
Vegetation Affects the Responses of Canopy Spider Communities to Elevation Gradients on Changbai Mountain, China

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
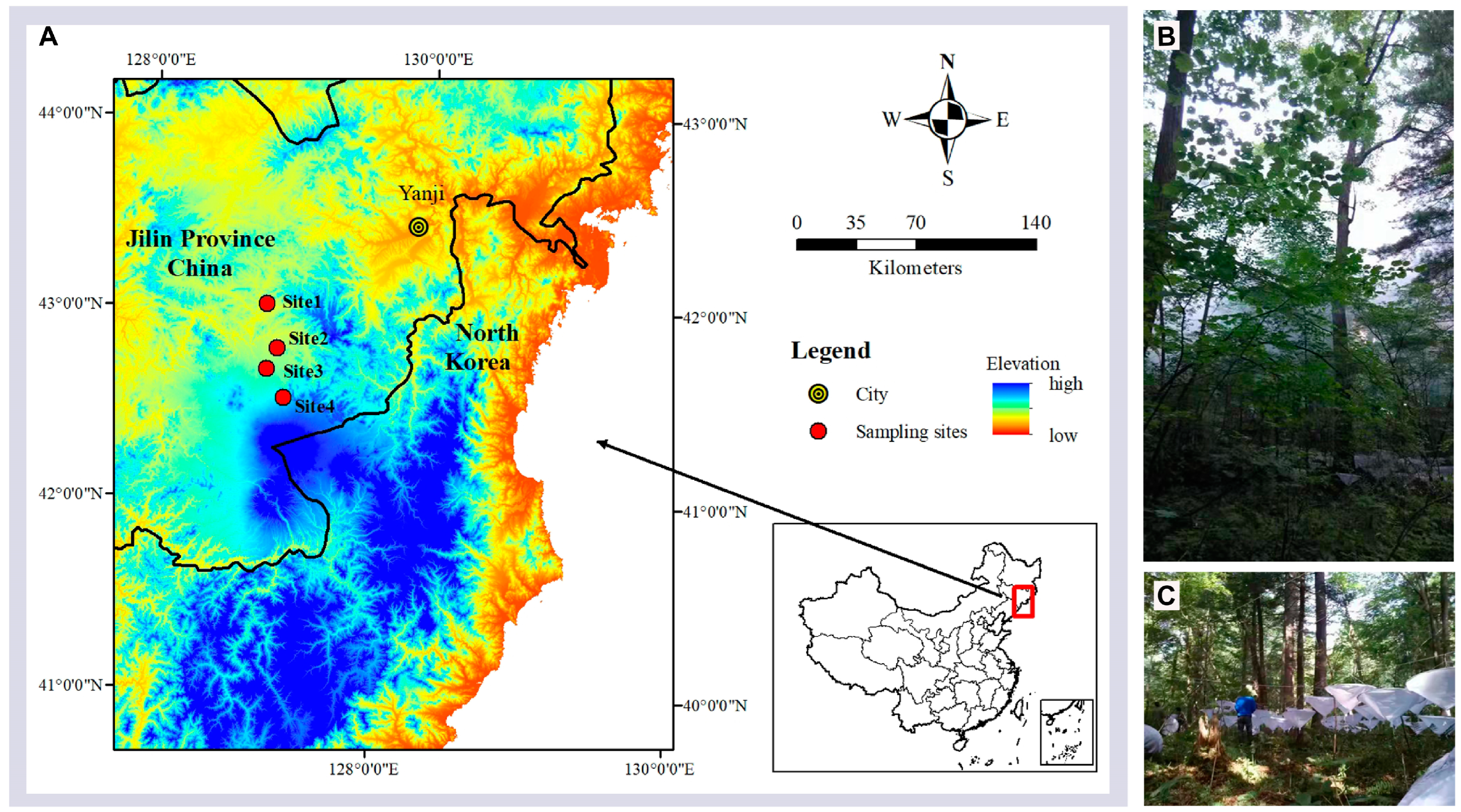
2.1. Study Area
2.2. Experimental Design and Spider Sampling
2.3. Spider Identification and Statistical Analyses
3. Results
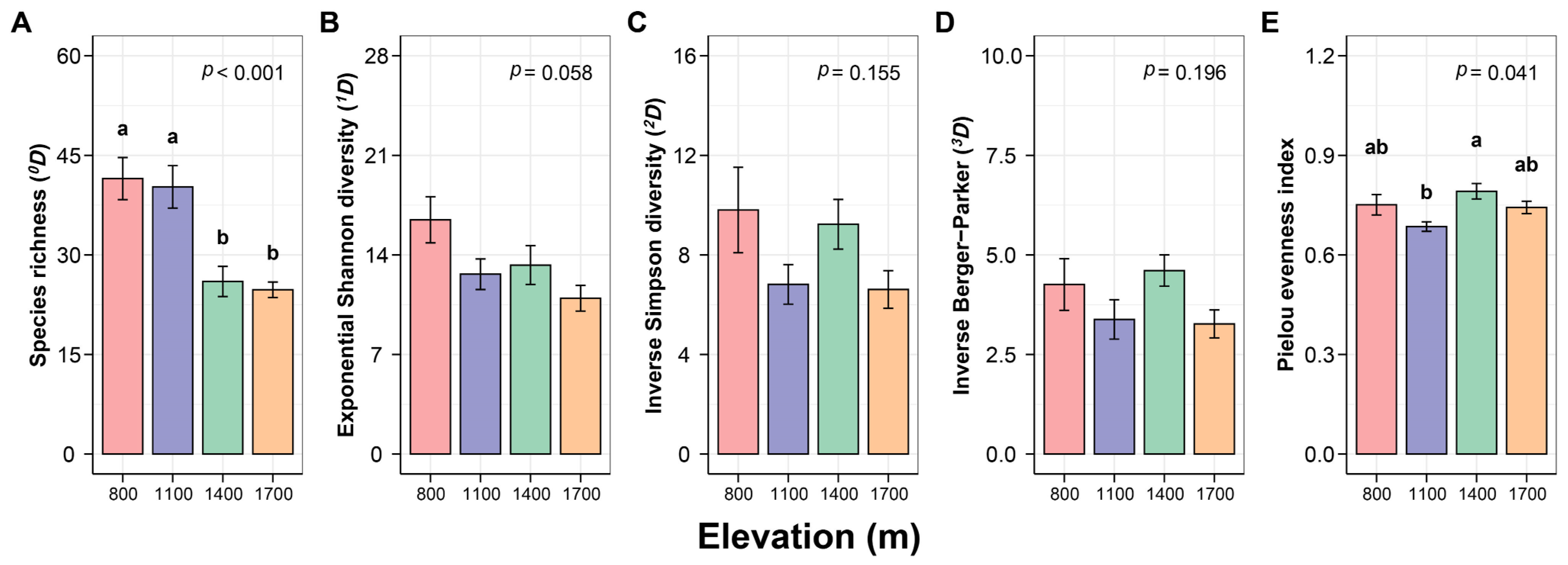
3.1. Response of Canopy Spider Diversity to Changes in Elevation
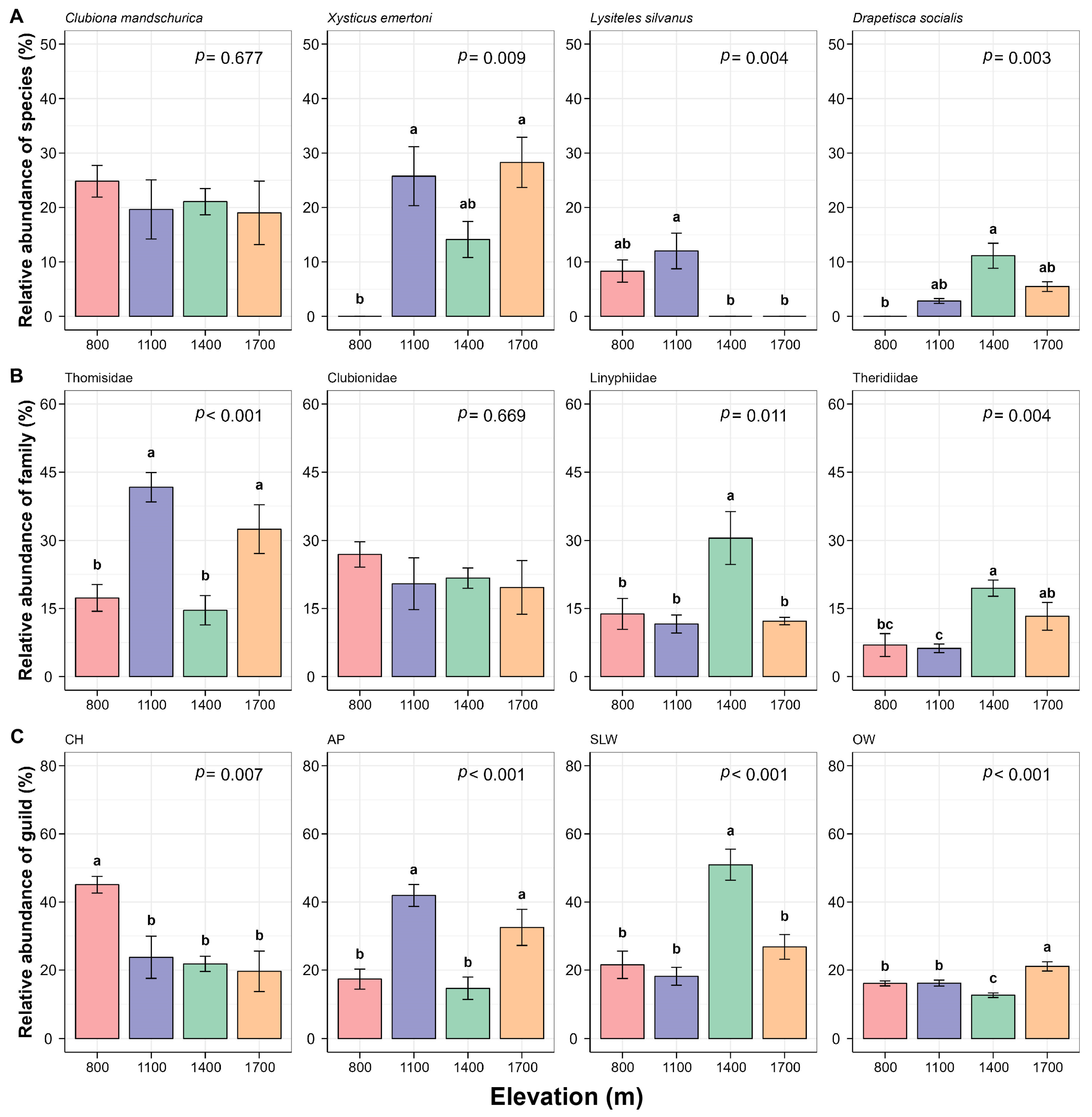
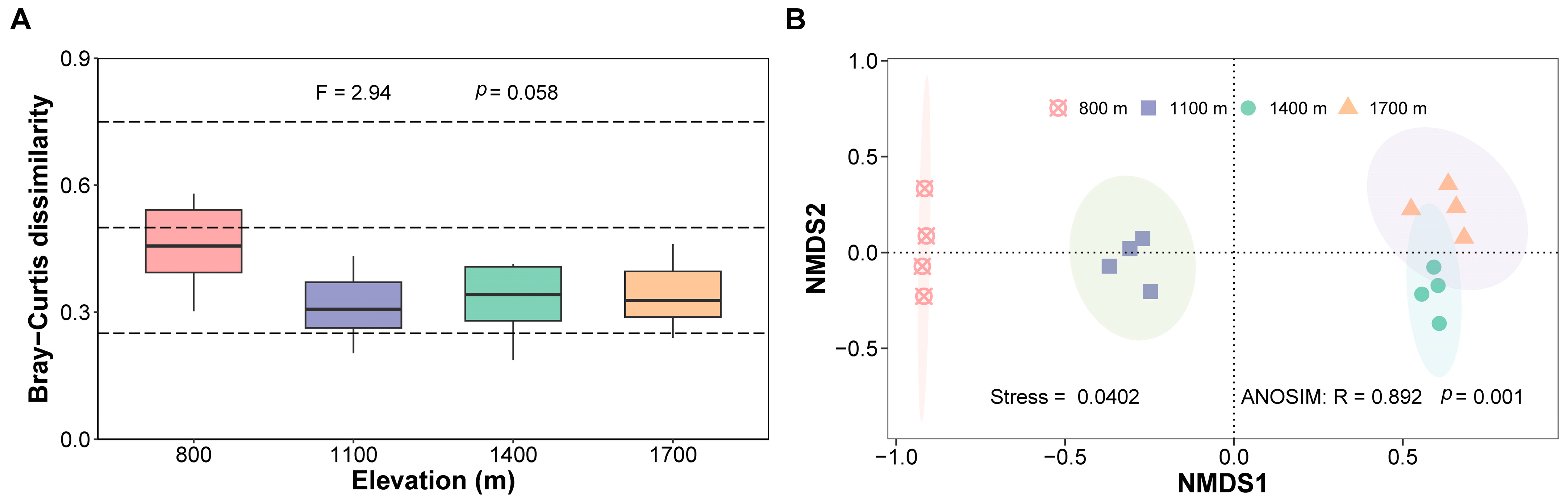
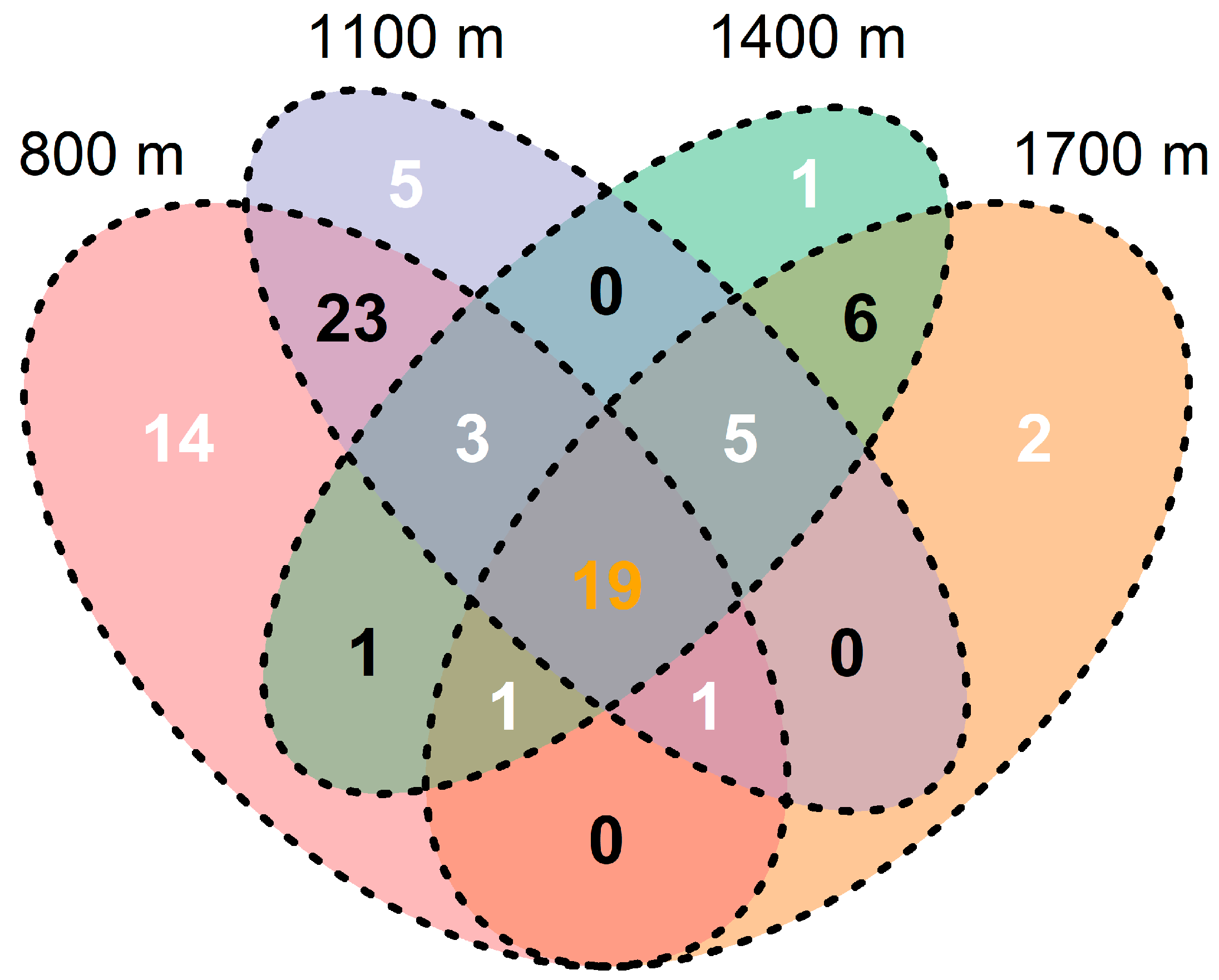
3.2. Response of Community Abundance and Composition of Canopy Spiders to Changes in Elevation
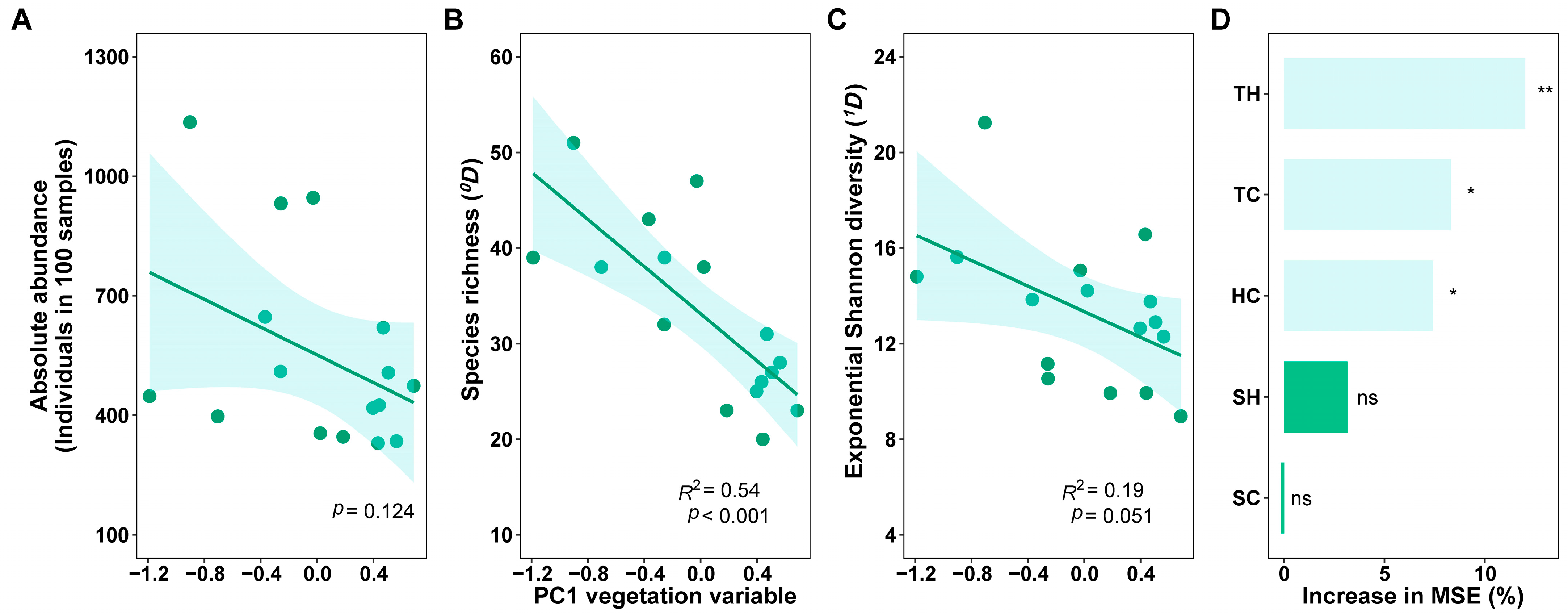
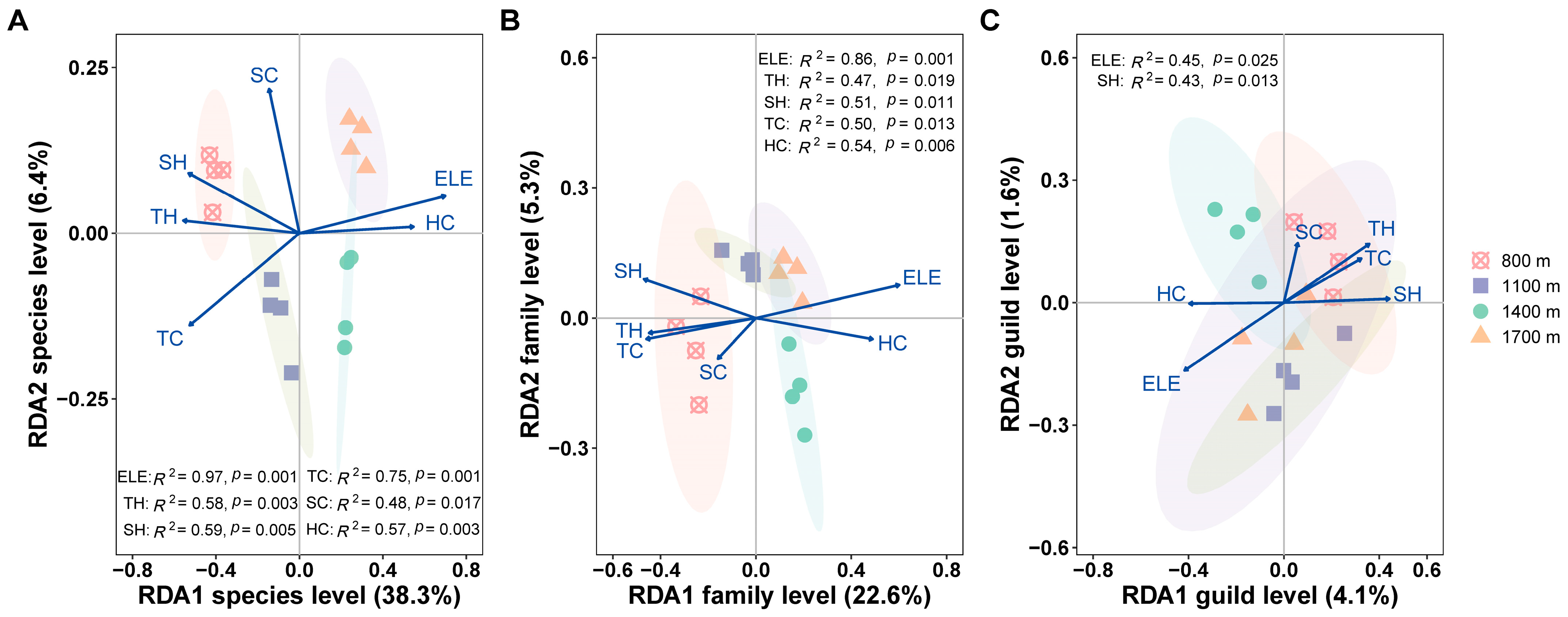
3.3. Effects of Habitat Factors on the Canopy Spider Community
4. Discussion
4.1. Difference in Diversity of Canopy Spiders
4.2. Changes in Abundance and Composition of Canopy Spiders
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quintero, I.; Jetz, W. Global elevational diversity and diversification of birds. Nature 2018, 555, 246–250. [Google Scholar] [CrossRef]
- Zizka, A.; Antonelli, A. Mountains of diversity. Nature 2018, 555, 173–174. [Google Scholar] [CrossRef]
- Cancino-López, R.J.; Moreno, C.E.; Contreras-Ramos, A. Diversity of lacewings (Neuroptera) in an altitudinal gradient of the Tacaná Volcano, Southern Mexico. Insects 2022, 13, 652. [Google Scholar] [CrossRef]
- Bai, F.; Sang, W.G.; Axmacher, J. Forest vegetation responses to climate and environmental change: A case study from Changbai Mountain, NE China. Forest Ecol. Manag. 2011, 262, 2052–2060. [Google Scholar] [CrossRef]
- McCain, C.M. Elevational gradients in diversity of small mammals. Ecology 2005, 86, 366–372. [Google Scholar] [CrossRef]
- Carvalho-Rocha, V.; Peres, C.A.; Neckel-Oliveira, S. Habitat amount and ambient temperature dictate patterns of anuran diversity along a subtropical elevational gradient. Divers. Distrib. 2021, 27, 344–359. [Google Scholar] [CrossRef]
- Dehling, D.M.; Fritz, S.A.; Töpfer, T.; Päckert, M.; Estler, P.; Böhning-Gaese, K.; Schleuning, M. Functional and phylogenetic diversity and assemblage structure of frugivorous birds along an elevational gradient in the tropical Andes. Ecography 2014, 37, 1047–1055. [Google Scholar] [CrossRef]
- Sanders, N.J. Elevational gradients in ant species richness area, geometry, and Rapoport’s rule. Ecography 2002, 25, 25–32. [Google Scholar] [CrossRef]
- Bärtschi, F.; McCain, C.M.; Ballesteros-Mejia, L.; Kitching, I.J.; Beerli, N.; Beck, J. Elevational richness patterns of sphingid moths support area effects over climatic drivers in a near-global analysis. Global Ecol. Biogeogr. 2019, 28, 917–927. [Google Scholar] [CrossRef]
- Chen, A.P.; Li, Z.J.; Zheng, Y.F.; Zhan, J.Y.; Yang, B.L.; Yang, Z.F. Decreasing species richness with increase in elevation and positive Rapoport effects of Crambidae (Lepidoptera) on Mount Taibai. Insects 2022, 13, 1125. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.K.; Hemp, A.; Appelhans, T.; Behler, C.; Clseen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef]
- Binkenstein, J.; Klein, A.M.; Assmann, T.; Buscot, F.; Erfmeier, A.; Ma, K.P.; Pietsch, K.A.; Schmidt, K.; Scholten, T.; Wubet, T.; et al. Multi-trophic guilds respond differently to changing elevation in a subtropical forest. Ecography 2018, 41, 1013–1023. [Google Scholar] [CrossRef]
- Lowman, M.D.; Moffett, M. The ecology of tropical rain forest canopies. Trends Ecol. Evol. 1993, 8, 104–107. [Google Scholar] [CrossRef]
- Ozanne, C.M.P.; Anhuf, D.; Boulter, S.L.; Keller, M.; Kitching, R.L.; Körner, C.; Meinzer, F.C.; Mitchell, A.W.; Nakashizuka, T.; Silva Dias, P.L.; et al. Biodiversity meets the atmosphere: A global view of forest canopies. Science 2003, 301, 183–186. [Google Scholar] [CrossRef]
- Pennisi, E. Forest research: Sky-high experiments. Science 2005, 309, 1314–1315. [Google Scholar] [CrossRef]
- Lowman, M.D.; Wittman, P.K. Forest canopies methods, hypotheses, and future directions. Annu. Rev. Ecol. Syst. 1996, 27, 55–81. [Google Scholar] [CrossRef]
- Moran, V.C.; Southwood, T.R.E. The guild composition of arthropod communities in trees. J. Anim. Ecol. 1982, 51, 289–306. [Google Scholar] [CrossRef]
- Zheng, G.; Li, S.Q. The biodiversity of arthropods from forest canopies. Acta Zootaxonomica Sinica 2013, 38, 178–182. [Google Scholar]
- Vaca-Sánchez, M.S.; Maldonado-López, Y.; González-Rodríguez, A.; Oyama, K.; Fernandes, G.W.; Fagundes, M.; López-Barbosa, E.C.; Aguilar-Peralta1, J.S.; Cuevas-Reyes, P. Canopy arthropod diversity associated with Quercus laurina: Importance of an oak species diversity gradient on abundance, species richness and guild composition. J. Insect Conserv. 2021, 25, 859–874. [Google Scholar] [CrossRef]
- Birkhofer, K.; Wolters, V. The global relationship between climate, net primary production and the diet of spiders. Global Ecol. Biogeogr. 2012, 21, 100–108. [Google Scholar] [CrossRef]
- Ferretti, N.E.; Arnedo, M.A.; Gonzalez, A. Impact of climate change on spider species distribution along the La Plata River Basin, southern South America: Projecting future range shifts for the genus Stenoterommata (Araneae, Mygalomorphae, Nemesiidae). Ann. Zool. Fenn. 2018, 55, 123–133. [Google Scholar] [CrossRef]
- Duan, M.C.; Hu, W.H.; Liu, Y.H.; Yu, Z.R.; Li, X.; Wu, P.L.; Zhang, F.; Shi, H.L.; Baudry, J. The influence of landscape alterations on changes in ground beetle (Carabidae) and spider (Araneae) functional groups between 1995 and 2013 in an urban fringe of China. Sci. Total Environ. 2019, 689, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef]
- Lawton, J.H. Plant architecture and the diversity of phytophagous insects. Annu. Rev. Entomol. 1983, 28, 23–39. [Google Scholar] [CrossRef]
- Basset, Y. The taxonomic composition of the arthropod fauna associated with an Australian rainforest tree. Aust. J. Zool. 1991, 39, 171–190. [Google Scholar] [CrossRef]
- Katayama, M.; Kishimoto-Yamada, K.; Tanaka, H.O.; Endo, T.; Hashimoto, Y.; Yamane, S.; Itioka, T. Negative correlation between ant and spider abundances in the canopy of a Bornean tropical rain forest. Biotropica 2015, 47, 363–368. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Zou, Y.; Sang, W.G.; Bai, F.; Axmacher, J.C. Relationships between plant diversity and the abundance and α-diversity of predatory ground beetles (Coleoptera: Carabidae) in a mature Asian temperate forest ecosystem. PLoS ONE 2013, 8, e82792. [Google Scholar] [CrossRef]
- Zou, Y.; Sang, W.G.; Zhou, H.C.; Huang, L.Y.; Axmacher, J.C. Altitudinal diversity patterns of ground beetles (Coleoptera: Carabidae) in the forests of Changbai Mountain, Northeast China. Insect Conserv. Diver. 2014, 7, 161–171. [Google Scholar] [CrossRef]
- Gao, W.T.; Chen, Y.B.; Meng, Q.F. Faunal composition and vertical distribution characteristics of Cerambycidae in Changbai Mountain Reserve. Sci. Silvae Sin. 2014, 50, 89–96. [Google Scholar]
- Jiang, Y.F.; Yin, X.Q.; Wang, F.B. Composition and spatial distribution of soil mesofauna along an elevation gradient on the north slope of the Changbai Mountains, China. Pedosphere 2015, 25, 811–824. [Google Scholar] [CrossRef]
- Yang, M.Y. A Comparative Study on the Diversity Composition of Oribatida on the North Slope and the South Slope of Changbai Mountain Nature Reserve. Master Dissertation, Beihua University, Jilin, China, 2020. [Google Scholar]
- Sun, X.; Deharveng, L.; Bedos, A.; Chang, L.; Scheu, S.; Wu, D.H. Changes in diversity and body size of Onychiurinae (Collembola: Onychiuridae) along an altitudinal gradient in Changbai Mountain, China. Soil Ecol. Lett. 2020, 2, 230–239. [Google Scholar] [CrossRef]
- Xie, Z.J.; Sun, X.; Lux, J.; Chen, T.W.; Potapov, M.; Wu, D.H.; Scheu, S. Drivers of Collembola assemblages along an altitudinal gradient in northeast China. Evol. Ecol. 2022, 12, e8559. [Google Scholar] [CrossRef]
- Wu, Y.G.; Xie, Z.J.; Wan, Z.M.; Ji, Q.-Q.; Yang, J.J.; Chen, T.-W.; Wu, D.H.; Scheu, S. Elevational changes in canopy Collembola community composition are primarily driven by species turnover on Changbai Mountain, northeastern China. Biodivers. Conserv. 2023, 32, 4853–4872. [Google Scholar] [CrossRef]
- Stone, R. A threatened nature reserve breaks down Asian borders. Science 2006, 313, 1379–1380. [Google Scholar] [CrossRef]
- He, H.S.; Hao, Z.Q.; Mladenoff, D.J.; Shao, G.F.; Hu, Y.M.; Chang, Y. Simulating forest ecosystem response to climate warming incorporating spatial effects in north-eastern China. J. Biogeogr. 2005, 32, 2043–2056. [Google Scholar] [CrossRef]
- Chen, L.; Wu, S.H.; Pan, T. Variability of climate–growth relationships along an elevation gradient in the Changbai Mountain, northeastern China. Trees 2011, 25, 1133–1139. [Google Scholar] [CrossRef]
- Chen, L.Z.; Bao, X.C.; Li, C.G. Major forests in various vertical zones on northern slope of Changbai Mountains of Jilin Province. Acta Phytoecol. Geobot. Sin. 1964, 2, 207–225. [Google Scholar]
- Sang, W.G.; Bai, F. Vascular diversity patterns of forest ecosystem before and after a 43-year interval under changing climate conditions in the Changbaishan Nature Reserve, northeastern China. Plant Ecol. 2009, 201, 115–130. [Google Scholar] [CrossRef]
- Otto, S.; Floren, A. The spider fauna (Araneae) of tree canopies in Białowieża Forest. Fragm. Faun. 2007, 50, 57–70. [Google Scholar] [CrossRef]
- Zheng, G.; Li, S.Q.; Yang, X.D. Spider diversity in canopies of Xishuangbanna rainforest (China) indicates an alarming juggernaut effect of rubber plantations. Forest Ecol. Manag. 2015, 338, 200–207. [Google Scholar] [CrossRef]
- Sørensen, L.L. Composition and diversity of the spider fauna in the canopy of a montane forest in Tanzania. Biodivers. Conserv. 2004, 13, 437–452. [Google Scholar] [CrossRef]
- Floren, A.; Otto, S.; Linsenmair, K.E. Do spider communities in primary forests differ from those in forest-plantations? A canopy study in the Bialowieza forest (Poland). In Canopy Arthropod Research in Europe: Basic and Applied Studies from the High Frontier; Floren, A., Schmidl, J., Eds.; Bioform Entomology: Nuremberg, Germany, 2008; pp. 489–506. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975. [Google Scholar]
- Chen, H.B. R Package Version 1.7.3. VennDiagram: Generate High-Resolution Venn and Euler Plots. 2022. Available online: https://cran.r-project.org/package=VennDiagram (accessed on 20 July 2023).
- Axmacher, J.C.; Holtmann, G.; Scheuermann, L.; Brehm, G.; Müller-Hohenstein, K.; Fiedler, K. Diversity of geometrid moths (Lepidoptera: Geometridae) along an Afrotropical elevational rainforest transect. Divers. Distrib. 2004, 10, 293–302. [Google Scholar] [CrossRef]
- Sivault, E.; Amick, P.K.; Armstrong, K.N.; Novotny, V.; Sam, K. Species richness and assemblages of bats along a forest elevational transect in Papua New Guinea. Biotropica 2023, 55, 81–94. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. RandomForest: Breiman and Cutler’s Random Forests for Classification and Regression. R Package Version 4.7-1.1. randomForest: Breiman and Cutler’s Random Forests for Classification and Regression. 2022. Available online: https://cran.r-project.org/package=randomforest (accessed on 20 July 2023).
- He, Z.B.; Chen, L.F.; Zhao, W.Z.; Liu, J.L.; Song, M.D.; Kong, J.Q.; Gao, Y. Co-ordinated elevational diversity patterns for soil bacteria, fungi, and plants in alkaline soils of arid northwestern China. Eur. J. Soil Sci. 2022, 73, e13209. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Solymos, P.; Stevens, H.M.H.; Szöcs, E.; et al. R Package Version 2.6-4. Vegan: Community Ecology Package. 2022. Available online: https://cran.r-project.org/package=vegan (accessed on 20 July 2023).
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. R Package Version 3.4.2. Ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. 2022. Available online: https://cran.r-project.org/package=ggplot2 (accessed on 20 July 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Russell-Smith, A.; Stork, N.E. Abundance and diversity of spiders from the canopy of tropical rainforests with particular reference to Sulawesi, Indonesia. J. Trop. Ecol. 1994, 10, 545–558. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Colwell, R.K.; Rahbek, C.; Gotelli, N.J. The mid-domain effect: There’s a baby in the bathwater. Am. Nat. 2005, 166, E149–E154. [Google Scholar] [CrossRef]
- McCain, C.M.; Grytnes, J.-A. Elevational gradients in species richness. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2010. [Google Scholar]
- Sanders, N.J.; Rahbek, C. The patterns and causes of elevational diversity gradients. Ecography 2012, 35, 1. [Google Scholar] [CrossRef]
- Burwell, C.J.; Nakamura, A. Distribution of ant species along an altitudinal transect in continuous rainforest in subtropical Queensland, Australia. Mem. Queensl. Mus. 2011, 55, 391–411. [Google Scholar]
- Kwon, T.-S.; Kim, S.-S.; Chun, J.H. Pattern of ant diversity in Korea: An empirical test of Rapoport’s altitudinal rule. J. Asia-Pacific Entomol. 2014, 17, 161–167. [Google Scholar] [CrossRef]
- Nunes, C.A.; Quintino, A.V.; Constantino, R.; Negreiros, D.; Junior, R.R.; Fernandes, G.W. Patterns of taxonomic and functional diversity of termites along a tropical elevational gradient. Biotropica 2017, 49, 186–194. [Google Scholar] [CrossRef]
- Finch, O.D.; Blick, T.; Schuldt, A. Macroecological patterns of spider species richness across Europe. Biodivers. Conserv. 2008, 17, 2849–2868. [Google Scholar] [CrossRef]
- Downie, I.S.; Butterfield, J.E.L.; Coulson, J.C. Habitat preferences of sub-montane spiders in northem England. Ecography 1995, 18, 51–61. [Google Scholar] [CrossRef]
- Purchart, L.; Tuf, I.H.; Hula, V.; Suchomel, J. Arthropod assemblages in Norway spruce monocultures during a forest cycle—A multi-taxa approach. Forest Ecol. Manag. 2013, 306, 42–51. [Google Scholar] [CrossRef]
- Mcnett, B.J.; Rypstra, A.L. Habitat selection in a large orb-weaving spider: Vegetational complexity determines site selection and distribution. Ecol. Entomol. 2000, 25, 423–432. [Google Scholar] [CrossRef]
- Ford, H.; Evans, B.; Van Klink, R.; Skov, M.W.; Garbutt, A. The importance of canopy complexity in shaping seasonal spider and beetle assemblages in saltmarsh habitats. Ecol. Entomol. 2017, 42, 145–155. [Google Scholar] [CrossRef]
- Ulyshen, M.D. Arthropod vertical stratification in temperate deciduous forests: Implications for conservation-oriented management. Forest Ecol. Manag. 2011, 261, 1479–1489. [Google Scholar] [CrossRef]
- Zhao, S.Q.; Fang, J.Y.; Zong, Z.J.; Zhu, B.; Shen, H.H. Composition, structure and species diversity of plant communities along an altitudinal gradient on the northern slope of Mt. Changbai, Northeast China. Biodivers. Sci. 2004, 12, 164–173. [Google Scholar] [CrossRef]
- Otto, C.; Svensson, B.S. Structure of communities of ground-living spiders along altitudinal gradients. Holarct. Ecol. 1982, 5, 35–47. [Google Scholar] [CrossRef]
- Chatzaki, M.; Lymberakis, P.; Markakis, G.; Mylonas, M. The distribution of ground spiders (Araneae, Gnaphosidae) along the altitudinal gradient of Crete, Greece: Species richness, activity and altitudinal range. J. Biogeogr. 2005, 32, 813–831. [Google Scholar] [CrossRef]
- Legendre, P.; Borcard, D.; Peres-Neto, P.R. Analyzing beta diversity: Partitioning the spatial variation of community composition data. Ecol. Monogr. 2005, 75, 435–450. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Comita, L.S.; Chase, J.M.; Sanders, N.J.; Swenson, N.G.; Crist, T.O.; Stegen, J.C.; Vellend, M.; Boyle, B.; Anderson, M.J.; et al. Disentangling the drivers of β diversity along latitudinal and elevational gradients. Science 2011, 333, 1755–1758. [Google Scholar] [CrossRef]
- Blandenier, G. Ballooning of spiders (Araneae) in Switzerland, general results from an eleven-year survey. Arachnology 2009, 14, 308–316. [Google Scholar] [CrossRef]
- Wu, P.F.; Cui, S.Y.; Ali, A.; Zheng, G. Advances in spider ballooning research. Biodivers. Sci. 2021, 29, 517–530. [Google Scholar] [CrossRef]
- Koponen, S. Communities of ground-living spiders in six habitats on a mountain in Quebec, Canada. Holarct. Ecol. 1987, 10, 278–285. [Google Scholar] [CrossRef]
- Wise, D.H. Spiders in Ecological Webs; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Entling, W.; Schmidt, M.H.; Bacher, S.; Brandl, R.; Nentwig, W. Niche properties of Central European spiders: Shading, moisture and the evolution of the habitat niche. Global Ecol. Biogeogr. 2007, 16, 440–448. [Google Scholar] [CrossRef]
- Muff, P.; Kropf, C.; Frick, H.; Nentwig, W.; Schmidt-Entling, M.H. Co-existence of divergent communities at natural boundaries: Spider (Arachnida: Araneae) diversity across an alpine timberline. Insect Conserv. Diver. 2009, 2, 36–44. [Google Scholar] [CrossRef]
- Cardoso, P.; Pekár, S.; Jocqué, R.; Coddington, J.A. Global patterns of guild composition and functional diversity of spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef]
- Szewczyk, T.; McCain, C.M. A systematic review of global drivers of ant elevational diversity. PLoS ONE 2016, 11, e0155404. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Romdal, T.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–219. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Site 1 | Site 2 | Site 3 | Site 4 | p-Value |
|---|---|---|---|---|---|
| elevation (m) | 800 | 1100 | 1400 | 1700 | / |
| tree height (TH) (m) | 26.3 ± 2.4 a | 23.0 ± 0.4 ab | 20.5 ± 0.5 ab | 19.5 ± 0.5 b | 0.021 |
| shrub height (SH) (m) | 5.4 ± 1.3 a | 3.3 ± 0.7 ab | 0.9 ± 0.1 c | 2.0 ± 0.5 bc | 0.002 |
| tree coverage (TC) (%) | 80.0 ± 2.9 a | 81.3 ± 1.5 a | 74.5 ± 0.5 b | 68.3 ± 1.2 c | <0.001 |
| shrub coverage (SC) (%) | 57.5 ± 4.3 a | 22.5 ± 6.0 c | 35.0 ± 8.7 bc | 47.5 ± 7.5 ab | 0.018 |
| herb coverage (HC) (%) | 52.5 ± 10.5 b | 60.0 ± 4.1 b | 87.5 ± 2.5 a | 82.5 ± 4.8 a | 0.005 |
| dominant tree species | Pinus koraiensis, Tilia amurensis and Populus cathayana | P. koraiensis, T. amurensis and Abies nephrolepis | A. nephrolepis, Betula ermanii, Picea jezoensis and Larix olgensis | A. nephrolepis and L. olgensis | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.; Xiang, L.; Zhao, Q.; Cui, S.; Ali, A.; Wu, D.; Zheng, G. Vegetation Affects the Responses of Canopy Spider Communities to Elevation Gradients on Changbai Mountain, China. Insects 2024, 15, 154. https://doi.org/10.3390/insects15030154
Wu P, Xiang L, Zhao Q, Cui S, Ali A, Wu D, Zheng G. Vegetation Affects the Responses of Canopy Spider Communities to Elevation Gradients on Changbai Mountain, China. Insects. 2024; 15(3):154. https://doi.org/10.3390/insects15030154
Chicago/Turabian StyleWu, Pengfeng, Lingxu Xiang, Qiang Zhao, Shuyan Cui, Abid Ali, Donghui Wu, and Guo Zheng. 2024. "Vegetation Affects the Responses of Canopy Spider Communities to Elevation Gradients on Changbai Mountain, China" Insects 15, no. 3: 154. https://doi.org/10.3390/insects15030154