2000, 1, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Pappu, H.R.; Jones, R.A.; Jain, R.K. Global status of tospovirus epidemics in diverse cropping systems: Successes achieved and challenges ahead. Virus Res. 2009, 141, 219–236. [Google Scholar] [CrossRef]
- Oliver, J.E.; Whitfield, A.E. The genus Tospovirus: Emerging Bunyaviruses that threaten food security. Annu. Rev. Virol. 2016, 3, 101–124. [Google Scholar] [CrossRef]
- Zhai, Y.; Bag, S.; Mitter, N.; Turina, M.; Pappu, H.R. Mutational analysis of two highly conserved motifs in the silencing suppressor encoded by Tomato spotted wilt virus (genus Tospovirus, family Bunyaviridae). Arch. Virol. 2014, 159, 1499–1504. [Google Scholar] [CrossRef]
- Zhai, Y.; Peng, H.; Neff, M.M.; Pappu, H.R. Putative auxin and light responsive promoter elements from the Tomato spotted wilt tospovirus genome, when expressed as cDNA, are functional in Arabidopsis. Front. Plant Sci. 2019, 10, 804. [Google Scholar] [CrossRef]
- Turina, M.; Kormelink, R.; Resende, R.O. Resistance to tospoviruses in vegetable crops: Epidemiological and molecular aspects. Annu. Rev. Phytopathol. 2016, 54, 347–371. [Google Scholar] [CrossRef]
- Hedil, M.; Kormelink, R. Viral RNA silencing suppression: The enigma of Bunyavirus NSs proteins. Viruses 2016, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Hedil, M.; de Ronde, D.; Kormelink, R. Biochemical analysis of NSs from different tospoviruses. Virus Res. 2017, 242, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Sugiyama, K.; Nagano, H.; Mori, M.; Kaido, M.; Mise, K.; Tsuda, S.; Okuno, T. Identification of a novel RNA silencing suppressor, NSs protein of Tomato spotted wilt virus. FEBS Lett. 2002, 532, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Margaria, P.; Ciuffo, M.; Pacifico, D.; Turina, M. Evidence that the nonstructural protein of Tomato spotted wilt virus is the avirulence determinant in the interaction with resistant pepper carrying the Tsw gene. Mol. Plant Microbe Interact. 2007, 20, 547–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ronde, D.; Butterbach, P.; Lohuis, D.; Hedil, M.; van Lent, J.W.; Kormelink, R. Tsw gene-based resistance is triggered by a functional RNA silencing suppressor protein of the Tomato spotted wilt virus. Mol. Plant Pathol. 2013, 14, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Almási, A.; Nemes, K.; Csömör, Z.; Tóbiás, I.; Palkovics, L.; Salánki, K. A single point mutation in Tomato spotted wilt virus NSs protein is sufficient to overcome Tsw-gene-mediated resistance in pepper. J. Gen. Virol. 2017, 98, 1521–1525. [Google Scholar] [CrossRef] [PubMed]
- De Ronde, D.; Pasquier, A.; Ying, S.; Butterbach, P.; Lohuis, D.; Kormelink, R. Analysis of Tomato spotted wilt virus NSs protein indicates the importance of the N-terminal domain for avirulence and RNA silencing suppression. Mol. Plant Pathol. 2014, 15, 185–195. [Google Scholar] [CrossRef]
- Zhao, L.; Hu, Z.; Li, S.; Zhou, X.; Li, J.; Su, X.; Zhang, L.; Zhang, Z.; Dong, J. Diterpenoid compounds from Wedelia trilobata induce resistance to Tomato spotted wilt virus via the JA signal pathway in tobacco plants. Sci. Rep. 2019, 9, 2763. [Google Scholar] [CrossRef]
- Wu, X.; Xu, S.; Zhao, P.; Zhang, X.; Yao, X.; Sun, Y.; Fang, R.; Ye, J. The Orthotospovirus nonstructural protein NSs suppresses plant MYC-regulated jasmonate signaling leading to enhanced vector attraction and performance. PLoS Pathog. 2019, 15, e1007897. [Google Scholar] [CrossRef]
- Thomas, E.L.; van der Hoorn, R.A.L. Ten prominent host proteases in plant-pathogen interactions. Int. J. Mol. Sci. 2018, 19, 639. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Win, J.; Song, J.; van der Hoorn, R.; van der Knaap, E.; Kamoun, S. A Phytophthora infestans cystatin-like protein targets a novel tomato papain-like apoplastic protease. Plant Physiol. 2007, 143, 364–377. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Ismayil, A.; Liu, Y. Autophagy in plant-virus interactions. Annu. Rev. Virol. 2020, 7, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Cheval, C.; Aldon, D.; Galaud, J.P.; Ranty, B. Calcium/calmodulin-mediated regulation of plant immunity. Biochim. Biophys. Acta 2013, 1833, 1766–1771. [Google Scholar] [CrossRef] [Green Version]
- Chiasson, D.; Ekengren, S.K.; Martin, G.B.; Dobney, S.L.; Snedden, W.A. Calmodulin-like proteins from Arabidopsis and tomato are involved in host defense against Pseudomonas syringae pv. Tomato. Plant Mol. Biol. 2005, 58, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Takabatake, R.; Karita, E.; Seo, S.; Mitsuhara, I.; Kuchitsu, K.; Ohashi, Y. Pathogen-induced calmodulin isoforms in basal resistance against bacterial and fungal pathogens in tobacco. Plant Cell Physiol. 2007, 48, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahara, K.S.; Masuta, C.; Yamada, S.; Shimura, H.; Kashihara, Y.; Wada, T.S.; Meguro, A.; Goto, K.; Tadamura, K.; Sueda, K.; et al. Tobacco calmodulin-like protein provides secondary defense by binding to and directing degradation of virus RNA silencing suppressors. Proc. Natl. Acad. Sci. USA 2012, 109, 10113–10118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Huang, C.; Li, Z.; Zhou, X. Suppression of RNA silencing by a plant DNA virus satellite requires a host calmodulin-like protein to repress RDR6 expression. PLoS Pathog. 2014, 10, e1003921. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Lee, L.Y.; Oltmanns, H.; Cao, H.; Veena; Cuperus, J.; Gelvin, S.B. IMPa-4, an Arabidopsis importin α isoform, is preferentially involved in Agrobacterium-mediated plant transformation. Plant Cell 2008, 20, 2661–2680. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.H.; Lacatus, G.; Sunter, G. Geminivirus AL2 protein induces expression of, and interacts with, a calmodulin-like gene, an endogenous regulator of gene silencing. Virology 2014, 460–461, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Zhao, N.; Li, Z.; Xu, X.; Wang, Y.; Yang, X.; Liu, S.S.; Wang, A.; Zhou, X. A calmodulin-like protein suppresses RNA silencing and promotes geminivirus infection by degrading SGS3 via the autophagy pathway in Nicotiana benthamiana. PLoS Pathog. 2017, 13, e1006213. [Google Scholar] [CrossRef]
- Guerra-Peraza, O.; Kirk, D.; Seltzer, V.; Veluthambi, K.; Schmit, A.C.; Hohn, T.; Herzog, E. Coat proteins of Rice tungro bacilliform virus and Mungbean yellow mosaic virus contain multiple nuclear-localization signals and interact with importin alpha. J. Gen. Virol. 2005, 86, 1815–1826. [Google Scholar] [CrossRef]
- Haas, G.; Azevedo, J.; Moissiard, G.; Geldreich, A.; Himber, C.; Bureau, M.; Fukuhara, T.; Keller, M.; Voinnet, O. Nuclear import of CaMV P6 is required for infection and suppression of the RNA silencing factor DRB4. EMBO J. 2008, 27, 2102–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, X.; Niu, S.; Han, C.; Yu, J.; Li, D. Nuclear localization of Beet black scorch virus capsid protein and its interaction with importin α. Virus Res. 2011, 155, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Lukhovitskaya, N.I.; Cowan, G.H.; Vetukuri, R.R.; Tilsner, J.; Torrance, L.; Savenkov, E.I. Importin-α-mediated nucleolar localization of Potato mop-top virus TRIPLE GENE BLOCK1 (TGB1) protein facilitates virus systemic movement, whereas TGB1 self-interaction is required for cell-to-cell movement in Nicotiana benthamiana. Plant Physiol. 2015, 167, 738–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Cañamás, M.; Hernández, C. New insights into the nucleolar localization of a plant RNA virus-encoded protein that acts in both RNA packaging and RNA silencing suppression: Involvement of importins α and relevance for viral infection. Mol. Plant Microbe Interact. 2018, 31, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, D.; Chakraborty, S. Chloroplast: The Trojan horse in plant-virus interaction. Mol. Plant Pathol. 2018, 19, 504–518. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Peng, H.; Neff, M.M.; Pappu, H.R. Emerging molecular links between plant photomorphogenesis and virus resistance. Front. Plant Sci. 2020, 11, 920. [Google Scholar] [CrossRef]
- Slaymaker, D.H.; Navarre, D.A.; Clark, D.; del Pozo, O.; Martin, G.B.; Klessig, D.F. The tobacco salicylic acid-binding protein 3 (SABP3) is the chloroplast carbonic anhydrase, which exhibits antioxidant activity and plays a role in the hypersensitive defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 11640–11645. [Google Scholar] [CrossRef] [Green Version]
- Restrepo, S.; Myers, K.L.; del Pozo, O.; Martin, G.B.; Hart, A.L.; Buell, C.R.; Fry, W.E.; Smart, C.D. Gene profiling of a compatible interaction between Phytophthora infestans and Solanum tuberosum suggests a role for carbonic anhydrase. Mol. Plant Microbe Interact. 2005, 18, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Kravats, A.N.; Hoskins, J.R.; Reidy, M.; Johnson, J.L.; Doyle, S.M.; Genest, O.; Masison, D.C.; Wickner, S. Functional and physical interaction between yeast Hsp90 and Hsp70. Proc. Natl. Acad. Sci. USA 2018, 115, E2210–E2219. [Google Scholar] [CrossRef] [Green Version]
- Hahn, A.; Bublak, D.; Schleiff, E.; Scharf, K.D. Crosstalk between Hsp90 and Hsp70 chaperones and heat stress transcription factors in tomato. Plant Cell 2011, 23, 741–755. [Google Scholar] [CrossRef] [Green Version]
- Aparicio, F.; Thomas, C.L.; Lederer, C.; Niu, Y.; Wang, D.; Maule, A.J. Virus induction of heat shock protein 70 reflects a general response to protein accumulation in the plant cytosol. Plant Physiol. 2005, 138, 529–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorovits, R.; Moshe, A.; Ghanim, M.; Czosnek, H. Recruitment of the host plant heat shock protein 70 by Tomato yellow leaf curl virus coat protein is required for virus infection. PLoS ONE 2013, 8, e70280. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Malcuit, I.; Moffett, P.; Ruiz, M.T.; Peart, J.; Wu, A.J.; Rathjen, J.P.; Bendahmane, A.; Day, L.; Baulcombe, D.C. High throughput virus-induced gene silencing implicates heat shock protein 90 in plant disease resistance. EMBO J. 2003, 22, 5690–5699. [Google Scholar] [CrossRef] [PubMed]
- Earley, K.W.; Haag, J.R.; Pontes, O.; Opper, K.; Juehne, T.; Song, K.; Pikaard, C.S. Gateway-compatible vectors for plant functional genomics and proteomics. Plant J. 2006, 45, 616–629. [Google Scholar] [CrossRef]
- Wang, L.; Ding, X.; Xiao, J.; Jiménez-Gόngora, T.; Liu, R.; Lozano-Durán, R. Inference of a geminivirus-host protein-protein interaction network through affinity purification and mass spectrometry analysis. Viruses 2017, 9, 275. [Google Scholar] [CrossRef]
- Sang, Y.; Wang, Y.; Ni, H.; Cazalé, A.C.; She, Y.M.; Peeters, N.; Macho, A.P. The Ralstonia solanacearum type III effector RipAY targets plant redox regulators to suppress immune responses. Mol. Plant Pathol. 2018, 19, 129–142. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Brodbelt, J.S.; Russell, D.H. Focus on the 20-year anniversary of SEQUEST. J. Am. Soc. Mass Spectrom. 2015, 26, 1797–1798. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Neff, M.M. Circadian Clock Associated 1 and ATAF2 differentially suppress cytochrome P450-mediated brassinosteroid inactivation. J. Exp. Bot. 2020, 71, 970–985. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Geisler-Lee, J.; O’Toole, N.; Ammar, R.; Provart, N.J.; Millar, A.H.; Geisler, M. A predicted interactome for Arabidopsis. Plant Physiol. 2007, 145, 317–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oughtred, R.; Stark, C.; Breitkreutz, B.J.; Rust, J.; Boucher, L.; Chang, C.; Kolas, N.; O’Donnell, L.; Leung, G.; McAdam, R.; et al. The BioGRID interaction database: 2019 update. Nucleic Acids Res. 2019, 47, D529–D541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerrien, S.; Aranda, B.; Breuza, L.; Bridge, A.; Broackes-Carter, F.; Chen, C.; Duesbury, M.; Dumousseau, M.; Feuermann, M.; Hinz, U.; et al. The IntAct molecular interaction database in 2012. Nucleic Acids Res. 2012, 40, D841–D846. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Accession | Description | #PSMs | Sum PEP Scores | Scores SEQUEST HT | |||
|---|---|---|---|---|---|---|---|
| R1 | R2 | R1 | R2 | R1 | R2 | ||
| A0A286RNF7 | Carbonic anhydrase | 33 | 28 | 65.105 | 67.837 | 86.19 | 71.41 |
| A0A0M3SBS3 | Heat shock protein 90-3 | 28 | 26 | 50.052 | 58.461 | 63.66 | 48.87 |
| A4D0J9 | Carbonic anhydrase (fragment) | 19 | 22 | 49.892 | 67.834 | 53.63 | 68.49 |
| I3QHX5 | Adenosylhomocysteinase | 15 | 9 | 23.165 | 17.085 | 27.23 | 11.48 |
| I0B7J2 | Chloroplast photosystem II subunit O2 (PSBO2) | 12 | 13 | 25.788 | 36.984 | 35.48 | 25.76 |
| I0B7J5 | Chloroplast photosystem II subunit P1 (PSBP1) | 10 | 9 | 29.376 | 31.322 | 28.37 | 29.27 |
| U5PY93 | MP-Interacting Protein (MIP) 1.2 | 10 | 6 | 18.593 | 13.573 | 23.1 | 12.96 |
| Q769C6 | Heat shock protein 70 (fragment) | 9 | 4 | 9.018 | 7.495 | 18.44 | 6.57 |
| U3MY90 | Proteinase inhibitor (fragment) | 8 | 10 | 19.765 | 31.157 | 20.94 | 26 |
| A0A0A7EAV4 | Ankyrin repeat containing protein 2 (AKR2) | 6 | 2 | 10.561 | 4.414 | 12.68 | 2.11 |
| F2Z9R2 | Glucose-6-phosphate 1-dehydrogenase (G6PD) | 6 | 2 | 7.83 | 2.791 | 10.65 | 1.79 |
| A1YUL9 | Importin subunit α | 5 | 5 | 13.326 | 9.718 | 17.15 | 6.01 |
| A0A0C4Y3N1 | RabG3c protein | 5 | 6 | 7.097 | 6.486 | 8.22 | 2.17 |
| A0A0S3ANR1 | NB-LRR HR-associated cell death (NRC) 2a | 5 | 3 | 6.318 | 2.992 | 1.87 | 1.63 |
| Q5YLB4 | DNA gyrase subunit B | 4 | 1 | 4.471 | 0.71 | 2.38 | 0 |
| U3MW48 | Calmodulin 3 (fragment) | 3 | 1 | 8.901 | 2.829 | 10.37 | 2.46 |
| Q5XPZ0 | Adenosine kinase (fragment) | 3 | 3 | 5.692 | 5.426 | 7.01 | 4.12 |
| R4S2V6 | Lipoxygenase (fragment) | 3 | 1 | 3.467 | 0.731 | 2.21 | 0 |
| A0A387K491 | Ran binding protein RanBP1-1b | 3 | 1 | 2.618 | 1.051 | 1.96 | 0 |
| A0A0K1U1X9 | Clade XV lectin receptor kinase | 3 | 8 | 1.266 | 1.145 | 4.89 | 7.08 |
| F8WQS4 | Quinone reductase (fragment) | 2 | 1 | 4.126 | 0.807 | 4.73 | 0 |
| A0A172WC56 | Defensin-like protein 1 | 2 | 1 | 3.97 | 0.732 | 2.84 | 0 |
| A2PYH3 | Nascent polypeptide associated complex α | 2 | 3 | 3.392 | 4.614 | 2.27 | 4.41 |
| Q6XX16 | Glutathione S-transferase U2 (fragment) | 2 | 3 | 2.565 | 1.677 | 3.44 | 0 |
| D6QX33 | Plastid RNA-binding protein | 2 | 1 | 2.326 | 0.695 | 0 | 0 |
| A0A0C5LA06 | Mitogen-activated protein kinase | 2 | 2 | 2.143 | 1.889 | 1.65 | 0 |
| F8WQS2 | Acetylglutamate kinase (fragment) | 1 | 2 | 3.648 | 2.075 | 3.27 | 0 |
| A0A0A7HDA5 | Epi-aristolochene dihydroxylase | 1 | 2 | 3.296 | 2.77 | 3.53 | 1.72 |
| Q18NX4 | Nitrate reductase | 1 | 1 | 2.896 | 1.947 | 2.65 | 0 |
| B0CN62 | Myosin VIII-1 | 1 | 1 | 2.364 | 1.331 | 2.55 | 0 |
| W6JJ90 | Nuclear pore complex protein Sec13d | 1 | 1 | 1.971 | 0.754 | 2.69 | 0 |
| Q20KN2 | Metacaspase type II (fragment) | 1 | 1 | 1.969 | 2.13 | 0 | 1.72 |
| Q5D1L7 | Serine/threonine protein kinase (fragment) | 1 | 1 | 1.888 | 1.185 | 2.11 | 0 |
| Q2QFR2 | Cysteine proteinase glycinain type (fragment) | 1 | 1 | 1.627 | 1.227 | 2.28 | 0 |
| C9DFC0 | Phytophthora-inhibited protease 1 (fragment) | 1 | 1 | 1.605 | 0.698 | 1.97 | 1.78 |
| A0A4Y5QRT8 | Serine/threonine protein kinase PBS1a | 1 | 2 | 1.396 | 1.303 | 0 | 1.77 |
| Q2QFR3 | Cysteine proteinase aleurain type | 1 | 1 | 1.191 | 1.367 | 0 | 1.62 |
| A0A024B875 | Dihydrolipoamide acetyltransferase component | 1 | 1 | 1.003 | 1.596 | 2.28 | 0 |
| D5JXY5 | Calcium-transporting ATPase | 1 | 1 | 0.754 | 0.754 | 0 | 0 |
| A0A1V1H6S6 | Calcium-dependent protein kinase isoform 2 | 1 | 1 | 0.732 | 0.801 | 2.11 | 0 |
| Q52JJ5 | Glutamyl-tRNA synthetase | 1 | 1 | 0.7 | 0.789 | 0 | 0 |
| A7L4B4 | Histone H3 | 1 | 2 | 0.509 | 1.589 | 1.76 | 2.54 |
| V5KY72 | Ubiquitin-conjugating enzyme variant | 1 | 1 | 0.503 | 0.766 | 0 | 0 |
| Description | Closest Homologs in | ||
|---|---|---|---|
| Tobacco | Tomato | Arabidopsis | |
| Carbonic anhydrase | LOC107768773 | Solyc02g086820 | AT3G01500 |
| Heat shock protein 90 (HSP90) | LOC107768797 | Solyc12g015880 | AT5G56000 |
| Adenosylhomocysteinase | LOC107783029 | Solyc09g092380 | AT4G13940 |
| Chloroplast photosystem II subunit O2 (PSBO2) | LOC107766588 | Solyc02g065400 | AT3G50820 |
| Chloroplast photosystem II subunit P1 (PSBP1) | LOC107830202 | Solyc07g044860 | AT1G06680 |
| MP-interacting protein (MIP) 1.2 | LOC107801992 | Solyc04g009770 | AT3G44110 |
| Heat shock protein 70 (HSP70) | LOC107803414 | Solyc11g066060 | AT3G12580 |
| Proteinase inhibitor | LOC107799889 | Solyc03g019690 | AT1G17860 |
| Ankyrin repeat containing protein 2 (AKR2) | LOC107793888 | Solyc01g104170 | AT2G17390 |
| Glucose-6-phosphate 1-dehydrogenase (G6PD) | LOC107794892 | Solyc07g045540 | AT5G35790 |
| Importin subunit α | LOC107810574 | Solyc01g060470 | AT4G16143 |
| RabG3 protein | LOC107815360 | Solyc03g120750 | AT1G52280 |
| NB-LRR HR-associated cell death (NRC) 2 | LOC107792680 | Solyc10g047320 | AT1G53350 |
| DNA gyrase subunit B | LOC107786139 | Solyc12g021230 | AT5G04130 |
| Calmodulin (CaM) | LOC107761764 | Solyc10g081170 | AT3G43810 |
| Adenosine kinase | LOC107790330 | Solyc09g007940 | AT5G03300 |
| Lipoxygenase | LOC107830099 | Solyc01g099160 | AT1G55020 |
| Ran binding protein RanBP | LOC107771336 | Solyc08g062660 | AT5G58590 |
| Lectin receptor kinase | LOC107782584 | Solyc03g080060 | AT5G55830 |
| Quinone reductase (fragment) | LOC107761412 | Solyc10g006650 | AT4G27270 |
| Defensin-like protein 1 | LOC107831752 | Solyc07g006380 | AT1G61070 |
| Nascent polypeptide associated complex α | LOC107791866 | Solyc10g081030 | AT3G12390 |
| Glutathione S-transferase U2 | LOC107782951 | Solyc07g056490 | AT1G78380 |
| Plastid RNA-binding protein | LOC107787150 | Solyc03g111050 | AT3G48500 |
| Mitogen-activated protein kinase (MAPK) | LOC107794128 | Solyc01g094960 | AT4G01370 |
| Acetylglutamate kinase | LOC107803486 | Solyc11g005620 | AT3G57560 |
| Epi-aristolochene dihydroxylase; CYP71B35 | LOC107759261 | Solyc04g083140 | AT3G26310 |
| Nitrate reductase | LOC107785409 | Solyc11g013810 | AT1G37130 |
| Myosin | LOC107806983 | Solyc02g020910 | AT3G19960 |
| Nuclear pore complex protein SEC13 | LOC107777830 | Solyc02g087300 | AT2G30050 |
| Metacaspase type II | LOC107824366 | Solyc09g098150 | AT1G79330 |
| Serine/threonine protein kinase (STPK) | LOC107808522 | Solyc02g067030 | AT3G01090 |
| Cysteine proteinase glycinain type | LOC107760226 | Solyc04g080960 | AT4G39090 |
| PIP1; cysteine endopeptidase | LOC107774651 | Solyc02g077040 | AT3G48340 |
| Serine/threonine protein kinase PBS1 | LOC107830934 | Solyc05g024290 | AT5G13160 |
| Cysteine proteinase aleurain type | LOC107784768 | Solyc07g041900 | AT5G60360 |
| Lipoamide acetyltransferase component | LOC107820956 | Solyc01g066520 | AT3G06850 |
| Calcium-transporting ATPase | LOC107814306 | Solyc04g016260 | AT3G57330 |
| Calcium-dependent protein kinase | LOC107805386 | Solyc07g064610 | AT3G20410 |
| Glutamyl-tRNA synthetase | LOC107774917 | Solyc01g112290 | AT5G64050 |
| Histone H3 | LOC107759185 | Solyc01g073970 | AT5G65360 |
| Ubiquitin-conjugating enzyme variant | LOC107831808 | Solyc04g007960 | AT1G70660 |
| GO Biological Process Complete | Arabidopsis Reference # | NSs-Interacting Proteins | ||||
|---|---|---|---|---|---|---|
| # | Expected | Fold Enrichment | +/− | p Value | ||
| Defense response to bacterium | 413 | 7 | 0.63 | 11.06 | + | 9.04 × 10−3 |
| Response to bacterium | 506 | 8 | 0.78 | 10.32 | + | 2.75 × 10−3 |
| Response to other organisms | 1092 | 12 | 1.67 | 7.17 | + | 1.74 × 10−4 |
| Interspecies interaction between organisms | 1120 | 12 | 1.72 | 6.99 | + | 2.28 × 10−4 |
| Response to external biotic stimulus | 1092 | 12 | 1.67 | 7.17 | + | 1.74 × 10−4 |
| Response to biotic stimulus | 1093 | 12 | 1.67 | 7.17 | + | 1.75 × 10−4 |
| Response to stimulus | 5567 | 22 | 8.53 | 2.58 | + | 1.20 × 10−2 |
| Response to external stimulus | 1509 | 15 | 2.31 | 6.49 | + | 9.27 × 10−6 |
| Defense response to other organisms | 805 | 9 | 1.23 | 7.30 | + | 8.95 × 10−3 |
| Defense response | 952 | 10 | 1.46 | 6.86 | + | 4.03 × 10−3 |
| Response to stress | 3091 | 18 | 4.74 | 3.80 | + | 6.09 × 10−4 |
| Cellular process | 11,979 | 33 | 18.35 | 1.80 | + | 1.67 × 10−2 |
| Unclassified | 5450 | 5 | 8.35 | 0.60 | − | 0.00 |
| GO Molecular Function Complete | Tobacco Reference # | NSs-Interacting Proteins | ||||
|---|---|---|---|---|---|---|
| # | Expected | Fold Enrichment | +/− | p Value | ||
| Unfolded protein binding | 284 | 4 | 0.16 | 24.64 | + | 4.15 × 10−2 |
| Binding | 21,517 | 26 | 12.30 | 2.11 | + | 5.53 × 10−3 |
| ATP binding | 4591 | 12 | 2.62 | 4.57 | + | 9.61 × 10−3 |
| Adenyl ribonucleotide binding | 4708 | 12 | 2.69 | 4.46 | + | 1.24 × 10−2 |
| Adenyl nucleotide binding | 4734 | 12 | 2.71 | 4.44 | + | 1.32 × 10−2 |
| Purine nucleotide binding | 5258 | 13 | 3.01 | 4.33 | + | 6.22 × 10−3 |
| Nucleotide binding | 5870 | 16 | 3.35 | 4.77 | + | 6.52 × 10−5 |
| Small molecule binding | 6512 | 17 | 3.72 | 4.57 | + | 3.66 × 10−5 |
| Nucleoside phosphate binding | 5870 | 16 | 3.35 | 4.77 | + | 6.52 × 10−5 |
| Purine ribonucleotide binding | 5217 | 13 | 2.98 | 4.36 | + | 5.70 × 10−3 |
| Ribonucleotide binding | 5285 | 14 | 3.02 | 4.63 | + | 9.62 × 10−4 |
| Carbohydrate derivative binding | 5332 | 14 | 3.05 | 4.59 | + | 1.07 × 10−3 |
| Purine ribonucleoside triphosphate binding | 5100 | 13 | 2.91 | 4.46 | + | 4.43 × 10−3 |
| Anion binding | 6438 | 16 | 3.68 | 4.35 | + | 2.38 × 10−4 |
| Ion binding | 11,853 | 20 | 6.77 | 2.95 | + | 1.59 × 10−3 |
| Unclassified | 26,668 | 2 | 15.24 | 0.13 | − | 0.00 |
| GO Molecular Function Complete | Arabidopsis Reference # | NSs-Interacting Proteins | ||||
| # | Expected | Fold Enrichment | +/− | p Value | ||
| Cysteine-type endopeptidase activity | 72 | 4 | 0.11 | 36.26 | + | 9.91 × 10−3 |
| Cysteine-type peptidase activity | 102 | 4 | 0.16 | 25.60 | + | 3.71 × 10−2 |
| Catalytic activity | 8305 | 27 | 12.72 | 2.12 | + | 1.06 × 10−2 |
| Cation binding | 1647 | 11 | 2.52 | 4.36 | + | 5.00 × 10−2 |
| Ion binding | 3071 | 16 | 4.70 | 3.40 | + | 1.04 × 10−2 |
| Binding | 9721 | 31 | 14.89 | 2.08 | + | 1.26 × 10−3 |
| Protein binding | 5109 | 23 | 7.83 | 2.94 | + | 3.25 × 10−4 |
| Unclassified | 5502 | 1 | 8.43 | 0.12 | − | 0.00 |
| GO Cellular Component Complete | Tomato Reference # | NSs-Interacting Proteins | ||||
|---|---|---|---|---|---|---|
| # | Expected | Fold Enrichment | +/− | p Value | ||
| Lysosome | 49 | 3 | 0.06 | 50.49 | + | 1.71 × 10−2 |
| Lytic vacuole | 52 | 3 | 0.06 | 47.58 | + | 2.02 × 10−2 |
| Intracellular membrane-bounded organelle | 5532 | 18 | 6.71 | 2.68 | + | 1.61 × 10−2 |
| Membrane-bounded organelle | 5782 | 19 | 7.01 | 2.71 | + | 7.25 × 10−3 |
| Organelle | 6262 | 19 | 7.59 | 2.50 | + | 2.30 × 10−2 |
| Cellular anatomical entity | 9174 | 26 | 11.12 | 2.34 | + | 7.57 × 10−4 |
| Intracellular organelle | 6130 | 19 | 7.43 | 2.56 | + | 1.70 × 10−2 |
| Intracellular | 7723 | 25 | 9.36 | 2.67 | + | 1.09 × 10−4 |
| Cytoplasm | 5053 | 21 | 6.13 | 3.43 | + | 3.26 × 10−5 |
| Extracellular space | 62 | 3 | 0.08 | 39.90 | + | 3.34 × 10−2 |
| Unclassified | 25,226 | 16 | 30.59 | 0.52 | − | 0.00 |
| GO Cellular Component Complete | Arabidopsis Reference # | NSs-Interacting Proteins | ||||
| # | Expected | Fold Enrichment | +/− | p Value | ||
| Lysosome | 46 | 3 | 0.07 | 42.57 | + | 3.98 × 10−2 |
| Vacuole | 1084 | 10 | 1.66 | 6.02 | + | 3.03 × 10−3 |
| Cytoplasm | 14,776 | 38 | 22.64 | 1.68 | + | 3.25 × 10−4 |
| Chloroplast stroma | 718 | 8 | 1.10 | 7.27 | + | 8.34 × 10−3 |
| Plastid stroma | 730 | 8 | 1.12 | 7.15 | + | 9.39 × 10−3 |
| Whole membrane | 830 | 8 | 1.27 | 6.29 | + | 2.33 × 10−2 |
| Membrane | 5495 | 22 | 8.42 | 2.61 | + | 2.28 × 10−3 |
| Bounding membrane of organelle | 921 | 8 | 1.41 | 5.67 | + | 4.82 × 10−2 |
| Cytosol | 3242 | 22 | 4.97 | 4.43 | + | 1.35 × 10−7 |
| Plasma membrane | 3529 | 18 | 5.41 | 3.33 | + | 1.05 × 10−3 |
| Cell periphery | 4001 | 19 | 6.13 | 3.10 | + | 1.35 × 10−3 |
| Unclassified | 1877 | 1 | 2.88 | 0.35 | − | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, Y.; Gnanasekaran, P.; Pappu, H.R. Identification and Characterization of Plant-Interacting Targets of Tomato Spotted Wilt Virus Silencing Suppressor. Pathogens 2021, 10, 27. https://doi.org/10.3390/pathogens10010027
Zhai Y, Gnanasekaran P, Pappu HR. Identification and Characterization of Plant-Interacting Targets of Tomato Spotted Wilt Virus Silencing Suppressor. Pathogens. 2021; 10(1):27. https://doi.org/10.3390/pathogens10010027
Chicago/Turabian StyleZhai, Ying, Prabu Gnanasekaran, and Hanu R. Pappu. 2021. "Identification and Characterization of Plant-Interacting Targets of Tomato Spotted Wilt Virus Silencing Suppressor" Pathogens 10, no. 1: 27. https://doi.org/10.3390/pathogens10010027