
Changes in the Range of Four Advantageous Grasshopper Habitats in the Hexi Corridor under Future Climate Conditions
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
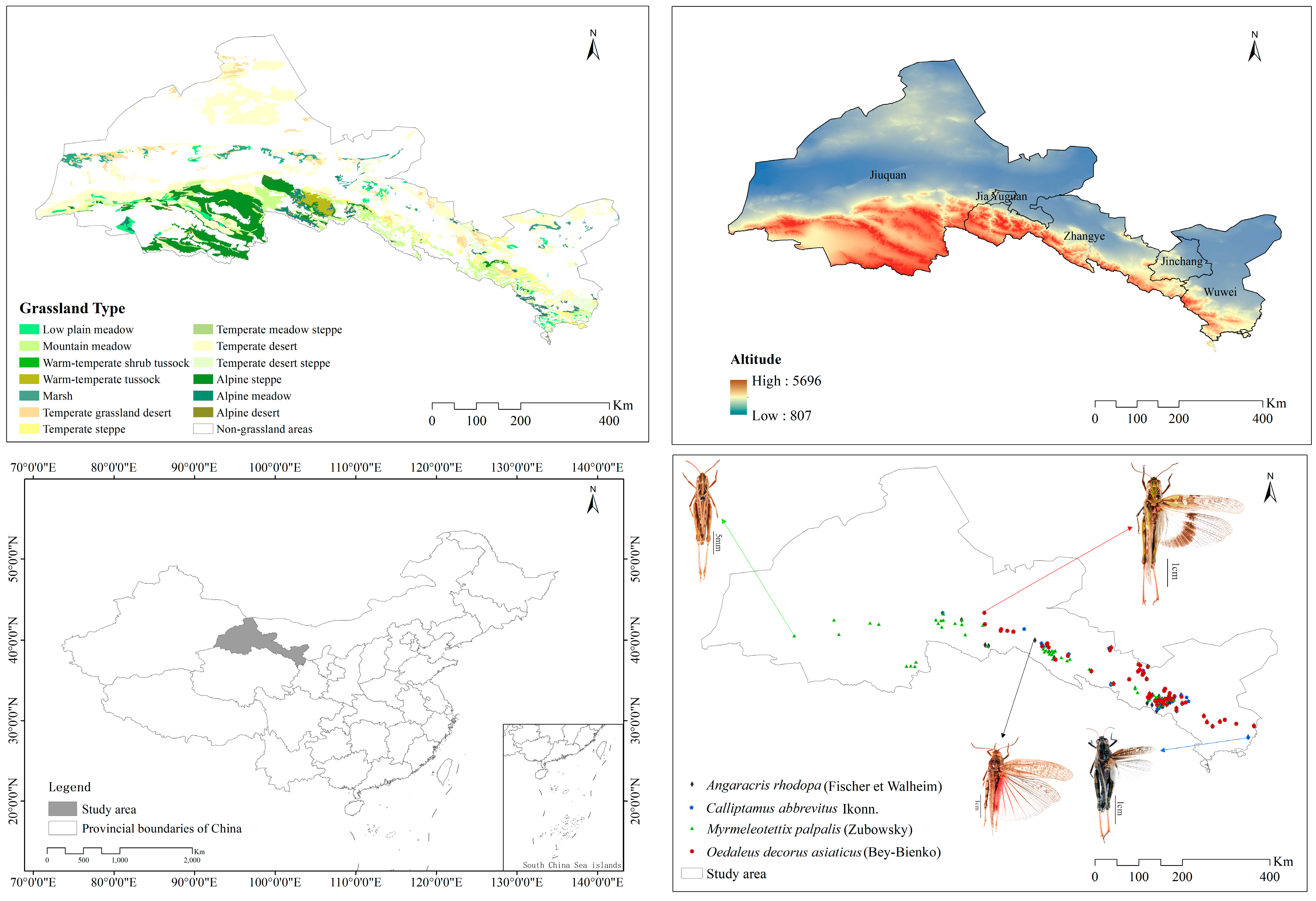
2.1. Study Area
2.2. Data Acquisition and Processing
2.2.1. Grasshopper Survey Data
2.2.2. Environmental Variables
2.2.3. Environment Variable De-Correlation
2.3. MaxEnt Model Runs
3. Results
3.1. Accuracy of the MaxEnt Model
3.2. Effects of Major Environmental Variables on the Distribution of Grasshoppers
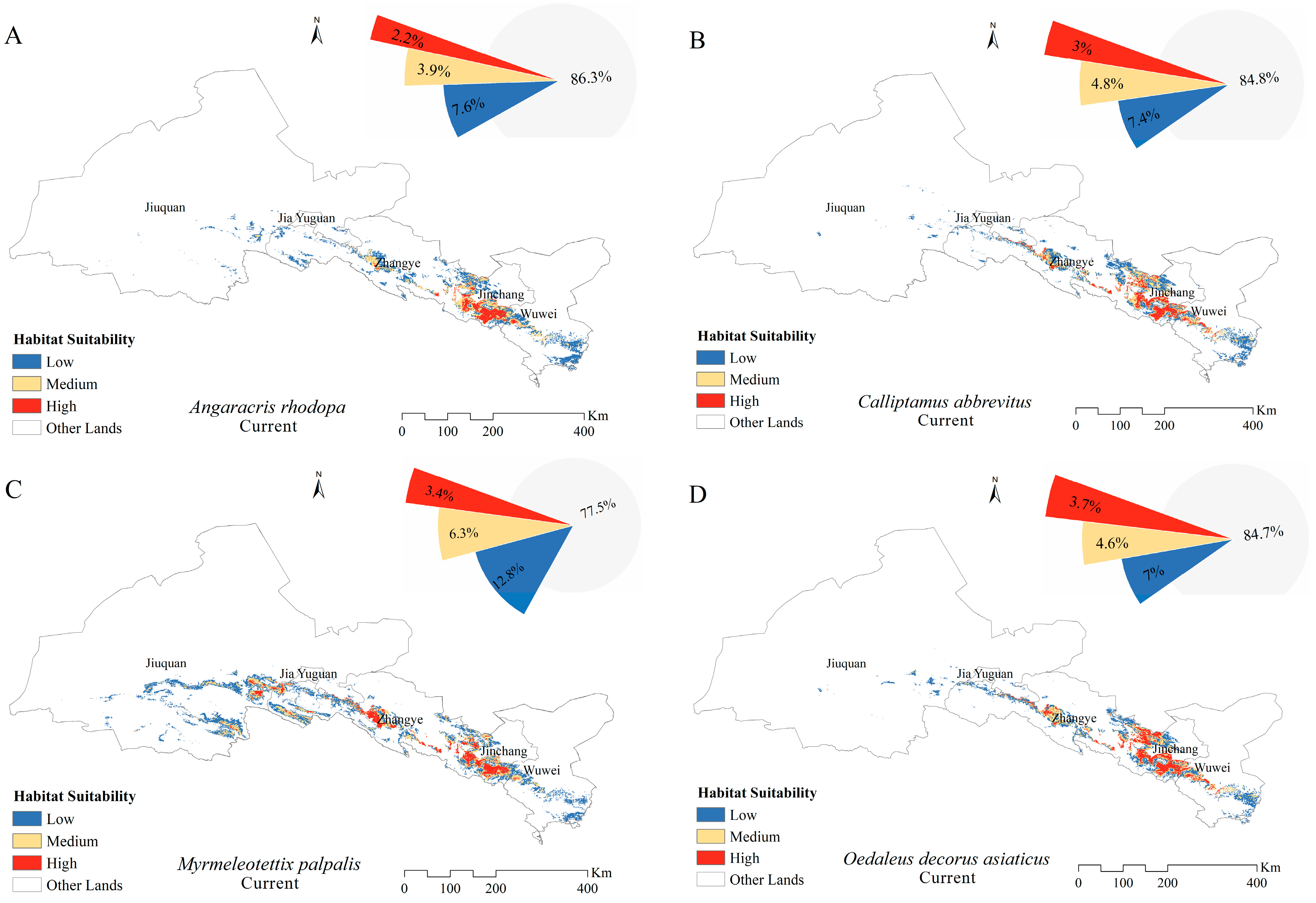
3.3. Distribution and Size of Suitable Areas for Grasshoppers in the Current Climate
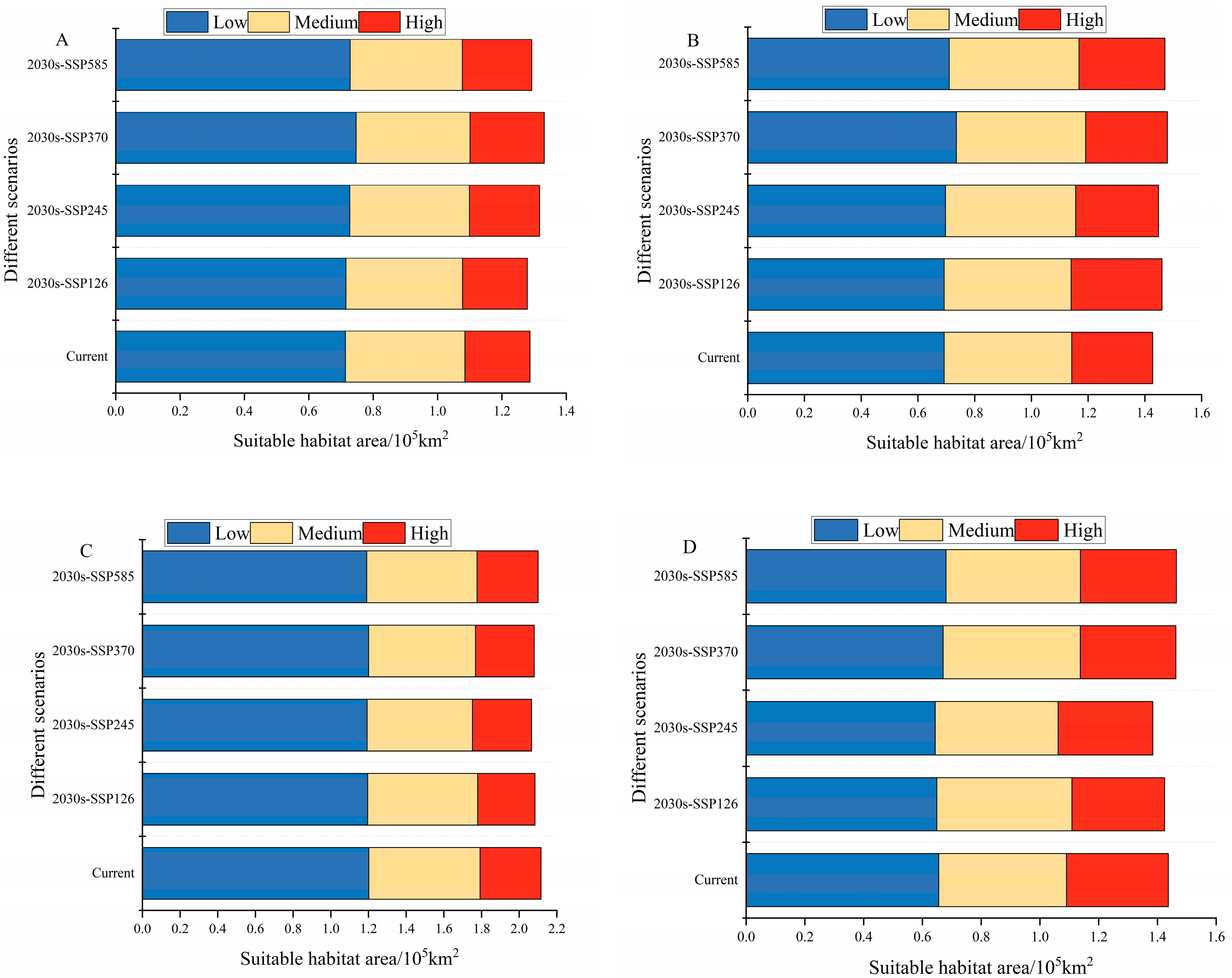
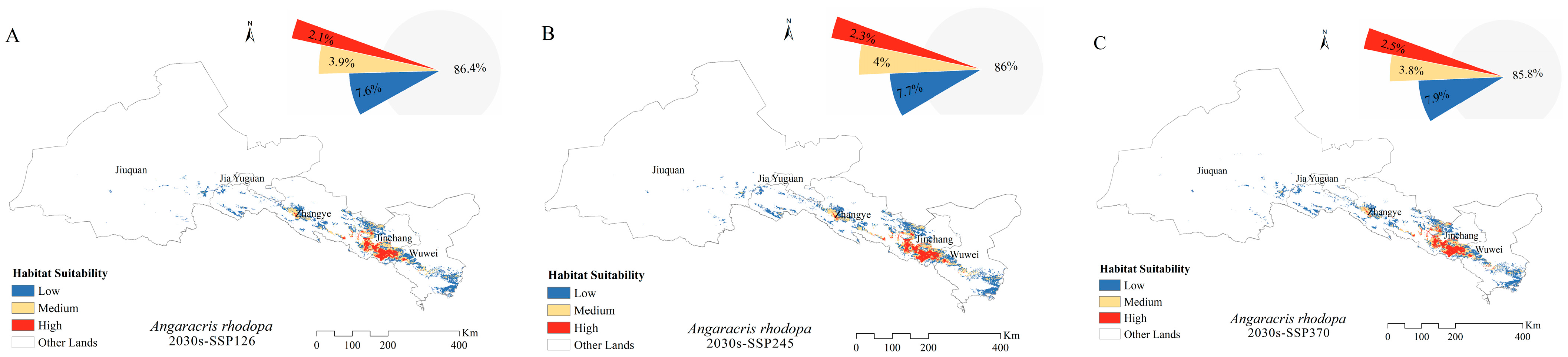
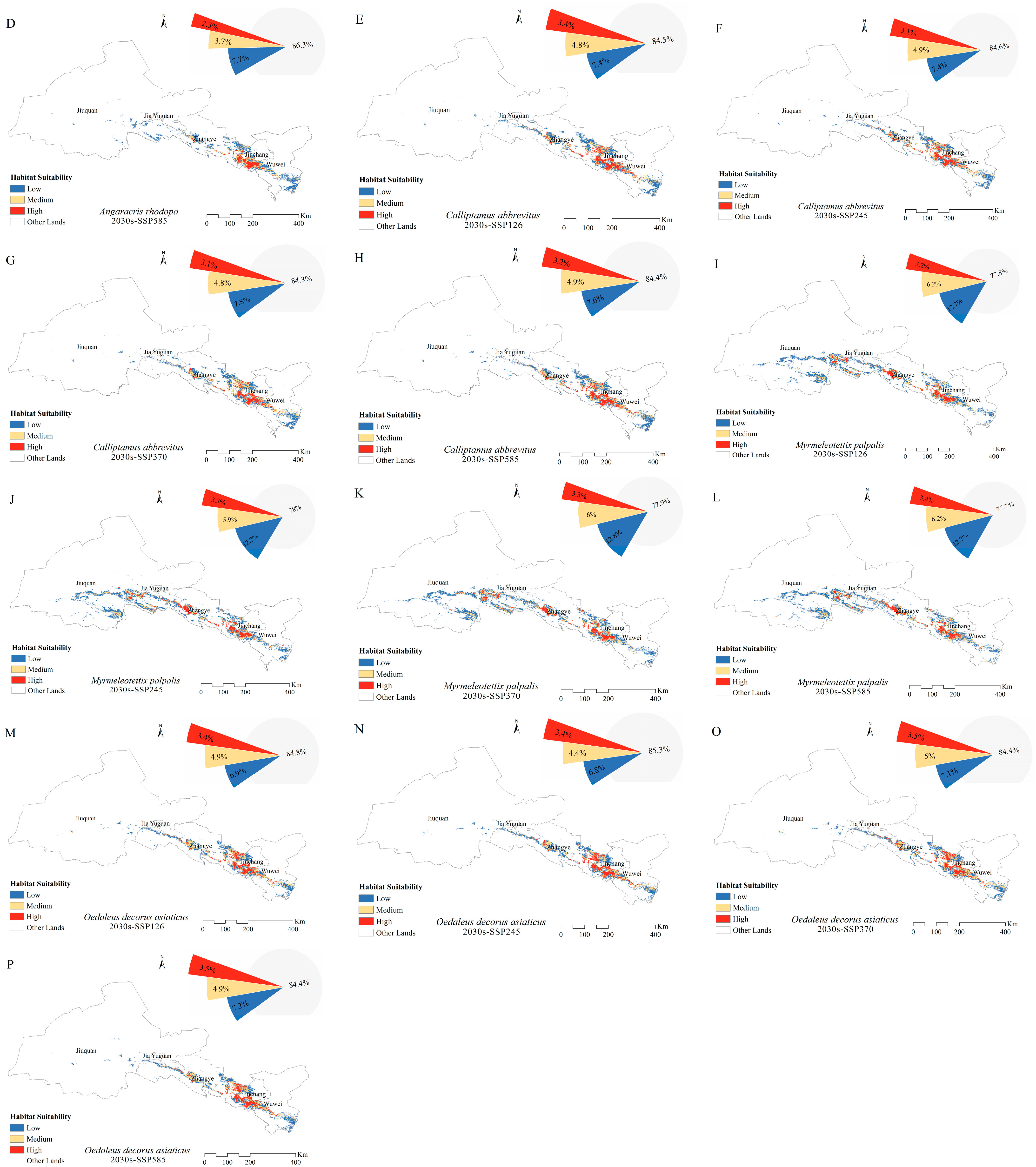
3.4. Potential Distribution of Grasshoppers under Future Climates
4. Discussion
4.1. Selection of Distribution Points and Variables for the MaxEnt Model
4.2. Influence of Environmental Variables on Grasshopper Distribution
4.3. Changes in the Distribution of Grasshopper Habitat Areas under Future Climate Scenarios
4.4. Monitoring and Protection
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, A.; Xu, C.; Zhang, M.; Guo, M.; Xing, X.Y.; Yang, D.; Xu, B.; Yang, X.C. Cross-scale mapping of above-ground biomass and shrub dominance by integrating UAV and satellite data in temperate grassland. Remote Sens. Environ. 2024, 304, 114024. [Google Scholar] [CrossRef]
- Hao, H.W.; Bao, M.; Ke, J.; Li, L.L.; Ma, C.X.; Dan, Z.C.; Chen, Z.N. Fauna elements and eco-geographical distribution of locus in Qinghai province. J. Biol. 2019, 36, 62–68. [Google Scholar] [CrossRef]
- Xin, B.; Dang, Y.Q.; Wang, X.Y.; Yang, Z.Q. Mechanisms and influential factors of southern limits in insect. Acta Ecol. Sin. 2019, 39, 9379–9386. [Google Scholar]
- Shen, Z.H.; Zhao, J. Prediction of spatial patterns of species richness based on plant-topography relationships: An application of GAMs approch. Acta Ecol. Sin. 2007, 27, 953–963. [Google Scholar]
- Wood, S.N. mgcv: GAMs and generalized ridge regression for R. R News 2001, 1, 20–25. [Google Scholar]
- Isley, F.B. The relations of Texas Acrididae to plants and soils. Ecol. Monogr. 1938, 8, 553–604. [Google Scholar] [CrossRef]
- Miao, H.T.; Liu, Y.; Shan, L.Y.; Wu, G.L. Linkages of plant-soil interface habitat and grasshopper occurrence of typical grassland ecosystem. Ecol. Indic. 2018, 90, 324–333. [Google Scholar] [CrossRef]
- VanDyke, K.A.; Latchininsky, A.V.; Schell, S.P. Importance of ecological scale in montane grasshopper (Orthoptera: Acrididae) species structure in similar habitat between differing soil textures and dominant vegetative canopy coverage. J. Orthoptera Res. 2009, 18, 215–223. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, N.; He, B.; Liu, C.Y.; Li, Y.; Zhang, H.Y.; Chen, X.Y.; Lin, H. Construction of a GeogDetector-based model system to indicate the potential occurrence of grasshoppers in Inner Mongolia steppe habitats. Bull. Entomol. Res. 2015, 105, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Zhao, C.Z.; Yin, C.Q.; Wang, D.W.; Zhang, J.X. Grasshopper (Orthoptera: Acrididae) density on natural grasslands in upper reaches of Heihe River, Northwest China in relation with topography: An analysis with generalized additive models (GAM). J. Ecol. 2012, 31, 3121–3126. [Google Scholar] [CrossRef]
- Bernays, E.A.; Gonzalez, N.; Angel, J.; Bright, K.L. Food mixing by generalist grasshoppers: Plant secondary compounds structure the pattern of feeding. J. Insect Behav. 1994, 8, 161–180. [Google Scholar] [CrossRef]
- Branson, D.H. Influence of a large late summer precipitation event on food limitation and grasshopper population dynamics in a northern great plains grassland. Environ. Entomol. 2008, 37, 686–695. [Google Scholar] [CrossRef]
- Wang, B.; Deveson, E.D.; Waters, C.; Spessa, A.; Lawton, D.; Feng, P.Y.; Li, D.L. Future climate change likely to reduce the Australian plague locust (Chortoicetes terminifera) seasonal outbreaks. Sci. Total Environ. 2019, 668, 947–957. [Google Scholar] [CrossRef]
- Saha, A.; Rahman, S.; Alam, S. Modeling current and future potential distributions of desert locust Schistocerca gregaria (Forskål) under climate change scenarios using MaxEnt. J. Asia-Pac. Biodivers. 2021, 14, 399–409. [Google Scholar] [CrossRef]
- Macfadyen, S.; McDonald, G.; Hill, M.P. From species distributions to climate change adaptation: Knowledge gaps in managing invertebrate pests in broad-acre grain crops. Agric. Ecosyst. Environ. 2018, 253, 208–219. [Google Scholar] [CrossRef]
- Ramirez-Cabral, N.Y.Z.; Kumar, L.; Shabani, F. Suitable areas of Phakopsora pachyrhizi, Spodoptera exigua, and their host plant Phaseolus vulgaris are projected to reduce and shift due to climate change. Theor. Appl. Climatol. 2018, 135, 409–424. [Google Scholar] [CrossRef]
- Zhang, X.; Cui, X.; Liang, S. Evolution and optimization of an ecological network in an Arid Region based on MSPA-MCR: A case study of the Hexi Corridor. Sustainability 2024, 16, 1704. [Google Scholar] [CrossRef]
- Sun, T.; Liu, Z.Y.; Qin, L.P.; Long, R.J. Grasshopper (Orthoptera: Acrididae) community composition in the rangeland of the northern slopes of the Qilian Mountains in northwestern China. J. Insect Sci. 2015, 15, 171. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.Y.; Huang, X.B.; Gao, L.J.; Xin, B.Y.; Pang, B.P.; Zhang, Z.H. Feeding features of Oedaleus asiaticus on three species of Poaceae plants in the Inner Mongolia typical steppe. J. Environ. Entomol. 2019, 41, 458–464. [Google Scholar]
- Huang, W.G.; Yu, Z.; Zhang, R.; Wang, L.; Zhang, Y.; Zhu, M.M.; Wei, S.H. Relationship between grasshopper outbreak and plant communitiy in typical grasslands of Yanchi in Ningxia. Pratacultural Sci. 2014, 31, 306–312. [Google Scholar] [CrossRef]
- Fang, Y.C. The occurrence and control situation of grassland locusts in Gansu Province. Pratacultural Sci. 2009, 26, 157–160. [Google Scholar]
- Anderson, R.P.; Raza, A. The effect of the extent of the study region on GIS models of species geographic distributions and estimates of niche evolution: Preliminary tests with montane rodents (genus Nephelomys) in Venezuela. J. Biogeogr. 2010, 37, 1378–1393. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wang, T.L.; Wang, G.Y.; Innes, J.; Nitschke, C.; Kang, H.J. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia–Pacific region. For. Ecol. Manag. 2016, 360, 357–366. [Google Scholar] [CrossRef]
- Leng, W.F.; He, H.S.; Liu, H.J. Response of larch species to climate changes. J. Plant Ecol. 2008, 1, 203–205. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Modell. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Hausser, J.; Chessel, D.; Perrin, N. Ecological-niche factor analysis: How to compute habitat-suitability maps without absence data? Ecology 2002, 83, 2027–2036. [Google Scholar] [CrossRef]
- Busby, J.R. BIOCLIM: A bioclimate analysis and prediction system. Plant Prot. Q. 1991, 6, 8–9. [Google Scholar]
- Zhang, Z.D.; Zang, R.G.; Convertino, M. Predicting the distribution of potential natural vegetation based on species functional groups in fragmented and species-rich forests. Plant Ecol. Evol. 2013, 146, 261–271. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.C.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Bai, D.F.; Chen, P.J.; Atzeni, L.; Cering, L.; Li, Q.; Shi, K. Assessment of habitat suitability of the snow leopard (Panthera uncia) in Qomolangma National Nature Reserve based on MaxEnt modeling. Zool. Res. 2018, 39, 373–386. [Google Scholar] [CrossRef]
- Wu, T.T.; Pan, C.T.; Bian, T.; Wang, Q.X.; Kou, J.; Zhou, B.W. Response of a sylvan moss species (Didymodon validus Limpr.) with a narrow distribution range to climate change. Forests 2023, 14, 2227. [Google Scholar] [CrossRef]
- Hu, J.H.; Liu, Y. Unveiling the conservation biogeography of a data-deficient endangered bird species under climate change. PLoS ONE 2014, 9, e84529. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Change Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]
- Cui, L.N.; Berger, U.; Cao, M.M.; Zhang, Y.Q.; He, J.M.; Pan, L.H.; Jiang, J. Conservation and restoration of mangroves in response to invasion of spartina alterniflora based on the MaxEnt Model: A case study in China. Forests 2023, 14, 1220. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Y.Y.; Wu, W.P.; He, X.Z.; Wang, Z.H.; Guan, Y.Y.; Xu, N.; Chen, Q.L.; Shen, Y.J.; Cao, J.P. Cryptosporidiosis threat under climate change in China: Prediction and validation of habitat suitability and outbreak risk for human-derived Cryptosporidium based on ecological niche models. Infect. Dis. Poverty 2023, 12, 35. [Google Scholar] [CrossRef]
- Cao, B.; Bai, C.K.; Wu, K.Y.; La, T.; Su, Y.Y.; Che, L.Y.; Zhang, M.; Lu, Y.M.; Gao, P.F.; Yang, J.J.; et al. Tracing the future of epidemics: Coincident niche distribution of host animals and disease incidence revealed climate-correlated risk shifts of main zoonotic diseases in China. Glob. Change Biol. 2023, 29, 3723–3746. [Google Scholar] [CrossRef]
- Wang, L.Y.; Hu, W.B.; Soares Magalhaes, R.J.; Bi, P.; Ding, F.; Sun, H.L.; Li, S.L.; Yin, W.W.; Wei, L.; Liu, Q.Y.; et al. The role of environmental factors in the spatial distribution of Japanese encephalitis in mainland China. Environ. Int. 2014, 73, 1–9. [Google Scholar] [CrossRef]
- Guo, J.; Lu, L.H.; Dong, Y.Y.; Huang, W.J.; Zhang, B.; Du, B.B.; Ding, C.; Ye, H.C.; Wang, K.; Huang, Y.R.; et al. Spatiotemporal distribution and main influencing factors of grasshopper potential habitats in two steppe types of inner mongolia, China. Remote Sens. 2023, 15, 866. [Google Scholar] [CrossRef]
- Wen, F.; Lu, L.H.; Nie, C.J.; Sun, Z.X.; Liu, R.H.; Huang, W.J.; Ye, H.C. Analysis of spatiotemporal variation in habitat suitability for Oedaleus decorus asiaticus Bei-Bienko on the Mongolian Plateau using Maxent and multi-source remote sensing data. Insects 2023, 14, 492. [Google Scholar] [CrossRef] [PubMed]
- Du, B.B.; Wei, J.; Lin, K.J.; Lu, L.H.; Ding, X.L.; Ye, H.C.; Huang, W.J.; Wang, N. Spatial and temporal variability of grassland grasshopper habitat suitability and its main influencing factors. Remote Sens. 2022, 14, 3910. [Google Scholar] [CrossRef]
- Zhang, X.W.; Huang, W.J.; Ye, H.C.; Lu, L.H. Study on the identification of habitat suitability areas for the dominant locust species Dasyhippus barbipes in inner Mongolia. Remote Sens. 2023, 15, 1718. [Google Scholar] [CrossRef]
- Lv, Y.Y.; Yu, H.Y.; Chen, W.; Li, M.; Yi, S.H.; Meng, B.P. Predicting inhabitable areas for locust based on field observation and multi-environmental factors in alpine grassland—A case study in the Qilian Mountain National Park, China. Front. Ecol. Evol. 2023, 11, 1149952. [Google Scholar] [CrossRef]
- Li, L.L.; Zhao, C.Z.; Zhao, X.W.; Wang, D.W.; Li, Y. Pattern of plant communities’ influence to grasshopper abundance distribution in heterogeneous landscapes at the upper reaches of Heihe River, Qilian Mountains, China. Environ. Sci. Pollut. Res. Int. 2021, 29, 13177–13187. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Zhao, C.Z.; Yin, C.Q.; Wang, D.W.; Zhang, J.X. Species richness of grasshoppers (Orthoptera: Acrididae) on natural grasslands in relation with topography in the upper reaches of Heihe River, western China analyzed with generalized additive models (GAMs). Acta Entomol. Sin. 2011, 54, 1312–1318. [Google Scholar] [CrossRef]
- Zhou, W.; Zhao, C.Z.; Wang, K.M. Distribution characteristics of grasshoppers in the Black River Basin and their relationship with habitats. J. Arid. Land 2010, 24, 147–152. [Google Scholar] [CrossRef]
- Sun, T.; Long, R.J.; Liu, Z.Y. A comparative study of grasshopper species (Orthoptera: Acridoidea) diversity in different grasslands in the northern slopes of Qilian Mountains. Acta Entomologica Sin. 2010, 53, 702–707. [Google Scholar] [CrossRef]
- Zheng, S.L. Natural grassland resources and their evaluation in Gansu Hexi Corridor. Chin. J. Grassl. 1989, 6, 5–10. [Google Scholar]
- Liu, Y.; Liu, X.; Zhao, C.; Wang, H.; Zang, F. The trade-offs and synergies of the ecological-production-living functions of grassland in the Qilian mountains by ecological priority. J. Environ. Manag. 2023, 327, 116883. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jia, W.X.; Shi, Y.; Zhang, Z.Y.; Xiong, H.; Zhu, G.F. Spatiotemporal differentiation of soil organic carbon of grassland and its relationship with soil physicochemical properties on the northern slope of Qilian Mountains, China. Sustainability 2020, 12, 9396. [Google Scholar] [CrossRef]
- Yang, X.T.; Fan, J.; Ge, J.M.; Du, M.G.; Jin, M. Soil physical and chemical properties and vegetation characteristics of different types of grassland in Qilian Mountains, China. J. Appl. Ecol. 2022, 33, 878–886. (In Chinese) [Google Scholar] [CrossRef]
- Zhao, Z.; He, Y.; Li, Q.; Jia, S.F.; Yang, P.Y.; Jin, C.F.; Yang, X.L.; Sun, X.Y.; Li, J.; Gu, Z.L.; et al. Investigation of grassland resources in Sunan Yugur Autonomous County. Acta Prataculturae Sin. 2010, 19, 231–247. [Google Scholar]
- Zhang, Y.S.; Jia, W.X.; Zhao, Y.F.; Liu, Y.R.; Zhao, Z.; Chen, J.H. Spatio-temporal variations of net primary productivity of Qilian Mountains vegetation based on CASA model. Acta Bot. Boreali-Occident. Sin. 2014, 34, 2085–2091. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, T.; Chen, N.L. Vertical distribution patterns of soil organic carbon and total nitrogen and related affecting factors along northern slope of Qilian Mountains. J. Appl. Ecol. 2009, 20, 518–524. [Google Scholar]
- Wang, S.; Lu, Y.; Han, M.; Li, L.; He, P.; Shi, A.; Bai, M. Using MaxEnt Model to Predict the Potential Distribution of Three Potentially Invasive Scarab Beetles in China. Insects 2023, 14, 239. [Google Scholar] [CrossRef]
- Dong, R.; Hua, L.M.; Hua, R.; Ye, G.H.; Bao, D.; Cai, X.C.; Cai, B.; Zhao, X.C.; Chu, B.; Tang, Z.S. Prediction of the potentially suitable areas of Ligularia virgaurea and Ligularia sagitta on the Qinghai–Tibet Plateau based on future climate change using the MaxEnt model. Front. Plant Sci. 2023, 14, 1193690. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Li, X.; Wen, Y.; Huang, J.; Du, P.; Su, W.; Geng, M. A global record of annual terrestrial Human Footprint dataset from 2000 to 2018. Sci. Data 2022, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, Y.; Jiang, X.; Chen, H.; Liu, M.; Wang, R. Potential geographical distribution of the edangred plant isoetes under human activities using MaxEnt and GARP. Glob. Ecol. Conserv. 2022, 38, e02186. [Google Scholar] [CrossRef]
- Naudiyal, N.; Wang, J.N.; Ning, W.; Gaire, N.P.; Shi, P.L.; Wei, Y.Q.; He, J.L.; Shi, N. Potential distribution of Abies, Picea, and Juniperus species in the sub-alpine forest of Minjiang headwater region under current and future climate scenarios and its implications on ecosystem services supply. Ecol. Indic. 2021, 121, 107131. [Google Scholar] [CrossRef]
- Guo, Y.L.; Wei, H.Y.; Lu, C.Y.; Gao, B.; Gu, W. Predictions of potential geographical distribution and quality of schisandra sphenanthera under climate change. PeerJ 2016, 4, e2554. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Fang, Y.Q.; Zhang, X.H.; Wei, H.Y.; Wang, D.J.; Chen, R.D.; Wang, L.K.; Gu, W. Predicting the invasive trend ofexotic plants in China based on the ensemble model under climate change: A case for three invasive plants of asteraceae. Sci. Total Environ. 2021, 756, 143841. [Google Scholar] [CrossRef]
- Yan, X.L. Mountainous grassland ecological system in Hexi corridor. Pratacultural Sci. 2013, 7, 853–858. [Google Scholar]
- Han, L.; Zhang, Z.; Zhang, Q.; Wan, X. Desertification assessments in the Hexi corridor of Northern China’s Gansu Province by remote sensing. Nat. Hazards 2015, 75, 2715–2731. [Google Scholar] [CrossRef]
- Meng, J.J.; Li, Z.G.; Wu, X.Q. Land use changes of Hexi Corridor between 1995 and 2000. J. Nat. Resour. 2003, 18, 645–651. [Google Scholar] [CrossRef]
- Tian, H.D.; Stige, L.C.; Cazelles, B.; Kausrud, K.L.; Svarverud, R.; Stenseth, N.C.; Zhang, Z.B. Reconstruction of a 1,910-y-long locust series reveals consistent associations with climate fluctuations in China. Proc. Natl. Acad. Sci. USA 2011, 108, 14521–14526. [Google Scholar] [CrossRef]
- Ma, S.C. Study on long-term prediction of locust population fluctuations. Acta Entomol. Sin. 1965, 14, 319–338. [Google Scholar]
- Yamashita, O.; Yaginuma, T.; Hasegawa, K. Hormonal and metabolic control of egg diapause of the silkworm, Bombyx mori (Lepidoptera: Bombycidae). Entomol. Gen. 1981, 7, 195–211. [Google Scholar] [CrossRef]
- Knop, E.; Schmid, B.; Herzog, F. Impact of regional species pool on grasshopper restoration in hay meadows. Restor. Ecol. 2008, 16, 34–38. [Google Scholar] [CrossRef]
- Yu, F. Vegetation Growth Based on MODIS and the Relationship between Vegetation and Grasshoppers of Grasshoppers Plague Areas in Altay; Xinjiang Normal University: Ürümqi, China, 2008. [Google Scholar]
- Cao, K.L.; Wang, Y.; Gao, Y.F.; Tan, S.Q.; Shi, W.P. Regulatory effects of vegetation on the behavior and population of grasshoppers. J. Plant Prot. 2021, 48, 54–59. [Google Scholar] [CrossRef]
- Yang, Y.H.; Rao, S.; Hu, H.F.; Chen, A.P.; Ji, C.J.; Zhu, B.; Zuo, W.Y.; Li, X.R.; Shen, H.H.; Wang, Z.H.; et al. Plant species richness of alpine grasslands in relation to environmental factors and biomass on the Tibetan Plateau. Biodiversity 2004, 12, 200–205. [Google Scholar] [CrossRef]
- Zhao, H.T.; Liu, T.; Lei, J.Q.; Gui, D.E.; Zhao, X.Q. β diversity characteristic of vegetation community on south part of Gurbantunggut Desert and its interpretation. Acta Prataculturae Sin. 2010, 19, 29–37. [Google Scholar]
- Austin, M.P. Searching for a model for use in vegetation analysis. Vegetatio 1980, 42, 11–21. [Google Scholar] [CrossRef]
- Wu, T.J.; Hao, S.G.; Kang, L. Effects of soil temperature and moisture on the development and survival of grasshopper eggs in inner mongolian grasslands. Front. Ecol. Evol. 2021, 9, 727911. [Google Scholar] [CrossRef]
- Shi, R.X.; Liu, G.; Li, D.M.; Xie, B.Y. Distribution of Locusts migratoria and soil in the locust plague area at Baiyangdian. J. Appl. Entomol. 2004, 41, 29–33. [Google Scholar]
- Zhong, Z.W.; Wang, D.L.; Zhu, H.; Wang, L.; Feng, C.; Wang, Z.N. Positive interactions between large herbivores and grasshoppers, and their consequences for grassland plant diversity. Ecology 2014, 95, 1055–1064. [Google Scholar] [CrossRef]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism: An update. Adv. Insect Physiol. 2009, 36, 1–272. [Google Scholar] [CrossRef]
- Hao, S.; Wang, S.; Cease, A.; Knag, L. Landscape level patterns of grasshopper communities in Inner Mongolia: Interactive effects of livestock grazing and a precipitation gradient. Landsc. Ecol. 2015, 30, 1657–1668. [Google Scholar] [CrossRef]
- Cease, A.J.; Elser, J.J.; Ford, C.F.; Hao, S.G.; Kang, L.; Harrison, J.F. Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content. Science 2012, 335, 467–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hunter, D.M. Management of locusts and grasshoppers in China. J. Orthoptera Res. 2017, 26, 155–159. [Google Scholar] [CrossRef]
- Tanaka, H. Embryonic diapause and life cycle in the migratory locust, Locusta migratoria L. (Orthoptera: Acrididae), in Kyoto. Appl. Entomol. Zool. 1994, 29, 179–191. [Google Scholar] [CrossRef]
- Meynard, C.N.; Gay, P.E.; Lecoq, M.; Foucart, A.; Piou, C.; Chapuis, M.P. Climate-driven geographic distribution of the desert locust during recession periods: Subspecies’ niche differentiation and relative risks under scenarios of climate change. Glob. Change Biol. 2017, 23, 4739–4749. [Google Scholar] [CrossRef] [PubMed]
- Josie, H. Modeling the Effect of Landscape Pattern on Mountain Pine Beetles; Simon Fraser University: Burnaby, BC, Canada, 2002. [Google Scholar]
- Vanhanen, H.; Veteli, T.O.; Paivinen, S.; Kellomaki, S.; Niemela, P. Climate change and range shifts in two insect defoliators: Gypsy moth and nun moth-a model study. Silva Fenn. 2007, 41, 621. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Appelberg, G.; Harding, S.; Bärring, L. Spatio-temporal impact of climate change on the activity and voltinism of the spruce bark beetle, Ips typographus. Glob. Change Biol. 2009, 15, 486–499. [Google Scholar] [CrossRef]
- Hof, A.R.; Svahlin, A. The potential effect of climate change on the geographical distribution of insect pest species in the Swedish boreal forest. Scand. J. For. Res. 2016, 31, 29–39. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Code | Variable Name |
|---|---|---|
| Climatic | Bio1 | Annual mean temperature |
| Bio2 | Mean diurnal range (monthly mean (max temp minus min temp)) | |
| Bio3 | Isother mality (BIO2/BIO7) (×100) | |
| Bio4 | Temperature seasonality (standard deviation × 100) | |
| Bio5 | Max temperature of warmest month | |
| Bio6 | Min temperature of coldest month | |
| Bio7 | Temperature annual range (BIO5 minus BIO6) | |
| Bio8 | Mean temperature of wettest quarter | |
| Bio9 | Mean temperature of driest quarter | |
| Bio10 | Mean temperature of warmest quarter | |
| Bio11 | Mean temperature of coldest quarter | |
| Bio12 | Annual precipitation | |
| Bio13 | Precipitation of wettest month | |
| Bio14 | Precipitation of driest month | |
| Bio15 | Precipitation seasonality (coefficient of variation) | |
| Bio16 | Precipitation of wettest quarter | |
| Bio17 | Precipitation of driest quarter | |
| Bio18 | Precipitation of warmest quarter | |
| Bio19 | Precipitation of coldest quarter | |
| LST | Land surface temperature | |
| Vegetation | NDVI | Normalized difference vegetation index |
| GT | Grassland type | |
| Topographical | Elevation | Elevation |
| Slop | Slop | |
| Aspect | Aspect | |
| Soil | AWC | Soil available water content |
| ST | Soil type | |
| PH | T_PH | |
| Human activity | HFP | Human Footprint Index |
| Time | Emission Scenarios | A. rhodopa | C. abbreviatus | M. palpalis | O. decorus asiaticus | ||||
|---|---|---|---|---|---|---|---|---|---|
| Training AUC | Test AUC | Training AUC | Test AUC | Training AUC | Test AUC | Training AUC | Test AUC | ||
| Current | 0.972 | 0.958 | 0.970 | 0.949 | 0.954 | 0.929 | 0.971 | 0.946 | |
| 2021–2040 | SSP126 | 0.972 | 0.953 | 0.971 | 0.944 | 0.955 | 0.932 | 0.973 | 0.947 |
| SSP245 | 0.973 | 0.955 | 0.967 | 0.942 | 0.956 | 0.933 | 0.974 | 0.950 | |
| SSP370 | 0.972 | 0.952 | 0.972 | 0.943 | 0.956 | 0.931 | 0.974 | 0.951 | |
| SSP585 | 0.972 | 0.954 | 0.970 | 0.946 | 0.956 | 0.929 | 0.972 | 0.946 | |
| A. rhodopa | C. abbreviatus | M. palpalis | O. decorus asiaticus | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variables | Percent Contribution (%) | Cumulative Contribution Rate (%) | Variables | Percent Contribution (%) | Cumulative Contribution Rate (%) | Variables | Percent Contribution (%) | Cumulative Contribution Rate (%) | Variables | Percent Contribution (%) | Cumulative Contribution Rate (%) |
| Bio12 | 35.5 | 35.5 | Bio12 | 33.5 | 33.5 | NDVI | 31.8 | 31.8 | Bio12 | 33.6 | 33.6 |
| NDVI | 30.3 | 65.8 | HFP | 19.9 | 53.4 | HFP | 12.4 | 44.2 | AWC | 18.8 | 52.4 |
| Bio7 | 8.5 | 74.3 | AWC | 15.9 | 69.3 | Bio12 | 9.8 | 54 | NDVI | 18.6 | 71 |
| HFP | 5.8 | 80.1 | NDVI | 9.7 | 79 | Bio1 | 9.5 | 63.5 | Bio7 | 9.3 | 80.3 |
| AWC | 5.4 | 85.5 | Bio7 | 7.3 | 86.3 | Bio2 | 8.2 | 71.7 | HFP | 8.9 | 89.2 |
| Bio15 | 4.1 | 89.6 | Slop | 4.8 | 91.1 | Bio7 | 8.1 | 79.8 | Slop | 3.8 | 93 |
| Slop | 2.8 | 92.4 | Bio3 | 2.7 | 93.8 | Slop | 5.9 | 85.7 | Bio15 | 2.8 | 95.8 |
| LST | 2.5 | 94.9 | Bio15 | 2 | 95.8 | Bio19 | 5.6 | 91.3 | Elevation | 1.5 | 97.3 |
| Bio2 | 1.9 | 96.8 | Elevation | 1.3 | 97.1 | GT | 2.9 | 94.2 | Bio2 | 0.9 | 98.2 |
| Aspect | 1.1 | 97.9 | Bio1 | 0.9 | 98 | Aspect | 1.8 | 96 | Bio1 | 0.8 | 99 |
| Bio1 | 0.7 | 98.6 | Aspect | 0.7 | 98.7 | AWC | 1.4 | 97.4 | Aspect | 0.4 | 99.4 |
| GT | 0.5 | 99.1 | GT | 0.5 | 99.2 | ST | 0.9 | 98.3 | GT | 0.2 | 99.6 |
| Bio19 | 0.4 | 99.5 | Bio2 | 0.3 | 99.5 | LST | 0.6 | 98.9 | Bio19 | 0.2 | 99.8 |
| Bio4 | 0.2 | 99.7 | ST | 0.2 | 99.7 | Elevation | 0.6 | 99.5 | ST | 0.1 | 99.9 |
| ST | 0.1 | 99.8 | PH | 0.1 | 99.8 | Bio14 | 0.3 | 99.8 | PH | 0.1 | 100 |
| Elevation | 0.1 | 99.9 | LST | 0.1 | 99.9 | PH | 0.1 | 99.9 | LST | 0 | 100 |
| PH | 0.1 | 100 | Bio4 | 0.1 | 100 | Bio4 | 0.1 | 100 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Gan, H.; Li, X.; Zhou, H.; Zhang, H.; Liu, Y.; Dong, R.; Hua, L.; Hu, G. Changes in the Range of Four Advantageous Grasshopper Habitats in the Hexi Corridor under Future Climate Conditions. Insects 2024, 15, 243. https://doi.org/10.3390/insects15040243
Li D, Gan H, Li X, Zhou H, Zhang H, Liu Y, Dong R, Hua L, Hu G. Changes in the Range of Four Advantageous Grasshopper Habitats in the Hexi Corridor under Future Climate Conditions. Insects. 2024; 15(4):243. https://doi.org/10.3390/insects15040243
Chicago/Turabian StyleLi, Donghong, Huilin Gan, Xiaopeng Li, Huili Zhou, Hang Zhang, Yaomeng Liu, Rui Dong, Limin Hua, and Guixin Hu. 2024. "Changes in the Range of Four Advantageous Grasshopper Habitats in the Hexi Corridor under Future Climate Conditions" Insects 15, no. 4: 243. https://doi.org/10.3390/insects15040243