Co-Infection with Anaplasma Species and Novel Genetic Variants Detected in Cattle and Goats in the Republic of Korea

 ,
,  and
and
Abstract
:
1. Introduction
2. Results
2.1. Distribution of Anaplasma Infection
2.2. Anaplasma Species Prevalence: Single Infection and Co-Infection Cases
2.3. Total Number of Animals Infected per Anaplasma Species Analyzed
2.4. PCR and Molecular Identification
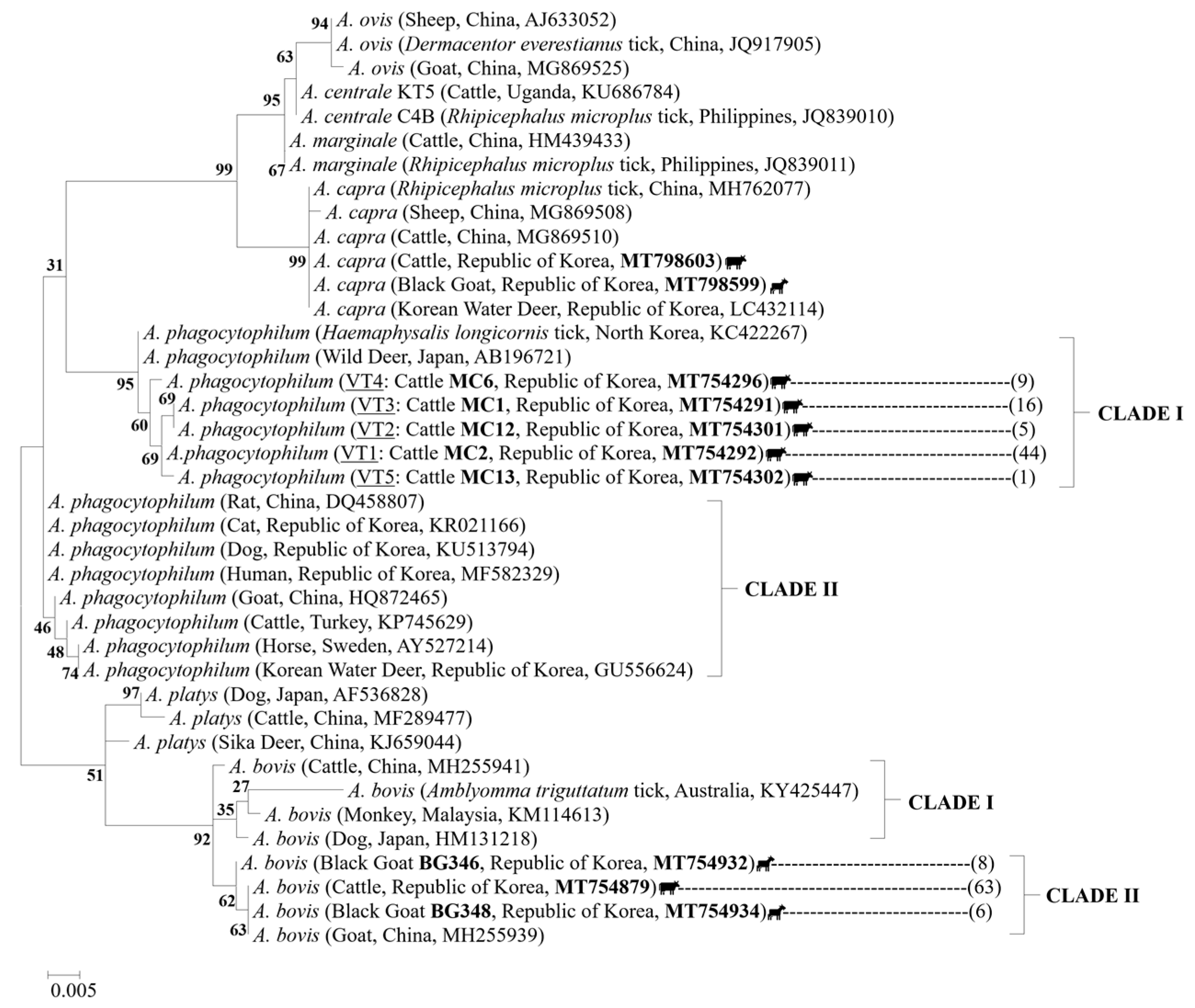
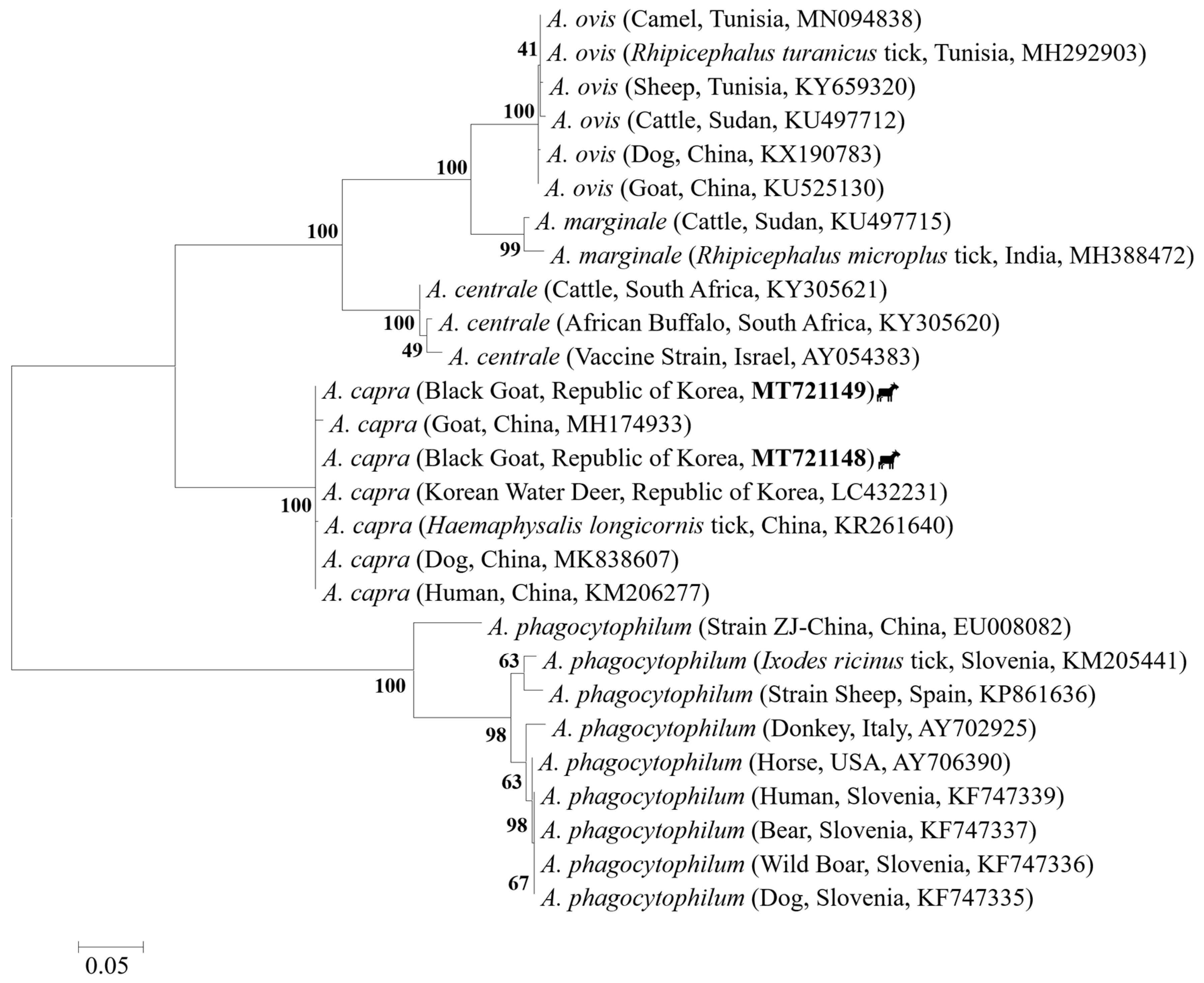
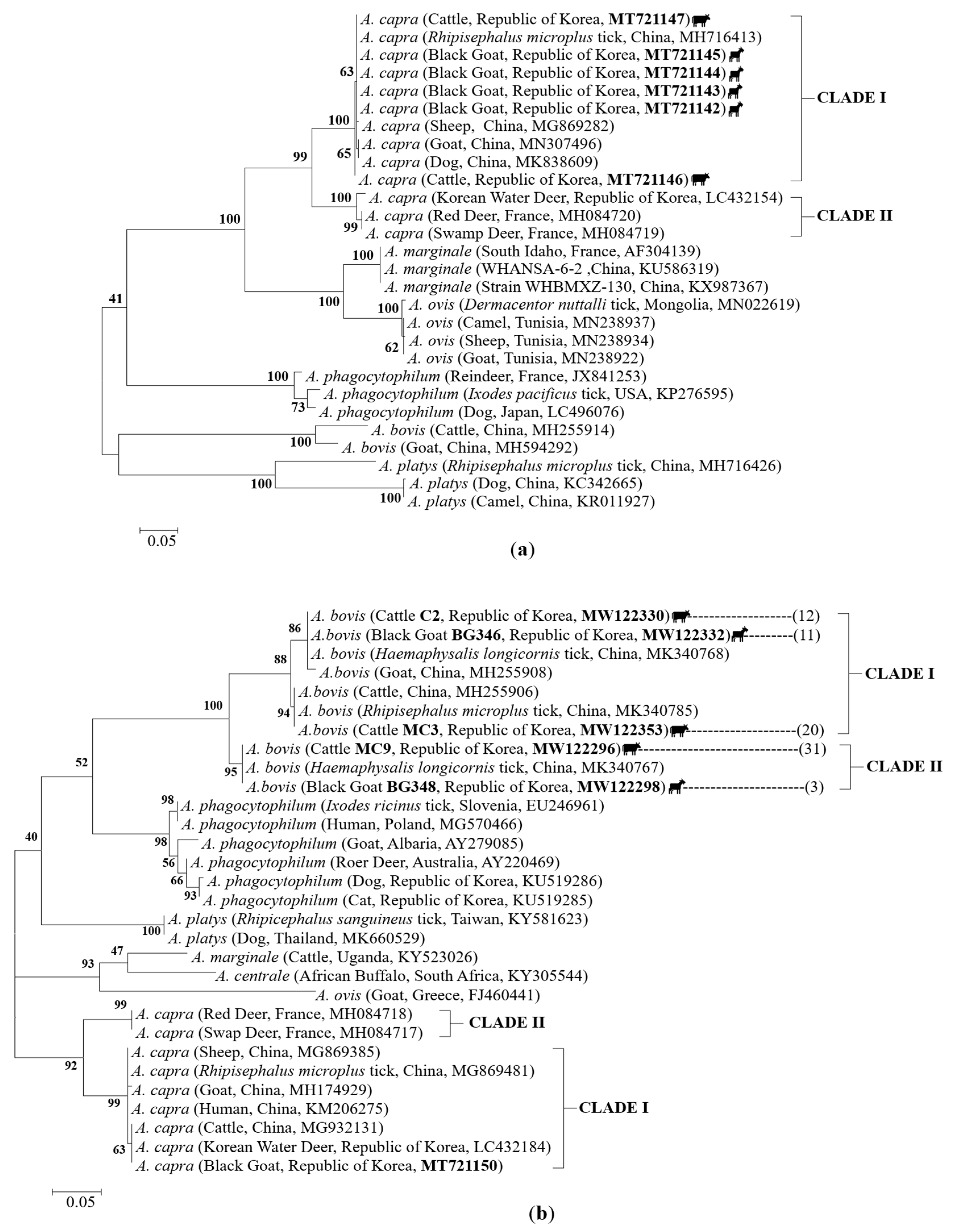
2.5. Sequences and Phylogenetic Analysis
3. Discussion
4. Material and Methods
4.1. Sample Collection
4.2. DNA Extraction
4.3. PCR Amplification
4.4. DNA Sequencing and Phylogenetic Analysis
4.5. Nucleotide Sequence Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aubry, P.; Geale, D.W. A review of Bovine anaplasmosis. Transbound. Emerg. Dis. 2011, 58, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Theiler, A. Anaplasma marginale (Gen. and Spec. Nov.): The Marginal Points in the Blood of Cattle Suffering from a Specific Disease; Report of the Government Veterinary Bacteriologist of the Transvaal; Transvaal Department of Agriculture: Pretoria, South Africa, 1910; pp. 7–64.
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 104–122. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, C.; Anandan, R.; Balasundaram, S.; Rajavelu, G. Morphology and staining characteristics of Ehrlichia bovis. Comp. Immunol. Microbiol. Infect. Dis. 1996, 1, 79–83. [Google Scholar] [CrossRef]
- Uilenberg, G.; Van Vorstenbosch, C.J.A.H.V.; Perié, N.M. Blood parasites of sheep in the netherlands. I. Anaplasma mesaeterum sp.n. (Rickettsiales, Anaplasmataceae). Vet. Q. 1979, 1, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Munderloh, U.G.; Lynch, M.J.; Herron, M.J.; Palmer, A.T.; Kurtti, T.J.; Nelson, R.D.; Goodman, J.L. Infection of endothelial cells with Anaplasma marginale and A. phagocytophilum. Vet. Microbiol. 2004, 101, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G.; Jiang, R.R.; Huo, Q.B.; Wang, Y.W.; Liu, H.B.; Chu, Y.L.; et al. Human infection with a novel tick-borne Anaplasma species in China: A surveillance study. Lancet Infect. Dis. 2015, 15, 663–670. [Google Scholar] [CrossRef]
- de la Fuente, J.; Lew, A.; Lutz, H.; Meli, M.L.; Hofmann-Lehmann, R.; Shkap, V.; Molad, T.; Mangold, A.J.; Almazán, C.; Naranjo, V.; et al. Genetic diversity of Anaplasma species major surface proteins and implications for anaplasmosis serodiagnosis and vaccine development. Anim. Health Res. Rev. 2005, 6, 75–89. [Google Scholar] [CrossRef] [Green Version]
- Underwood, W.J.; Blauwiekel, R.; Delano, M.L.; Gillesby, R.; Mischler, S.A.; Schoell, A. Chapter 15—Biology and Diseases of Ruminants (Sheep, Goats, and Cattle). In Laboratory Animal Medicine, 3rd ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 623–694. [Google Scholar] [CrossRef]
- Ybañez, A.P.; Inokuma, H. Anaplasma species of veterinary importance in Japan. Vet. World 2016, 11, 1190–1196. [Google Scholar] [CrossRef] [Green Version]
- Han, D.G.; Ryu, J.H.; Chae, J.B.; Kim, D.W.; Kwon, C.H.; Choi, K.S. First report of Anaplasma phagocytophilum infection in Holstein cattle in the Republic of Korea. Acta Trop. 2018, 183, 110–113. [Google Scholar] [CrossRef]
- Park, J.; Han, D.G.; Ryu, J.H.; Chae, J.B.; Chae, J.S.; Yu, D.H.; Park, B.K.; Kim, H.C.; Choi, K.S. Molecular detection of Anaplasma bovis in Holstein cattle in the Republic of Korea. Acta Vet. Scand. 2018, 60, 1–5. [Google Scholar] [CrossRef]
- Seo, M.G.; Ouh, I.O.; Lee, H.; Geraldino, P.J.L.; Rhee, M.H.; Kwon, O.D.; Kwak, D. Differential identification of Anaplasma in cattle and potential of cattle to serve as reservoirs of Anaplasma capra, an emerging tick-borne zoonotic pathogen. Vet. Microbiol. 2018, 226, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Carelli, G.; Decaro, N.; Lorusso, E.; Paradies, P.; Elia, G.; Martella, V.; Buonavoglia, C.; Ceci, L. First report of bovine anaplasmosis caused by Anaplasma centrale in Europe. Proc. Ann. N. Y. Acad. Sci. 2008, 1149, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, M.; Wang, Z.; Wang, J.; Peng, Y.; Li, Y.; Guan, G.; Luo, J.; Yin, H. Molecular survey and genetic identification of Anaplasma species in goats from central and southern China. Appl. Environ. Microbiol. 2012, 78, 464–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.J.; Jin, B.C.; Kim, K.H.; Yoo, M.S.; Seong, K.W.; Jeong, S.J.; Hyun, B.H.; Cho, Y.S. Molecular detection and phylogenetic analysis of Anaplasma spp. in Korean native goats from Ulsan Metropolitan City, Korea. Vector-Borne Zoonotic Dis. 2019, 19, 773–776. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Liu, Z.; Liu, J.; Niu, Q.; Ren, Q.; Chen, Z.; Guan, G.; Luo, J.; Yin, H. Molecular detection and characterization of Anaplasma spp. in sheep and cattle from Xinjiang, northwest China. Parasites Vectors 2015, 8, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.G.; Ouh, I.O.; Kwon, O.D.; Kwak, D. Molecular detection of Anaplasma phagocytophilum-like Anaplasma spp. and pathogenic A. Phagocytophilum in cattle from South Korea. Mol. Phylogenet. Evol. 2018, 126, 23–30. [Google Scholar] [CrossRef]
- Hoogstraal, H. Ticks in relation to human diseases caused by Rickettsia species. Annu. Rev. Entomol. 1967, 12, 377–420. [Google Scholar] [CrossRef]
- Kim, K.H.; Yi, J.; Oh, W.S.; Kim, N.H.; Choi, S.J.; Choe, P.G.; Kim, N.J.; Lee, J.K.; Oh, M.D. Human granulocytic anaplasmosis, South Korea, 2013. Emerg. Infect. Dis. 2014, 20, 1708. [Google Scholar] [CrossRef]
- Chochlakis, D.; Ioannou, I.; Tselentis, Y.; Psaroulaki, A. Human anaplasmosis and Anaplasma ovis variant. Emerg. Infect. Dis. 2010, 16, 1031–1032. [Google Scholar] [CrossRef]
- Heo, E.J.; Park, J.H.; Koo, J.R.; Park, M.S.; Park, M.Y.; Dumler, J.S.; Chae, J.S. Serologic and molecular detection of Ehrlichia chaffeensis and Anaplasma phagocytophila (human granulocytic ehrlichiosis agent) in Korean patients. J. Clin. Microbiol. 2002, 40, 3082–3085. [Google Scholar] [CrossRef] [Green Version]
- Battilani, M.; De Arcangeli, S.; Balboni, A.; Dondi, F. Genetic diversity and molecular epidemiology of Anaplasma. Infect. Genet. Evol. 2017, 49, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Ko, S.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Chae, J.B.; Jo, Y.S.; Choi, K.S.; Yu, D.H.; Park, B.K.; et al. Prevalence of Anaplasma and Bartonella spp. In ticks collected from Korean water deer (Hydropotes inermis argyropus). Korean J. Parasitol. 2016, 54, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Molecular and phylogenetic analysis of tick-borne pathogens in ticks parasitizing native korean goats (Capra hircus coreanae) in South Korea. Pathogens 2020, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.W.; Doan, H.T.T.; Choe, S.E.; Noh, J.H.; Yoo, M.S.; Reddy, K.E.; Kim, Y.H.; Kweon, C.H.; Jung, S.C.; Chang, K.Y. Molecular investigation of tick-borne pathogens in ticks from grazing cattle in Korea. Parasitol. Int. 2013, 62, 276–282. [Google Scholar] [CrossRef]
- Oh, J.Y.; Moon, B.C.; Bae, B.K.; Shin, E.H.; Ko, Y.H.; Kim, Y.J.; Park, Y.H.; Chae, J.S. Genetic identification and phylogenetic analysis of Anaplasma and Ehrlichia species in Haemaphysalis longicornis collected from Jeju island, Korea. J. Bacteriol. Virol. 2009, 39, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Durden, L.A. Taxonomy, host associations, life cycles and vectorial importance of ticks parasitizing small mammals. In Micromammals and Macroparasites: From Evolutionary Ecology to Management; Springer: Tokyo, Japan, 2006; pp. 91–102. [Google Scholar] [CrossRef]
- Baneth, G. Tick-borne infections of animals and humans: A common ground. Int. J. Parasitol. 2014, 44, 591–596. [Google Scholar] [CrossRef]
- Lee, S.H.; Jung, B.Y.; Kwak, D. Evidence of Anaplasma spp. exposure in native Korean goats (Capra hircus coreanae). Vet. Med. 2015, 60, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Barlough, J.E.; Madigan, J.E.; DeRock, E.; Bigornia, L. Nested polymerase chain reaction for detection of Ehrlichia equi genomic DNA in horses and ticks (Ixodes pacificus). Vet. Parasitol. 1996, 63, 319–329. [Google Scholar] [CrossRef]
- Kang, J.G.; Ko, S.; Kim, Y.J.; Yang, H.J.; Lee, H.; Shin, N.S.; Choi, K.S.; Chae, J.S. New genetic variants of Anaplasma phagocytophilum and Anaplasma bovis from Korean water deer (Hydropotes inermis argyropus). Vector-Borne Zoonotic Dis. 2011, 11, 929–938. [Google Scholar] [CrossRef]
- Lin, Q.; Rikihisa, Y.; Felek, S.; Wang, X.; Massung, R.F.; Woldehiwet, Z. Anaplasma phagocytophilum has a functional msp2 gene that is distinct from p44. Infect. Immun. 2004, 3883–3889. [Google Scholar] [CrossRef] [Green Version]
- Massung, R.F.; Owens, J.H.; Ross, D.; Reed, K.D.; Petrovec, M.; Bjoersdorff, A.; Coughlin, R.T.; Beltz, G.A.; Murphy, C.I. Sequence analysis of the ank gene of granulocytic ehrlichiae. J. Clin. Microbiol. 2000, 36, 1090–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.P.; Wang, X.; Li, Y.N.; Xu, G.; Wang, Y.H.; Zhou, E.M. GroEL gene typing and genetic diversity of Anaplasma bovis in ticks in Shaanxi, China. Infect. Genet. Evol. 2019, 74, 103927. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Han, R.; Liu, G.; Shi, Y.; Luo, J.; Yin, H. Molecular survey and characterization of a novel Anaplasma species closely related to Anaplasma capra in ticks, northwestern China. Parasites Vectors 2016, 9, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Han, R.; Guan, G.; Hassan, M.A.; Liu, G.; Luo, J.; Yin, H. A novel zoonotic Anaplasma species is prevalent in small ruminants: Potential public health implications. Parasites Vectors 2017, 10, 264. [Google Scholar] [CrossRef]
- de la Fuente, J.; Atkinson, M.W.; Naranjo, V.; Fernández de Mera, I.G.; Mangold, A.J.; Keating, K.A.; Kocan, K.M. Sequence analysis of the msp4 gene of Anaplasma ovis strains. Vet. Microbiol. 2007, 119, 375–381. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Species | No. Tested | No. of Positive (IR 1, %) | Type of Infection Identified in Tested Blood Samples | Not Infected (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Single Infection | Double Infection | Triple Infection | ||||||||
| A. phago2 (%) | A. bovis (%) | A. capra (%) | A. ovis (%) | A. bovis + A. phago 2 (%) | A. bovis + A. capra (%) | A. phago2 + A. bovis + A. capra (%) | ||||
| Cattle | 384 | 87 (22.7) | 16 (4.2) | 5 (1.3) | 1 (0.3) | 0 (0) | 64 (16.7) | 0 (0) | 1 (0.3) | 297 (77.3) |
| Goat | 302 | 23 (8.0) | 0 (0) | 19 (6.3) | 1 (0.3) | 0 (0) | 0 (0) | 3 (1.0) | 0 (0) | 279 (94.4) |
| Total | 686 | 110 (16.0) | 16 (2.3) | 24 (3.5) | 2 (0.3) | 0 (0) | 64 (9.3) | 3 (0.4) | 1 (0.1) | 576 (83.9) |
| Host | Collected Province | No. Tested | A. phago1 (%) | A. bovis (%) | A. capra (%) | A. ovis (%) |
|---|---|---|---|---|---|---|
| Cattle | Gyeongsangbuk-do | 90 | 80 (88.9) | 67 (74.4) | 1 (1.1) | 0 (0) |
| Gyeongsangnam-do | 65 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Jeollanam-do | 65 | 0 (0) | 1 (1.5) | 0 (0) | 0 (0) | |
| Gyonggi-do | 50 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Chungbuk-do | 54 | 1 (1.9) | 1 (1.9) | 1 (1.9) | 0 (0) | |
| Chungnam-do | 60 | 0 (0) | 1 (1.7) | 0 (0) | 0 (0) | |
| Subtotal | 384 | 81 (21.1) | 70 (18.2) | 2 (0.5) | 0 (0) | |
| Goat | Jeollabuk-do | 37 | 0 (0) | 10 (27.0) | 2 (5.4) | 0 (0) |
| Gwangju Metropolitan City | 41 | 0 (0) | 3 (7.3) | 0 (0) | 0 (0) | |
| Jeollanam-do | 224 | 0 (0) | 9 (4.0) | 2 (1.0) | 0 (0) | |
| Subtotal | 302 | 0 (0) | 22 (7.3) | 4 (1.3) | 0 (0) | |
| Grand total | 686 | 81 (11.8) | 92 (13.4) | 6 (0.9) | 0 (0) | |
| Species | Target Gene | Primer Name and PCR Conditions | Primer Sequences (5′-3′) | Cycles | Amplicon Size (bp) | References | ||
|---|---|---|---|---|---|---|---|---|
| Denaturation (°C/min) | Annealing (°C/min) | Extension (°C/min) | ||||||
| Anaplasma spp. | 16S rRNA1 | AE1-F | AAGCTTAACACATGCAAGTCGAA | 35 | 1406 | Oh et al. (2009) [27] | ||
| AE1-R | AGTCACTGACCCAACCTTAAATG | |||||||
| Conditions | 94/1 | 56/1 | 72/1.5 | |||||
| A. phagocytophilum | 16S rRNA2 | EE3 | GTCGAACGGATTATTCTTTATAGCTTGC | 25 | 926 | Barlough et al. (1996) [31] | ||
| EE4 | CCCTTCCGTTAAGAAGGATCTAATCTCC | |||||||
| Conditions | 94/0.50 | 56/0.50 | 72/0.75 | |||||
| msp2 | msp2fullF | TCAGAAAGATACACGTGCGCCC | 35 | 1079 | Lin et al. (2004) [33] | |||
| msp2fullR | TTATGATTAGGCCTTTGGGCATG | |||||||
| Conditions | 94/1 | 54/1 | 72/1 | |||||
| msp2F | GGTTACATAAGGGCCGCAAAGGTG | 25 | 467 | |||||
| msp2R | CCGGCGCATGTGTAAGGTGAAA | |||||||
| Conditions | 94/0.5 | 57/0.5 | 72/0.5 | |||||
| ankA | U7 | GCGTCTGTAAGGCAGATTGTG | 35 | 1696 | Massung et al. (2000) [34] | |||
| 1R1 | TATACACCTGGAGTAGGAAC | |||||||
| Conditions | 94/1 | 57/1 | 72/1.5 | |||||
| U8 | TAAGATAGGTTTAGTAAGACG | 25 | 460 | |||||
| 1R7 | TGCATCGTCATTACGCACAAGGTC | |||||||
| Conditions | 94/0.75 | 57/0.75 | 72/0.75 | |||||
| A. bovis | 16S rRNA3 | ABKf | TAGCTTGCTATGGGGACAA | 25 | 547 | Kang et al. (2011) [32] | ||
| AB1r | TCTCCCGGACTCCAGTCTG | |||||||
| Conditions | 94/0.5 | 59/0.5 | 72/0.5 | |||||
| groEL | bovis-groEL-F1 | GTTCGCAGTATTTTGCCAGT | 30 | 845 | Guo et al. (2019) [35] | |||
| bovisgroEL-R | CTGCRTTCAGAGTCATAAATAC | |||||||
| bovis-groEL-F2 | ATCTGGAAGRCCACTATTGAT | |||||||
| Conditions | 94/0.7 | 56/0.7 | 72/1 | |||||
| A. capra | 16S rRNA | Forward | TTGAGAGTTTGATCCTGGCTCAGAACG | 57 | 1499 | Li et al. (2015) [7] | ||
| Reverse | WAAGGWGGTAATCCAGC | |||||||
| Conditions | 94/0.75 | 57/0.75 | 72/0.75 | |||||
| gltA | Outer-f | GCGATTTTAGAGTGYGGAGATTG | 30 | 1031 | ||||
| Outer-r | TACAATACCGGAGTAAAAGTCAA | |||||||
| Conditions | 94/0.75 | 55/0.75 | 72/0.75 | |||||
| Inner-f | TCATCTCCTGTTGCACGGTGCCC | 30 | 594 | Yang et al. (2016) [36] | ||||
| Inner-r | CTCTGAATGAACATGCCCACCCT | |||||||
| Conditions | 94/0.75 | 60/0.75 | 72/0.75 | |||||
| groEL | Forward | TGAAGAGCATCAAACCCGAAG | 30 | 874 | Yang et al. (2017) [37] | |||
| Reverse | CTGCTCGTGATGCTATCGG | |||||||
| Conditions | 94/0.75 | 55/0.75 | 72.0.75 | |||||
| msp4 | Forward | GGGTTCTGATATGGCATCTTC | 30 | 656 | ||||
| Reverse | GGGAAATGTCCTTATAGGATTCG | |||||||
| Conditions | 94/0.75 | 53/0.75 | 72/0.75 | |||||
| A. ovis | msp4 | MSP45 | GGGAGCTCCTATGAATTACAGAGAATTGTTTAC | 35 | 852 | De la Fuente et al. (2007) [38] | ||
| MSP43 | CCGGATCCTTAGCTGAACAGGAATCTTGC | |||||||
| Conditions | 94/0.5 | 60/0.5 | 68/1 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, E.A.; Han, S.-W.; Cho, Y.-K.; Choi, K.-S.; Chae, J.-S. Co-Infection with Anaplasma Species and Novel Genetic Variants Detected in Cattle and Goats in the Republic of Korea. Pathogens 2021, 10, 28. https://doi.org/10.3390/pathogens10010028
Miranda EA, Han S-W, Cho Y-K, Choi K-S, Chae J-S. Co-Infection with Anaplasma Species and Novel Genetic Variants Detected in Cattle and Goats in the Republic of Korea. Pathogens. 2021; 10(1):28. https://doi.org/10.3390/pathogens10010028
Chicago/Turabian StyleMiranda, Evelyn Alejandra, Sun-Woo Han, Yoon-Kyong Cho, Kyoung-Seong Choi, and Joon-Seok Chae. 2021. "Co-Infection with Anaplasma Species and Novel Genetic Variants Detected in Cattle and Goats in the Republic of Korea" Pathogens 10, no. 1: 28. https://doi.org/10.3390/pathogens10010028