
Molecular Profiling of Athletes Performing High-Intensity Exercises in Extreme Environments

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Stress Testing
2.3. Plasma Samples
2.4. Mass Spectrometric Analysis
2.5. ELISA
2.6. Statistical Analysis
3. Results
3.1. Analysis of the Metabolomic Profile of Participants before and after Exercise
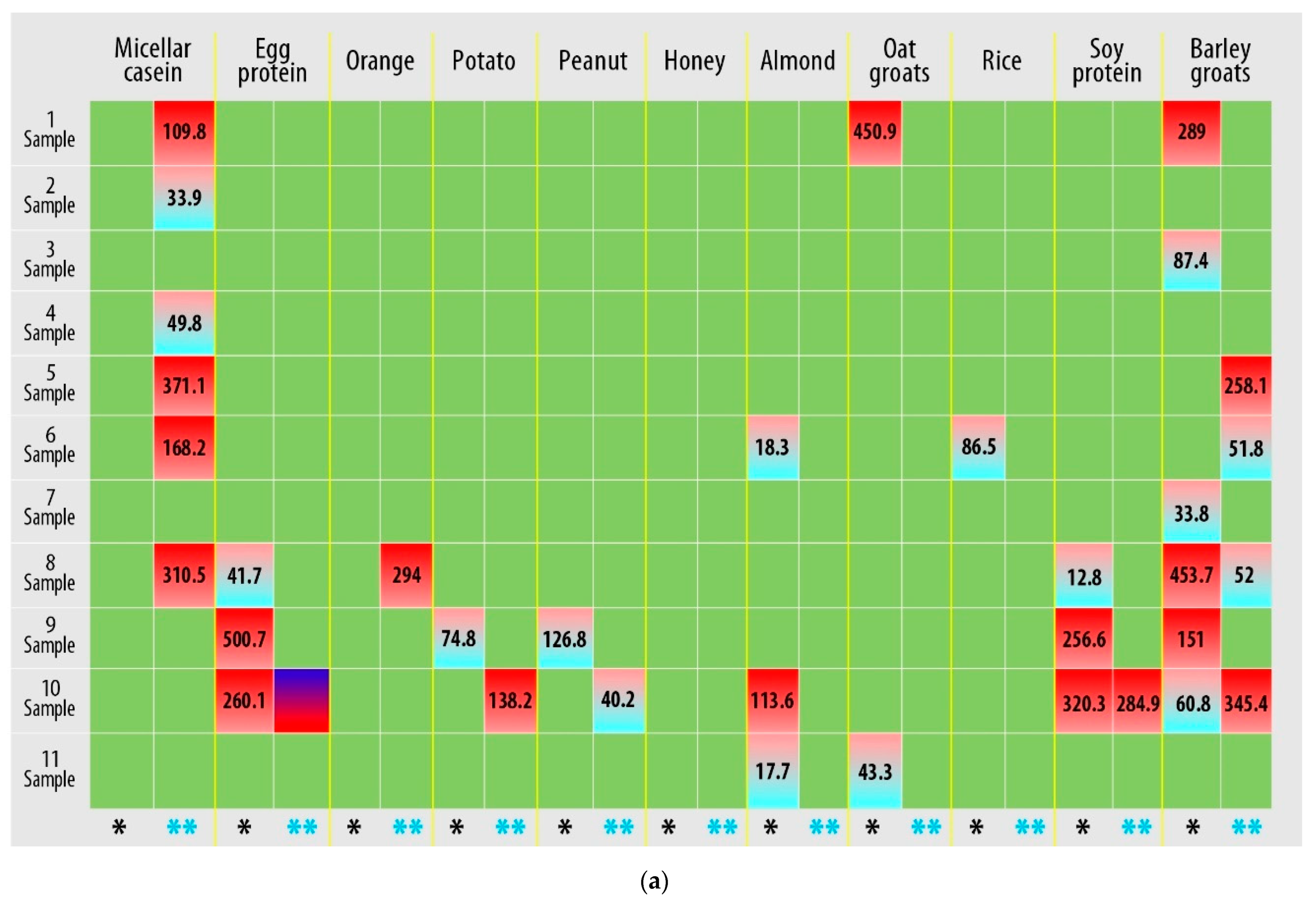
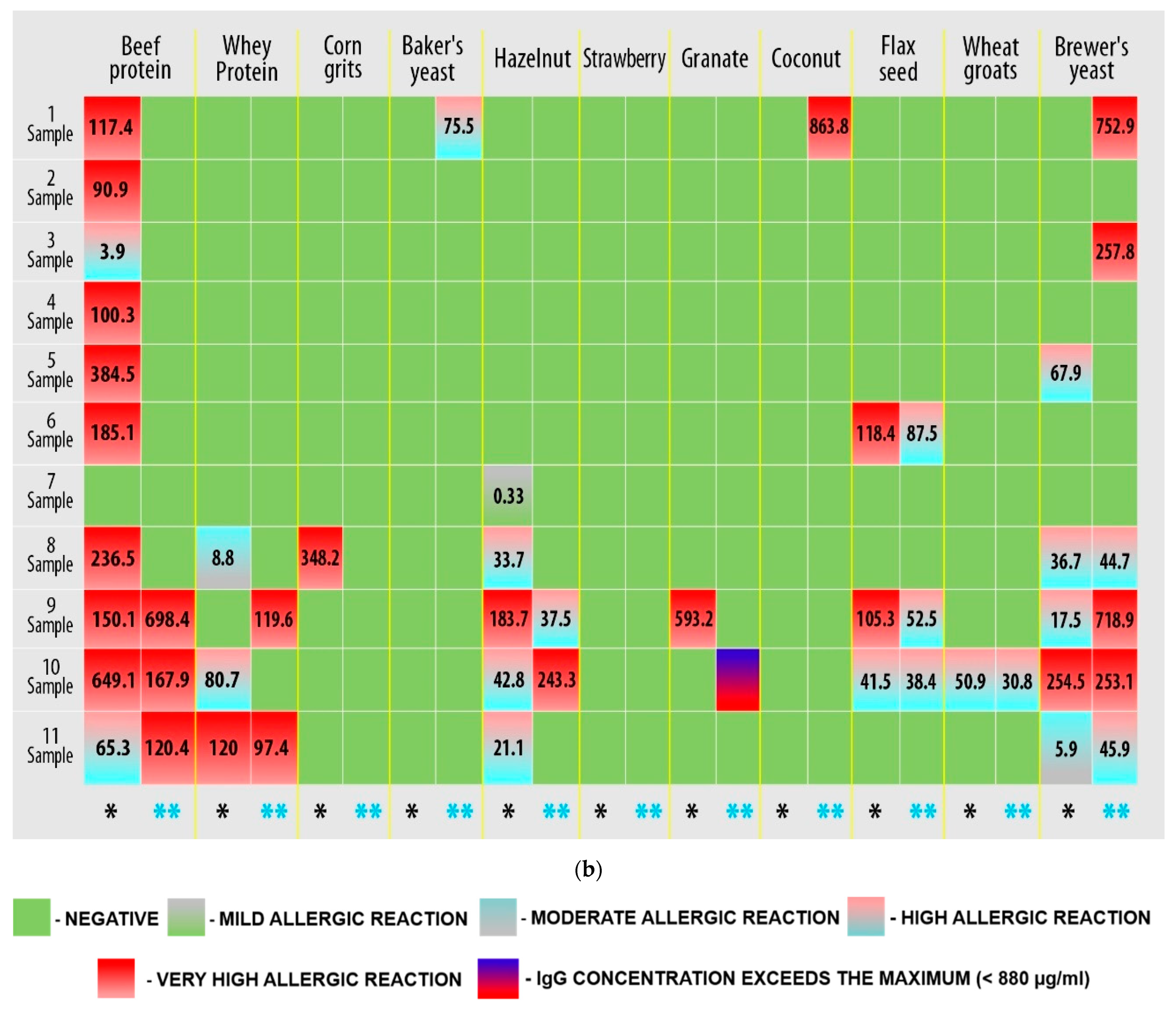
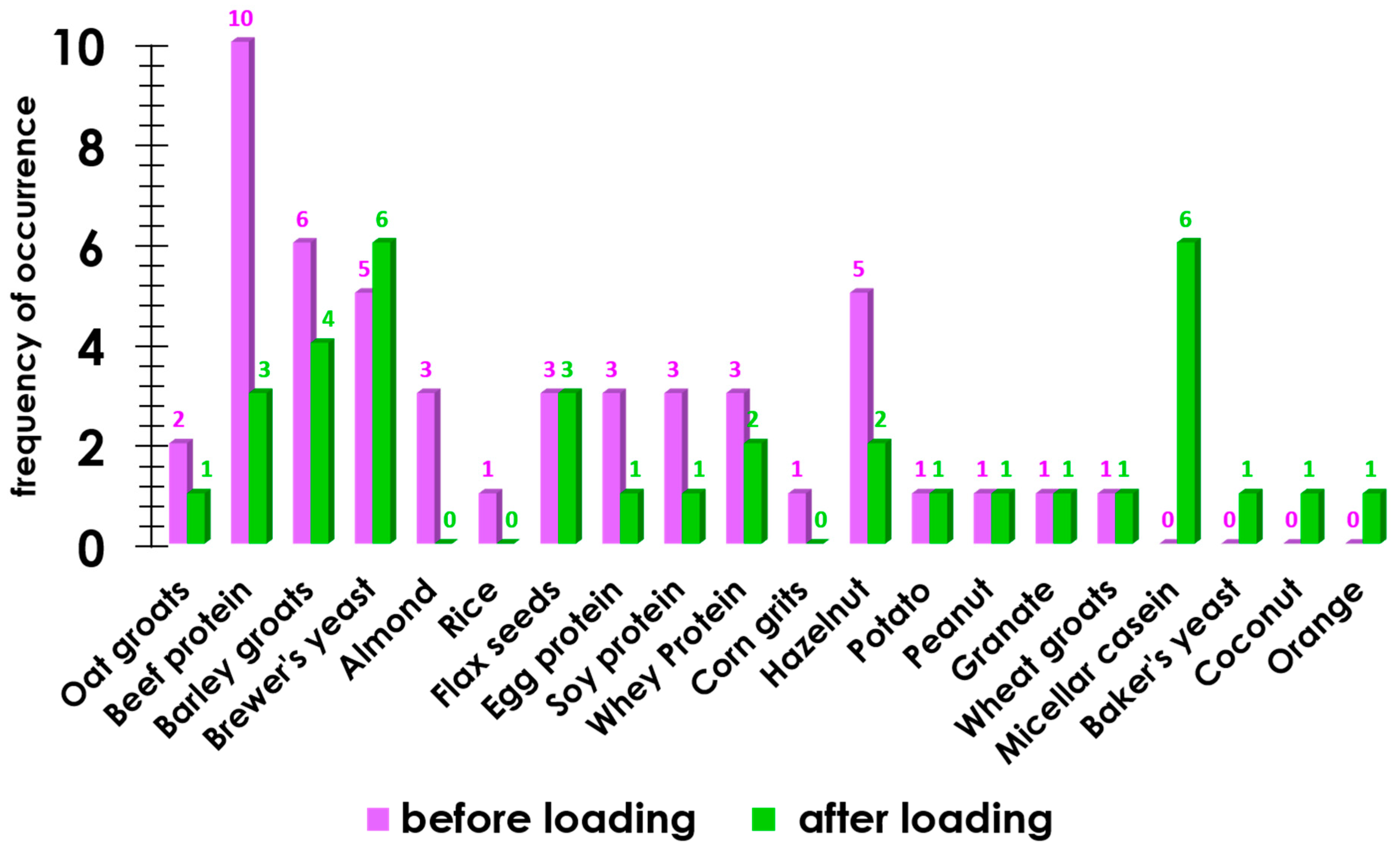
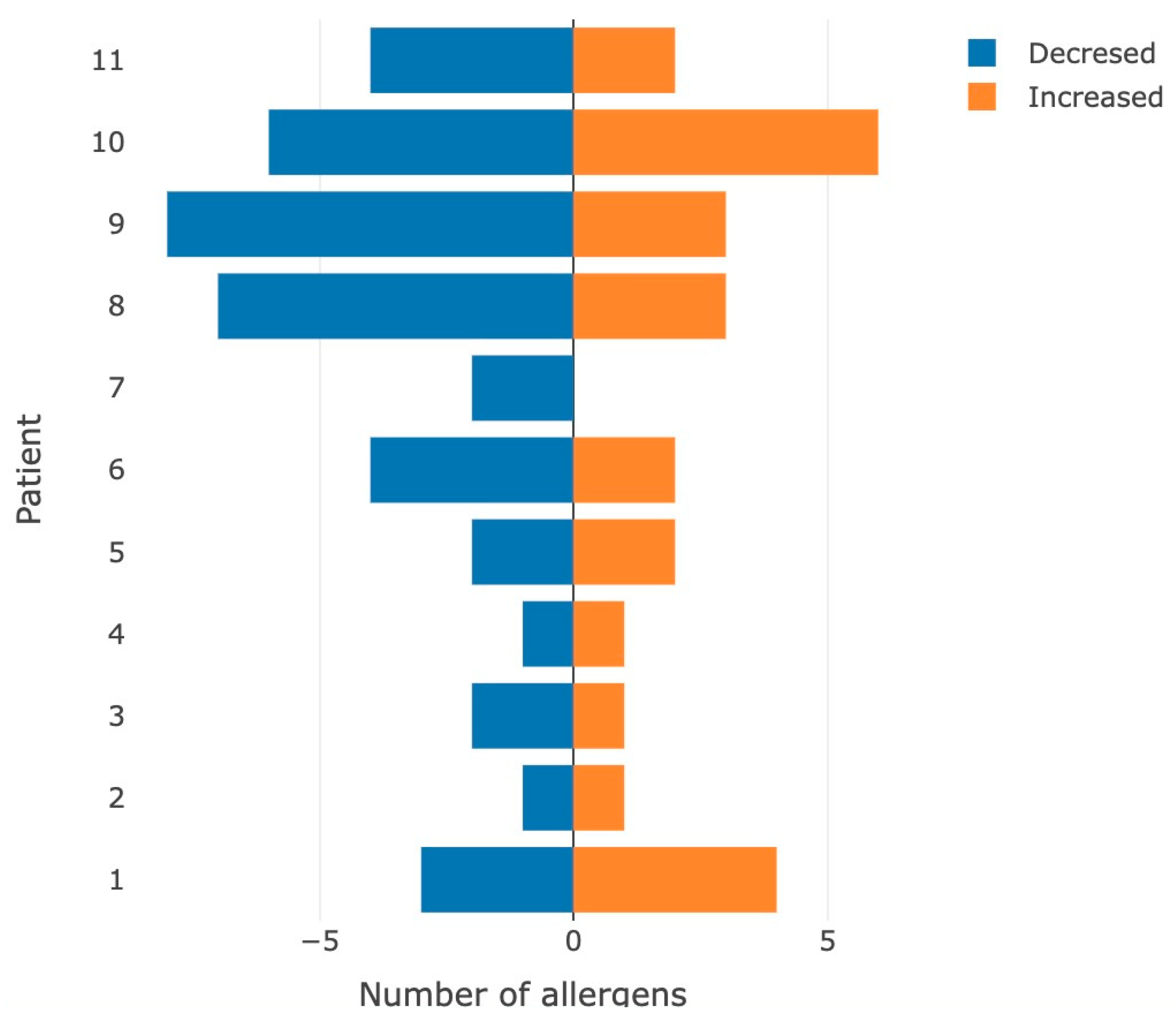
3.2. Hyper-Reactive Reaction to Food Allergens Mediated by Immune Processes
4. Discussion
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peake, J.M.; Tan, S.J.; Markworth, J.F.; Broadbent, J.A.; Skinner, T.L.; Cameron-Smith, D. Metabolic and Hormonal Responses to Isoenergetic High-Intensity Interval Exercise and Continuous Moderate-Intensity Exercise. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E539–E552. [Google Scholar] [CrossRef] [PubMed]
- Gillen, J.B.; Gibala, M.J. Is High-Intensity Interval Training a Time-Efficient Exercise Strategy to Improve Health and Fitness? Appl. Physiol. Nutr. Metab. 2014, 39, 409–412. [Google Scholar] [CrossRef]
- Lewis, G.D.; Farrell, L.; Wood, M.J.; Martinovic, M.; Arany, Z.; Rowe, G.C.; Souza, A.; Cheng, S.; McCabe, E.L.; Yang, E.; et al. Metabolic Signatures of Exercise in Human Plasma. Sci. Transl. Med. 2010, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Lac, G.; Maso, F. Biological Markers for the Follow-up of Athletes throughout the Training Season. Pathol. Biol. 2004, 52, 43–49. [Google Scholar] [CrossRef]
- Yan, B.; Jiye, A.; Wang, G.; Lu, H.; Huang, X.; Liu, Y.; Zha, W.; Hao, H.; Zhang, Y.; Liu, L.; et al. Metabolomic Investigation into Variation of Endogenous Metabolites in Professional Athletes Subject to Strength-Endurance Training. J. Appl. Physiol. 2009, 106, 531–538. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Connelly, J.; Lindon, J.C.; Holmes, E. Metabonomics: A Platform for Studying Drug Toxicity and Gene Function. Nat. Rev. Drug Discov. 2002, 1, 153–161. [Google Scholar] [CrossRef]
- Heaney, L.M.; Deighton, K.; Suzuki, T. Non-Targeted Metabolomics in Sport and Exercise Science. J. Sport. Sci. 2019, 37, 959–967. [Google Scholar] [CrossRef]
- Goodwin, M.L.; Harris, J.E.; Hernández, A.; Gladden, L.B. Blood Lactate Measurements and Analysis during Exercise: A Guide for Clinicians. J. Diabetes. Sci. Technol. 2007, 1, 558–569. [Google Scholar] [CrossRef]
- Dudzinska, W.; Lubkowska, A.; Dolegowska, B.; Safranow, K.; Jakubowska, K. Adenine, Guanine and Pyridine Nucleotides in Blood during Physical Exercise and Restitution in Healthy Subjects. Eur. J. Appl. Physiol. 2010, 110, 1155–1162. [Google Scholar] [CrossRef]
- Howarth, K.R.; LeBlanc, P.J.; Heigenhauser, G.J.F.; Gibala, M.J. Effect of Endurance Training on Muscle TCA Cycle Metabolism during Exercise in Humans. J. Appl. Physiol. (1985) 2004, 97, 579–584. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Iemitsu, M.; Katayama, K.; Ishida, K.; Kanao, Y.; Saito, M. Responses of Sex Steroid Hormones to Different Intensities of Exercise in Endurance Athletes: Sex Steroid Hormone Responses in Athletes. Exp. Physiol. 2016, 101, 168–175. [Google Scholar] [CrossRef] [PubMed]
- David, W. The Immunoassay Handbook. Theory and Applications of Ligand Binding, ELISA and Related Techniques, 4th ed.; Elsevier Science: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Voller, A.; Bidwell, D.E.; Bartlett, A. Enzyme Immunoassays in Diagnostic Medicine. Theory and Practice. Bull. World Health Organ. 1976, 53, 55–65. [Google Scholar] [PubMed]
- Ménard, S.; Cerf-Bensussan, N.; Heyman, M. Multiple Facets of Intestinal Permeability and Epithelial Handling of Dietary Antigens. Mucosal. Immunol. 2010, 3, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Rosenstein, A.Z.; Kondakov, S.E.; Rosenstein, M.Y.; Cherevko, N.A. Fundamentals of Immunodietology; Technosphere: Moscow, Russia; New York, NY, USA, 2020. [Google Scholar]
- Rosenstein, M.Y.; Rosenstein, A.Z.; Kondakov, S.E.; Cherevko, N.A. Dynamics of specific IgG to food antigens as a personalized marker of the state of the human immune system. Russ. J. Immunol. 2015, 9, 153–155. [Google Scholar]
- Al-Khelaifi, F.; Diboun, I.; Donati, F.; Botrè, F.; Alsayrafi, M.; Georgakopoulos, C.; Suhre, K.; Yousri, N.A.; Elrayess, M.A. A Pilot Study Comparing the Metabolic Profiles of Elite-Level Athletes from Different Sporting Disciplines. Sport. Med. Open 2018, 4, 2. [Google Scholar] [CrossRef]
- da Cruz, J.P.; Dos Santos, F.N.; Rasteiro, F.M.; Marostegan, A.B.; Manchado-Gobatto, F.B.; Gobatto, C.A. A Metabolomic Approach and Traditional Physical Assessments to Compare U22 Soccer Players According to Their Competitive Level. Biology 2022, 11, 1103. [Google Scholar] [CrossRef]
- Al-Khelaifi, F.; Diboun, I.; Donati, F.; Botrè, F.; Alsayrafi, M.; Georgakopoulos, C.; Yousri, N.A.; Suhre, K.; Elrayess, M.A. Metabolomics Profiling of Xenobiotics in Elite Athletes: Relevance to Supplement Consumption. J. Int. Soc. Sport. Nutr. 2018, 15, 48. [Google Scholar] [CrossRef]
- Williams, M.H. Dietary Supplements and Sports Performance: Introduction and Vitamins. J. Int. Soc. Sport. Nutr. 2004, 1, 1. [Google Scholar] [CrossRef]
- Malsagova, K.A.; Kopylov, A.T.; Pustovoyt, V.I.; Stepanov, A.A.; Enikeev, D.V.; Potoldykova, N.V.; Balakin, E.I.; Kaysheva, A.L. Pilot Study of the Metabolomic Profile of an Athlete after Short-Term Physical Activity. Data 2022, 8, 3. [Google Scholar] [CrossRef]
- A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 10 December 2022).
- Wagenmakers, A.J. Muscle Amino Acid Metabolism at Rest and during Exercise: Role in Human Physiology and Metabolism. Exerc. Sport Sci. Rev. 1998, 26, 287–314. [Google Scholar] [CrossRef]
- Quintas, G.; Reche, X.; Sanjuan-Herráez, J.D.; Martínez, H.; Herrero, M.; Valle, X.; Masa, M.; Rodas, G. Urine Metabolomic Analysis for Monitoring Internal Load in Professional Football Players. Metabolomics 2020, 16, 45. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, C.M.; Madigan, S.M.; Garcia-Perez, I.; Rankin, A.; O’ Sullivan, O.; Cotter, P.D. Distinct Microbiome Composition and Metabolome Exists across Subgroups of Elite Irish Athletes. J. Sci. Med. Sport 2020, 23, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-N.; Choi, J.Y.; Cho, Y.-O. Regular Moderate Exercise Training Can Alter the Urinary Excretion of Thiamin and Riboflavin. Nutr. Res. Pract. 2015, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Yoshizawa, F.; Nishizawa, N. Plasma Nτ -Methylhistidine Concentration Is a Sensitive Index of Myofibrillar Protein Degradation during Starvation in Rats. Biosci. Biotechnol. Biochem. 1996, 60, 501–502. [Google Scholar] [CrossRef]
- Dohm, G.L.; Williams, R.T.; Kasperek, G.J.; van Rij, A.M. Increased Excretion of Urea and N Tau -Methylhistidine by Rats and Humans after a Bout of Exercise. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1982, 52, 27–33. [Google Scholar] [CrossRef]
- Décombaz, J.; Reinhardt, P.; Anantharaman, K.; von Glutz, G.; Poortmans, J.R. Biochemical Changes in a 100 Km Run: Free Amino Acids, Urea, and Creatinine. Eur. J. Appl. Physiol. Occup. Physiol. 1979, 41, 61–72. [Google Scholar] [CrossRef]
- Rennie, M.J.; Edwards, R.H.; Krywawych, S.; Davies, C.T.; Halliday, D.; Waterlow, J.C.; Millward, D.J. Effect of Exercise on Protein Turnover in Man. Clin. Sci. 1981, 61, 627–639. [Google Scholar] [CrossRef]
- Radha, E.; Bessman, S.P. Effect of Exercise on Protein Degradation: 3-Methylhistidine and Creatinine Excretion. Biochem. Med. 1983, 29, 96–100. [Google Scholar] [CrossRef]
- Derave, W.; Everaert, I.; Beeckman, S.; Baguet, A. Muscle Carnosine Metabolism and Beta-Alanine Supplementation in Relation to Exercise and Training. Sports Med. 2010, 40, 247–263. [Google Scholar] [CrossRef]
- Hill, C.A.; Harris, R.C.; Kim, H.J.; Harris, B.D.; Sale, C.; Boobis, L.H.; Kim, C.K.; Wise, J.A. Influence of Beta-Alanine Supplementation on Skeletal Muscle Carnosine Concentrations and High Intensity Cycling Capacity. Amino Acids 2007, 32, 225–233. [Google Scholar] [CrossRef]
- Holeček, M.; Vodeničarovová, M. Effects of Histidine Load on Ammonia, Amino Acid, and Adenine Nucleotide Concentrations in Rats. Amino Acids 2019, 51, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.C.; Wise, J.A.; Price, K.A.; Kim, H.J.; Kim, C.K.; Sale, C. Determinants of Muscle Carnosine Content. Amino Acids 2012, 43, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.; Abe, H. Metabolic Transformation of Neuropeptide Carnosine Modifies Its Biological Activity. Cell. Mol. Neurobiol. 1999, 19, 163–175. [Google Scholar] [CrossRef]
- Pérez-Guisado, J.; Jakeman, P.M. Citrulline Malate Enhances Athletic Anaerobic Performance and Relieves Muscle Soreness. J. Strength Cond. Res. 2010, 24, 1215–1222. [Google Scholar] [CrossRef]
- Mutch, B.J.; Banister, E.W. Ammonia Metabolism in Exercise and Fatigue: A Review. Med. Sci. Sports Exerc. 1983, 15, 41–50. [Google Scholar] [CrossRef]
- Fitts, R.H. Cellular Mechanisms of Muscle Fatigue. Physiol. Rev. 1994, 74, 49–94. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal Muscle Fatigue: Cellular Mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef]
- Faude, O.; Kindermann, W.; Meyer, T. Lactate Threshold Concepts: How Valid Are They? Sport. Med. 2009, 39, 469–490. [Google Scholar] [CrossRef] [PubMed]
- Donovan, C.M.; Pagliassotti, M.J. Endurance Training Enhances Lactate Clearance during Hyperlactatemia. Am. J. Physiol. 1989, 257 Pt 1, E782–E789. [Google Scholar] [CrossRef]
- Donovan, C.M.; Pagliassotti, M.J. Enhanced Efficiency of Lactate Removal after Endurance Training. J. Appl. Physiol. (1985) 1990, 68, 1053–1058. [Google Scholar] [CrossRef]
- Valero, T. Mitochondrial Biogenesis: Pharmacological Approaches. Curr. Pharm. Des. 2014, 20, 5507–5509. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M. Vitamin C: Effects of Exercise and Requirements with Training. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 125–151. [Google Scholar] [CrossRef] [PubMed]
- Murguia, M.J.; Vailas, A.; Mandelbaum, B.; Norton, J.; Hodgdon, J.; Goforth, H.; Riedy, M. Elevated Plasma Hydroxyproline: A Possible Risk Factor Associated with Connective Tissue Injuries during Overuse. Am. J. Sports Med. 1988, 16, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Tofas, T.; Jamurtas, A.Z.; Fatouros, I.; Nikolaidis, M.G.; Koutedakis, Y.; Sinouris, E.A.; Papageorgakopoulou, N.; Theocharis, D.A. Plyometric Exercise Increases Serum Indices of Muscle Damage and Collagen Breakdown. J. Strength Cond. Res. 2008, 22, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Child, R.B.; Day, S.H.; Donnelly, A.E. Indices of Skeletal Muscle Damage and Connective Tissue Breakdown Following Eccentric Muscle Contractions. Eur. J. Appl. Physiol. Occup. Physiol. 1997, 75, 369–374. [Google Scholar] [CrossRef]
- Steele, D.S.; Smith, G.L.; Miller, D.J. The Effects of Taurine on Ca2+ Uptake by the Sarcoplasmic Reticulum and Ca2+ Sensitivity of Chemically Skinned Rat Heart. J. Physiol. 1990, 422, 499–511. [Google Scholar] [CrossRef]
- Ward, R.J.; Francaux, M.; Cuisinier, C.; Sturbois, X.; De Witte, P. Changes in Plasma Taurine Levels after Different Endurance Events. Amino Acids 1999, 16, 71–77. [Google Scholar] [CrossRef]
- Athanasiou, N.; Bogdanis, G.C.; Mastorakos, G. Endocrine Responses of the Stress System to Different Types of Exercise. Rev. Endocr. Metab. Disord. 2022, 15, 1–16. [Google Scholar] [CrossRef]
- Hackney, A.C.; Walz, E.A. Hormonal Adaptation and the Stress of Exercise Training: The Role of Glucocorticoids. Trends Sport Sci. 2013, 20, 165–171. [Google Scholar]
- Kozlov, V.A.; Kudaeva, O.T. Immunity and Exercise Stress. Med. Immunol. 2002, 4, 427–438. [Google Scholar]
- Pourvaghar, M.J.; Gaeini, A.A.; Ravasi, A.A.; Kordi, M.R.; Shaykh, D. AleslamThe Effects of Training Time on Serum Immunoglobulin Alterations and Cortisol Testosterone Responses in Male Athlete Students. Biol. Sport 2010, 27, 2143. [Google Scholar] [CrossRef]
- Nieman, D.C.; Nehlsen-Cannarella, S.L. The Effects of Acute and Chronic Exercise of Immunoglobulins. Sports Med. 1991, 11, 183–201. [Google Scholar] [CrossRef] [PubMed]
- Emelyanov, B.A.; Kuzmin, S.N.; Dzagurova, M.S.; Moshiashvili, I.Y.; Pershin, B.B. Reproduction of the Phenomenon of the Disappearance of Antibodies in the Experiment. Immunology 1986, 3, 58–62. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
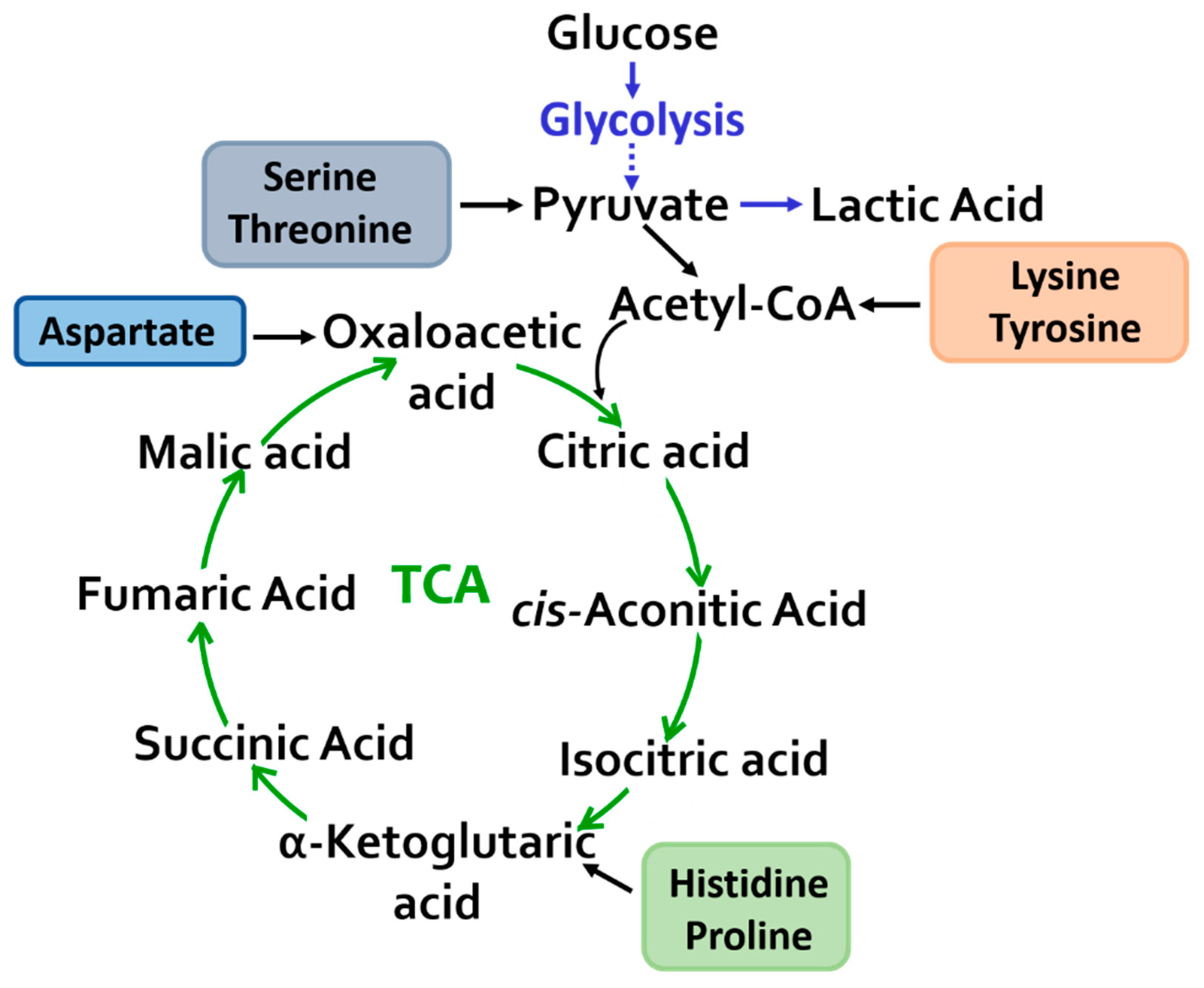{kind=link}
| Sample No. # | Age (Years) | Weight (kg) | Height (cm) | BMI (kg/m2) |
|---|---|---|---|---|
| 1 | 28 | 82.5 | 179 | 25.7 |
| 2 | 28 | 67 | 176 | 21.6 |
| 3 | 28 | 76.4 | 180 | 23.6 |
| 4 | 29 | 70.8 | 181 | 21.4 |
| 5 | 26 | 72.4 | 176 | 23.4 |
| 6 | 28 | 76.3 | 180 | 23.5 |
| 7 | 28 | 69.0 | 177 | 22.0 |
| 8 | 29 | 80.0 | 178 | 22.9 |
| 9 | 32 | 63.6 | 171 | 21.8 |
| 10 | 27 | 72.9 | 171 | 24.9 |
| 11 | 32 | 82.2 | 179 | 25.6 |
| Indicator | Unit | Mean (SD) |
|---|---|---|
| VO2 max | mL/min/kg | 64.1 ± 6.4 |
| RQ | relative units | 1.14 ± 0.1 |
| METs | relative units | 18 ± 1.9 |
| Resting heart rate | bpm | 82 ± 6.5 |
| Aerobic threshold | ||
| VO2 | mL/min | 47.2 ± 9 |
| VE | L/min | 90 ± 17.8 |
| Heart rate | bpm | 154 ± 12 |
| Anaerobic threshold | ||
| VO2 | mL/min | 58 ± 9.2 |
| VE | L/min | 138 ± 21.5 |
| Heart rate | bpm | 176 ± 7.6 |
| Maximal oxygen consumption | ||
| VE | L/min | 174 ± 11.4 |
| Heart rate | bpm | 186 ± 5.5 |
| Stage | Speed (km/h) | Time (min) |
|---|---|---|
| 1 | 5.00 | 0.00–1.40 |
| 2 | 6.00 | 1.40–3.20 |
| 3 | 7.00 | 3.20–5.00 |
| 4 | 8.00 | 5.00–6.40 |
| 5 | 8.00 | 6.40–8.20 |
| 6 | 10.00 | 8.20–9.00 |
| 7 | 11.00 | 9.00–10.40 |
| 8 | 12.00 | 10.40–12.20 |
| 9 | 13.00 | 12.20–14.00 |
| 10 | 14.00 | 14.00–15.40 |
| 11 | 15.00 | 15.40–17.20 |
| 12 | 16.00 | 17.20–19.00 |
| 13 | 17.00 | 19.00–20.40 |
| 14 | 18.00 | 20.40–22.00 |
| 15 | 19.00 | 22.00–23.40 |
| 16 | 20.00 | 23.40–25.00 |
| Recovery | 4.00 | 2.00 |
| 2.7 | 3.00 |
| No. | Allergen | No. | Allergen |
|---|---|---|---|
| 1. | Milk protein concentrate (micellar casein) | 12. | Beef protein hydrolysate (beef protein) |
| 2. | Dry egg white (egg protein) | 13. | Whey protein concentrate (whey protein) |
| 3. | Orange | 14. | Corn grits |
| 4. | Potato | 15. | Baker’s yeast |
| 5. | Peanut | 16. | Hazelnut |
| 6. | Honey | 17. | Strawberry |
| 7. | Almond | 18. | Pomegranate |
| 8. | Oat groats | 19. | Coconut |
| 9. | A mixture of brown and wild rice | 20. | Flax seed |
| 10. | Soy protein isolate (soy protein) | 21. | Wheat groats |
| 11. | Barley groats | 22. | Brewer’s yeast |
| IgG Concentration (µg/mL) | Result |
|---|---|
| 0 | Negative |
| 0–0.88 | Mild allergic reaction |
| 0.88–8.8 | Moderate allergic reaction |
| 8.8–88 | Highly allergic reaction |
| 88–880 | Very highly allergic reaction |
| No. | Metabolite | Mean Concentration (µM/L), before Loading | Mean Concentration (µM/L), after Recovery | SD before Loading | SD after Loading | Change |
|---|---|---|---|---|---|---|
| 1. | 2-ketoglutaric acid | 1.26 | 2.68 | 1.60 | 4.14 | ↑2.11 |
| 2. | 3-Methylhistidine | 0.77 | 0.13 | 1.01 | 0.17 | ↓0.17 |
| 3. | Ascorbic acid | 1.86 | 3.19 | 1.88 | 3.87 | ↑1.711 |
| 4. | Aspartic acid | 2.01 | 3.16 | 1.71 | 2.97 | ↑1.56 |
| 5. | Carnosine | 1.26 | 0.40 | 0.96 | 0.18 | ↓0.31 |
| 6. | cis-Aconitic acid | 1.29 | 3.37 | 1.38 | 4.38 | ↑2.59 |
| 7. | Citric acid | 1.25 | 4.58 | 0.09 | 0.66 | ↑3.66 |
| 8. | Citrulline | 2.87 | 1.48 | 0.76 | 0.50 | ↓0.51 |
| 9. | Creatinine | 15.14 | 8.20 | - | - | ↓0.54 |
| 10. | Histidine | 34.63 | 16.82 | - | - | ↓0.48 |
| 11. | Hydroxyproline | 2.36 | 1.10 | - | - | ↓0.46 |
| 12. | Lactic acid | 30.04 | 10.57 | 3.27 | 4.14 | ↓0.35 |
| 13. | Lysine | 14.97 | 5.32 | - | - | ↓0.35 |
| 14. | Ornithine | 5.07 | 1.91 | - | - | ↓0.37 |
| 15. | Oxalic acid | 5.33 | 3.13 | 3.22 | 1.38 | ↓0.58 |
| 16. | Proline | 61.74 | 33.69 | - | - | ↓0.54 |
| 17. | Serine | 25.67 | 46.73 | 9.84 | 14.37 | ↑1.82 |
| 18. | Succinic acid | 0.39 | 0.18 | 0.30 | 0.25 | ↓0.47 |
| 19. | Taurine | 14.51 | 28.83 | - | - | ↑1.98 |
| 20. | Threonine | 9.64 | 18.68 | - | - | ↑1.93 |
| 21. | Tyrosine | 17.03 | 6.90 | 1.60 | 4.14 | ↓0.40 |
| Hormones | ||||||
| 22. | 11-Deoxycortisol | – | – | – | – | ↑1.22 |
| 23. | Cortisol | – | – | – | – | ↓0.47 |
| 24. | Estriol (E3) | – | – | – | – | ↑1.43 |
| 25. | Dehydroepiandrosterone | – | – | – | – | ↓0.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malsagova, K.A.; Kopylov, A.T.; Stepanov, A.A.; Enikeev, D.V.; Potoldykova, N.V.; Balakin, E.I.; Pustovoyt, V.I.; Kaysheva, A.L. Molecular Profiling of Athletes Performing High-Intensity Exercises in Extreme Environments. Sports 2023, 11, 36. https://doi.org/10.3390/sports11020036
Malsagova KA, Kopylov AT, Stepanov AA, Enikeev DV, Potoldykova NV, Balakin EI, Pustovoyt VI, Kaysheva AL. Molecular Profiling of Athletes Performing High-Intensity Exercises in Extreme Environments. Sports. 2023; 11(2):36. https://doi.org/10.3390/sports11020036
Chicago/Turabian StyleMalsagova, Kristina A., Arthur T. Kopylov, Alexander A. Stepanov, Dmitry V. Enikeev, Natalia V. Potoldykova, Evgenii I. Balakin, Vasiliy I. Pustovoyt, and Anna L. Kaysheva. 2023. "Molecular Profiling of Athletes Performing High-Intensity Exercises in Extreme Environments" Sports 11, no. 2: 36. https://doi.org/10.3390/sports11020036