
Biodiversity and Spatiotemporal Variations of Mecoptera in Thailand: Influences of Elevation and Climatic Factors

 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
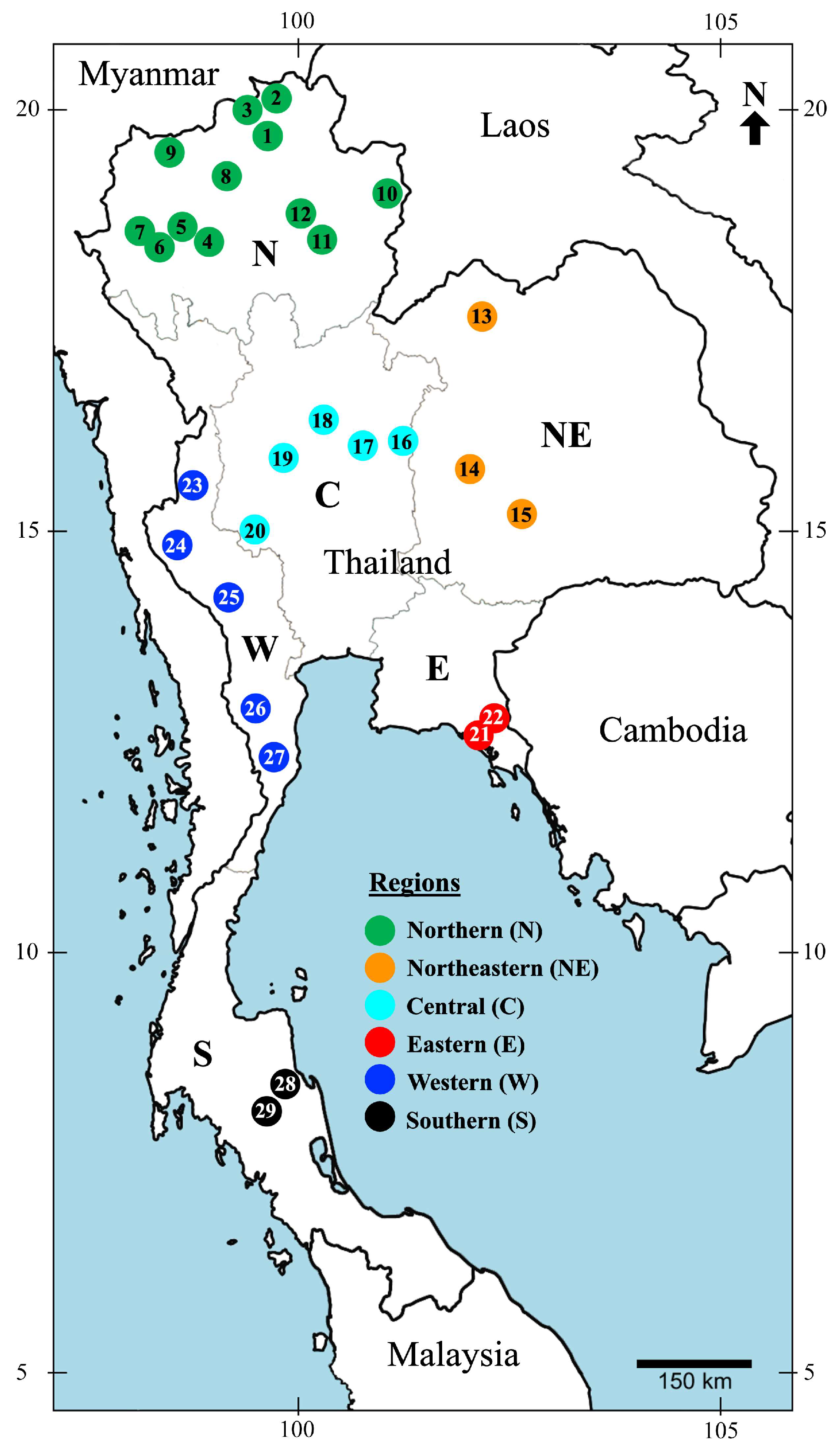
2.1. Study Sites
2.2. Collection Methods
2.3. Species Identification
2.4. Climatic Variables
2.5. Statistical Analyses
3. Results
3.1. Biodiversity and Community Structure of Mecoptera in Thailand
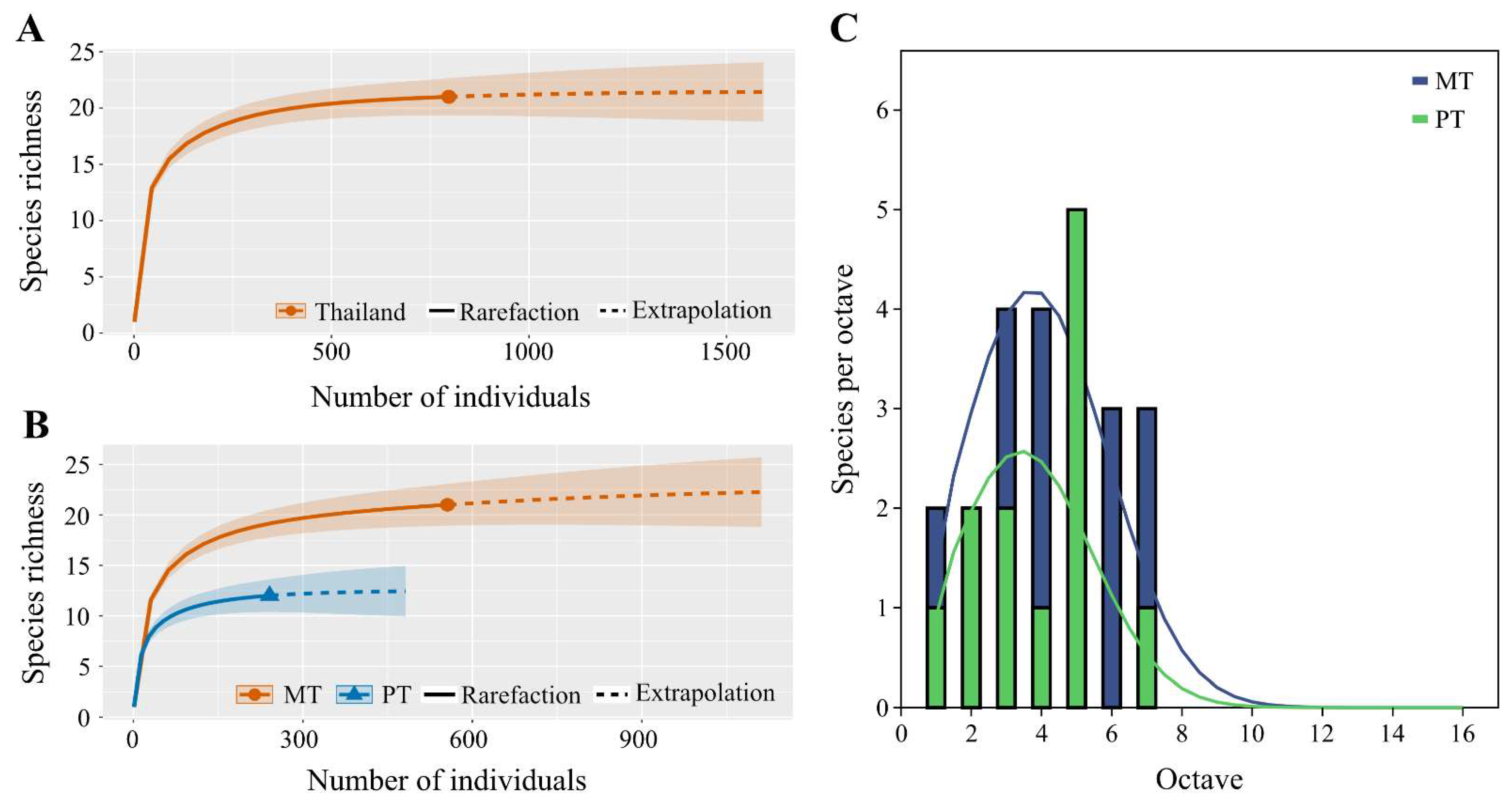
3.1.1. Biodiversity of Mecoptera in Thailand
3.1.2. Community Structure of Mecoptera in Thailand
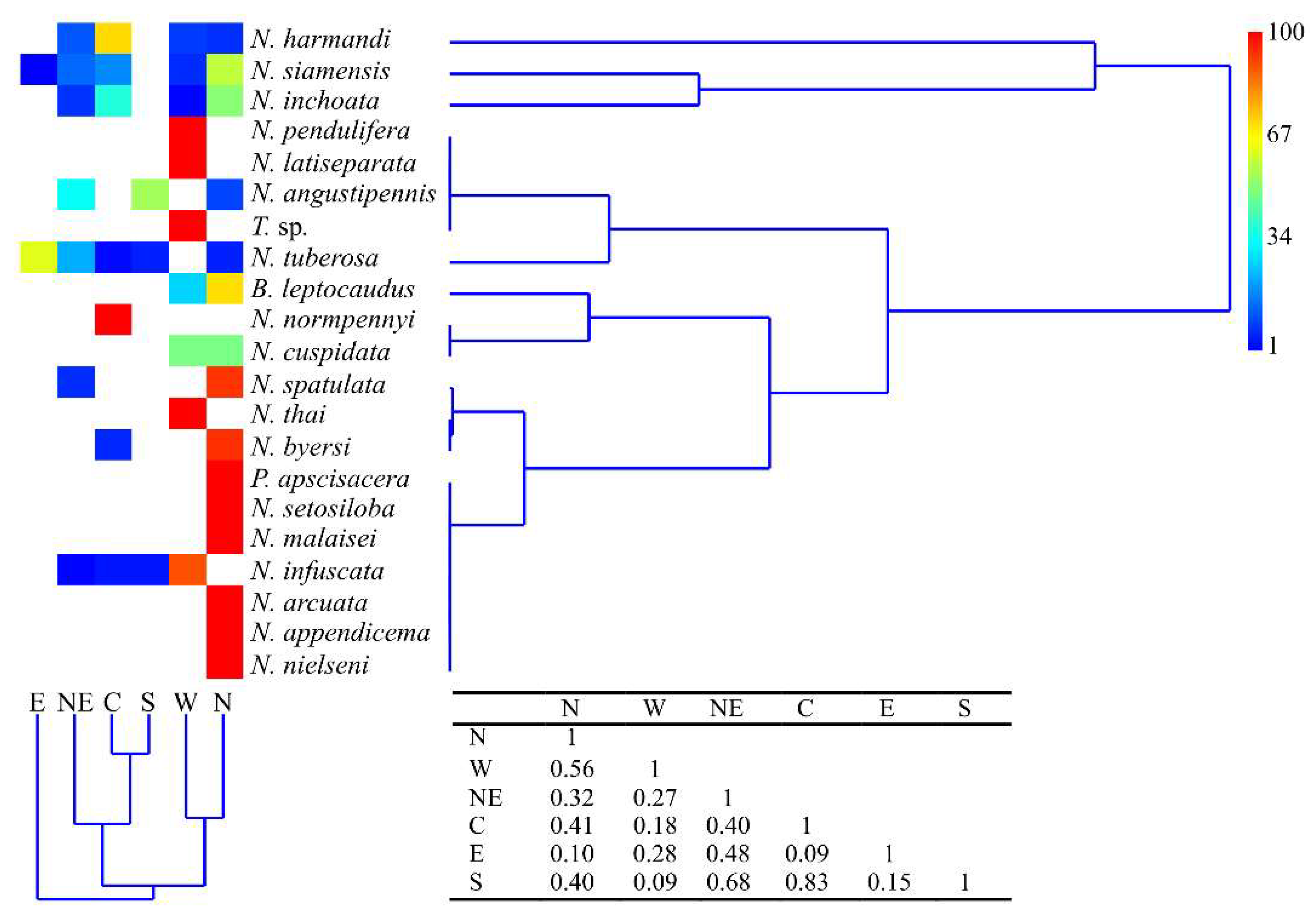
3.1.3. Regional Distribution of Mecoptera in Thailand
3.2. Spatiotemporal Variation of Mecoptera in Thailand
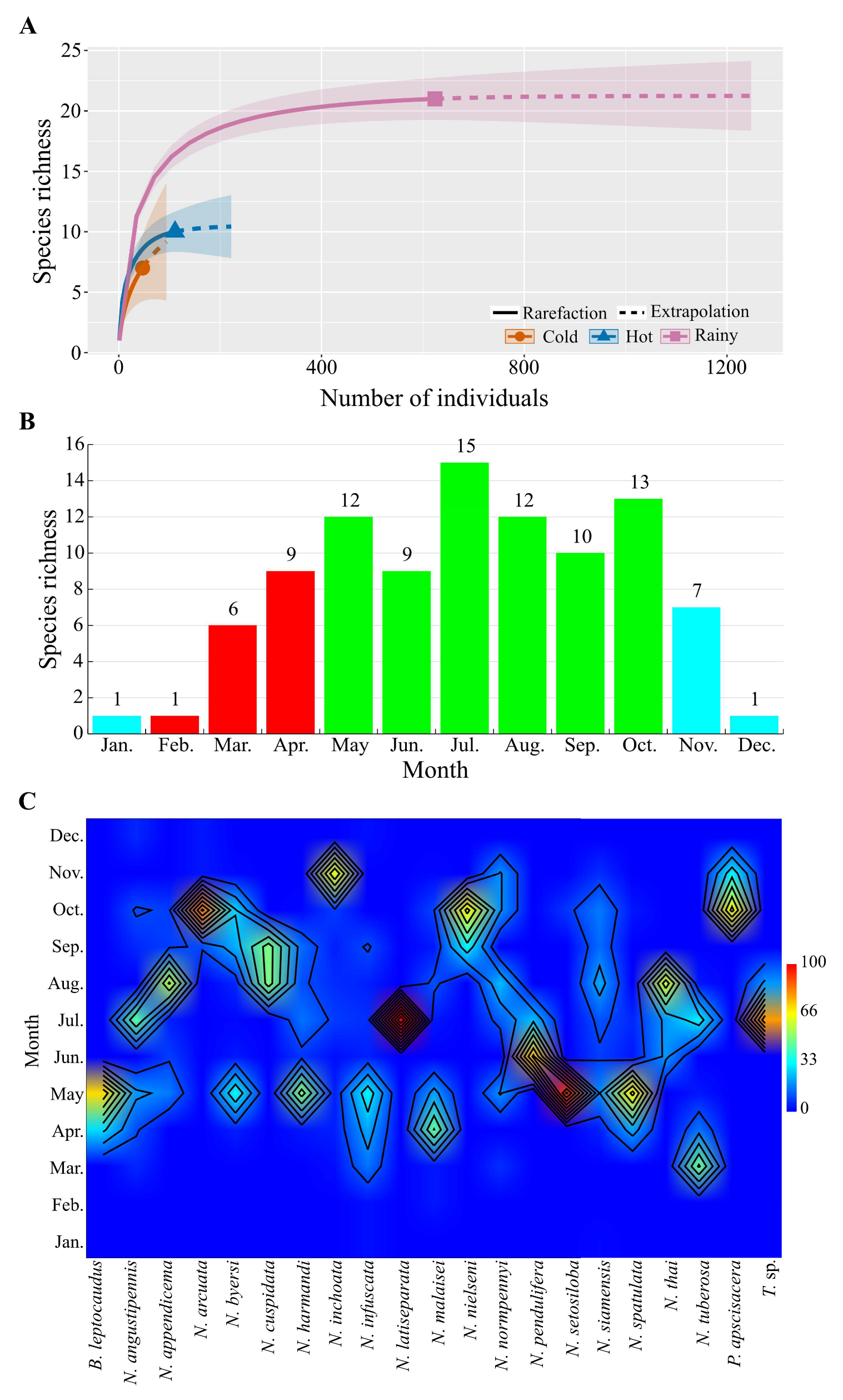
3.2.1. Seasonal Species Richness and Monthly Variation or Emergence Time
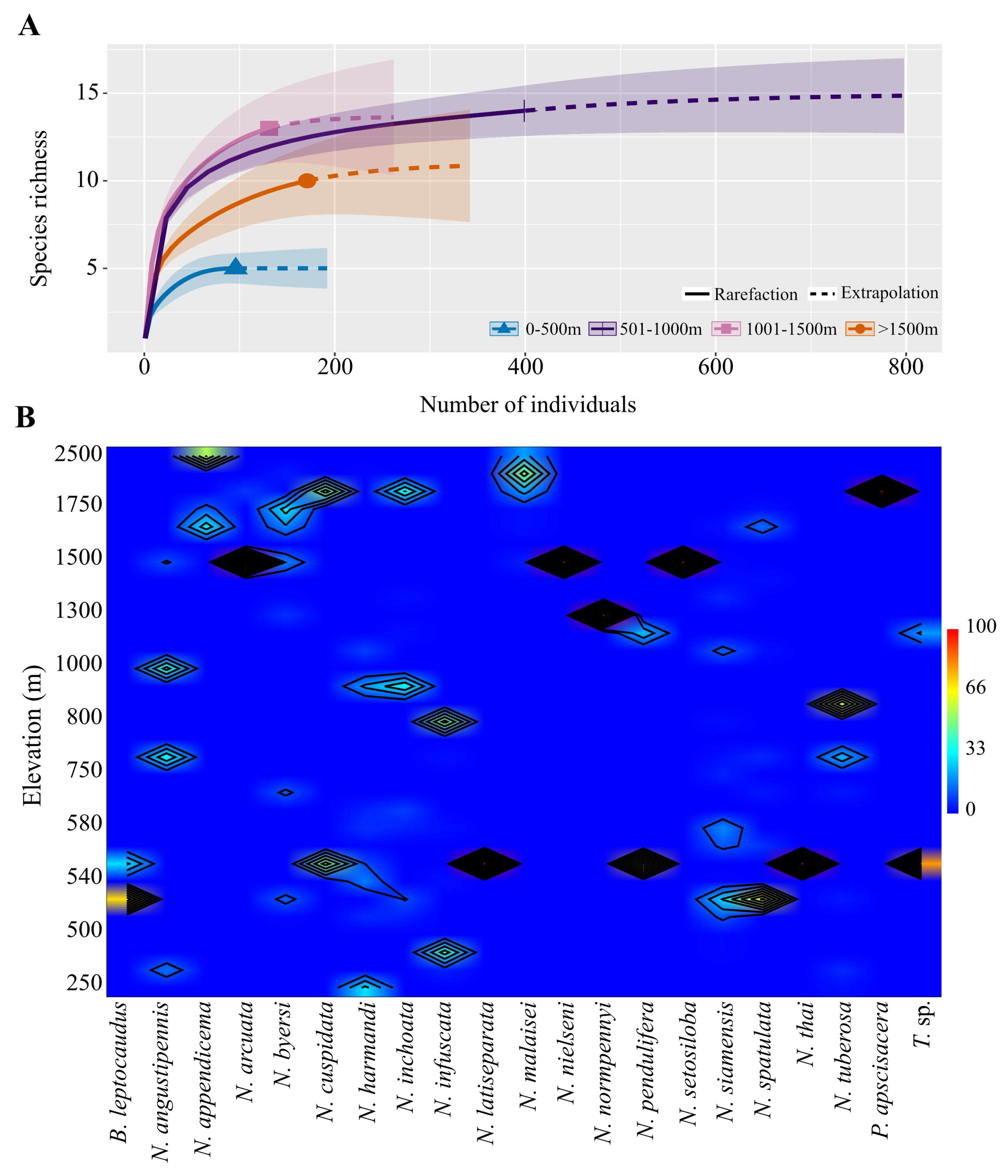
3.2.2. Elevational Species Richness and Abundance
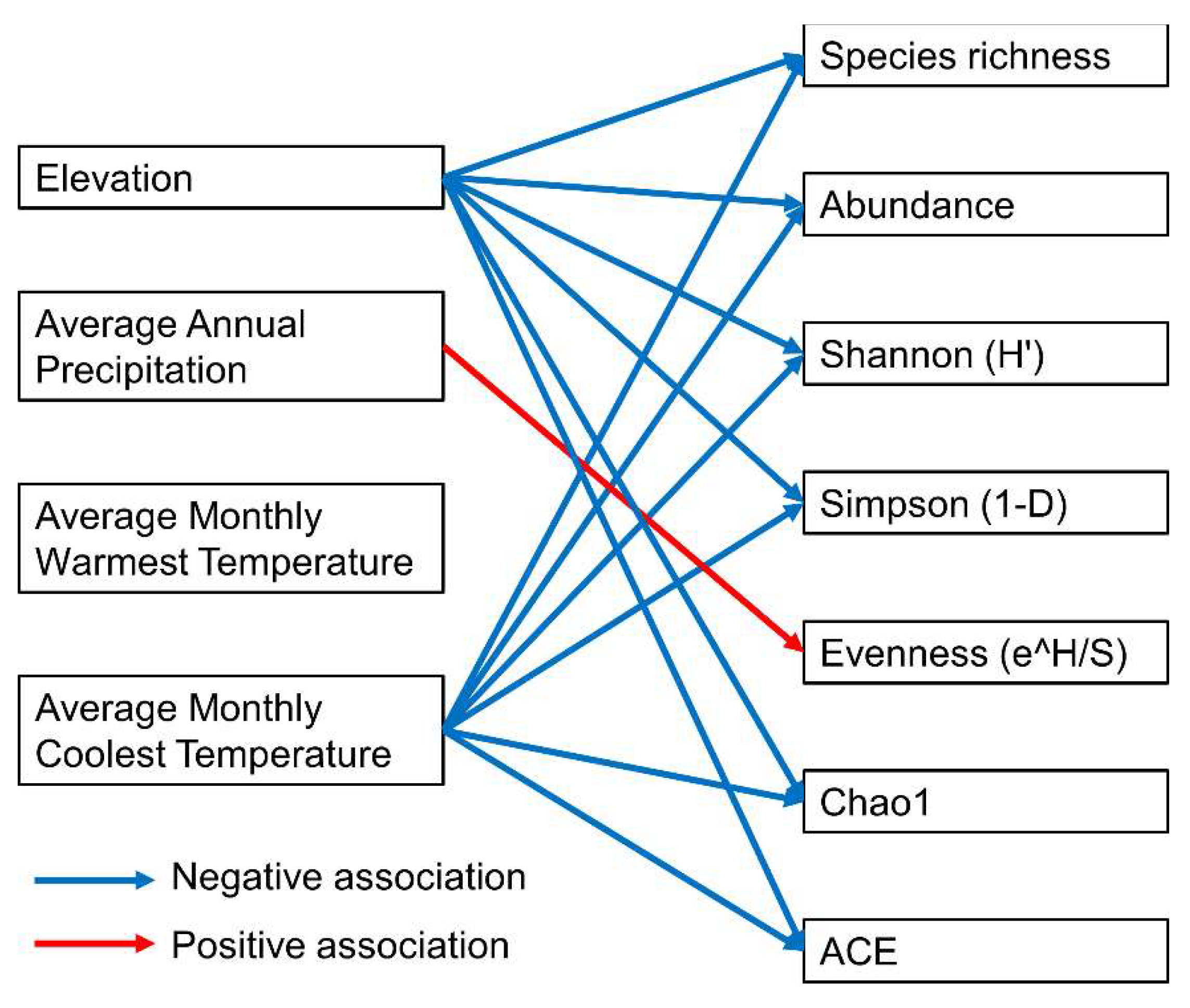
3.3. Impact of Environmental Variables on Mecopteran Species Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bicha, J.W. The scorpionflies (Mecoptera) of Indochina with the description of new species of Bittacus and Neopanorpa. Proc. Entomol. Soc. Wash. 2015, 117, 435–451. [Google Scholar] [CrossRef]
- Bicha, J.W. Biodiversity of Mecoptera. In Insect Biodiversity: Science and Society, 2nd ed.; Foottit, R.G., Adler, P.H., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2018; Volume 2, pp. 705–720. [Google Scholar]
- Bicha, J.W.; Schiff, N.; Lancaster, A.; Suttiprapan, P.; Dokjan, T.; Scheffler, B. A key to species of the scorpionflies of Thailand with the description of new species of Panorpa and Neopanorpa (Mecoptera: Panorpidae) based on morphology and DNA barcodes. Trans. Am. Entomol. Soc. 2022, 146, 201–228. [Google Scholar] [CrossRef]
- Penny, N.D.; Byers, G.W. A check-list of the Mecoptera of the world. Acta Amazon. 1979, 9, 365–388. [Google Scholar] [CrossRef]
- Bicha, J.W. Scorpionflies (Mecoptera: Panorpidae) collected during Project Tiger with the description of three new species from Thailand. Pan-Pac. Entomol. 2019, 95, 49–63. [Google Scholar] [CrossRef]
- Wang, J.S.; Hua, B.Z. Morphological phylogeny of Panorpidae (Mecoptera: Panorpoidea). Syst. Entomol. 2021, 46, 526–557. [Google Scholar] [CrossRef]
- Whiting, M.F. Mecoptera is paraphyletic: Multiple genes and phylogeny of Mecoptera and Siphonaptera. Zool. Scr. 2002, 31, 93–104. [Google Scholar] [CrossRef]
- Pechal, J.L.; Benbow, M.E.; Jeffery, K. Merope tuber Newman (Mecoptera: Meropeidae) collected in association with carrion in Greene County, Ohio, USA: An infrequent collection of an elusive species. Am. Midl. Nat. 2011, 166, 453–457. [Google Scholar] [CrossRef]
- Wang, M.; Hua, B.Z. High species diversity of the genus Neopanorpa (Mecoptera: Panorpidae) in Yunnan Province, China. Zootaxa 2018, 4483, 36–66. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Liu, W.; Hu, F.; Miao, P.; Xing, L.; Hua, Y. The distribution pattern and species richness of scorpionflies (Mecoptera: Panorpidae). Insects 2023, 14, 332. [Google Scholar] [CrossRef]
- Lindgren, N.K.; Sisson, M.S.; Archambeault, A.D.; Rahlwes, B.C.; Willett, J.R.; Bucheli, S.R. Four forensic entomology case studies: Records and behavioral observations on seldom reported cadaver fauna with notes on relevant previous occurrences and ecology. J. Med. Entomol. 2015, 52, 143–150. [Google Scholar] [CrossRef]
- Palmer, C.M.; Yeates, D.K. Diet and feeding behavior in adults of the Apteropanorpidae (Mecoptera). J. Insect Behav. 2005, 18, 209–231. [Google Scholar] [CrossRef]
- Wang, J.S.; Hua, B.Z. Megapanorpa, a new genus with a single anal horn in males from Oriental China (Mecoptera: Panorpidae). Entomol. Sci. 2018, 22, 64–79. [Google Scholar] [CrossRef]
- Vincent, A.; Tillier, P.; Vicent-Barbaroux, C.; Bouget, C.; Salle, A. Influence of forest decline on the abundance and diversity of Raphidioptera and Mecoptera species dwelling in oak canopies. Eur. J. Entomol. 2020, 117, 372–379. [Google Scholar] [CrossRef]
- Wang, J.S.; Gong, Y.J. Taxonomy of the Panorpa guttata group (Mecoptera: Panorpidae), with descriptions of fourteen new species from China. Zootaxa 2021, 4981, 241–274. [Google Scholar] [CrossRef]
- Zangl, L.; Glatzhofer, E.; Schmid, R.; Randolf, S.; Koblmüller, S. DNA barcoding of Austrian snow scorpionflies (Mecoptera, Boreidae) reveals potential cryptic diversity in Boreus westwoodi. PeerJ 2021, 9, e11424. [Google Scholar] [CrossRef]
- Dvořák, L.; Ruchin, A.B.; Egorov, L.V.; Aleksanov, V.V.; Alekseev, S.K.; Shulaev, N.V.; Zakharova, E.Y. Distribution of species from the genus Panorpa (Mecoptera, Panorpidae) in European Russia except the Caucasus. Nat. Conserv. Res. 2023, 8, 24–33. [Google Scholar] [CrossRef]
- Byers, G.W.; Thornhill, R. Biology of Mecoptera. Ann. Rev. Entomol. 1983, 28, 203–208. [Google Scholar] [CrossRef]
- Thornhill, R. Scorpionflies as kleptoparasites of web-building spiders. Nature 1975, 258, 709–711. [Google Scholar] [CrossRef]
- Bockwinkel, G.; Sauer, K.P. Panorpa scorpionflies foraging in spider webs kleptoparasitism at low risk. Bull. Br. Arachnol. Soc. 1993, 9, 110–112. [Google Scholar]
- Lian, X.N.; Cai, C.Y.; Huang, D.Y. The earliest known species of Permeca (Insecta, Mecoptera, Permochoristidae) from the late Guadalupian Yinping Formation of China. Palaeoentomology 2022, 5, 395–399. [Google Scholar] [CrossRef]
- Wang, P.Y.; Hua, B.Z. Elevational diversity pattern and allochronic divergence of scorpionflies in the Qinling Mountains. Ecol. Indic. 2022, 134, 108500. [Google Scholar] [CrossRef]
- Zhao, L.; Gao, R.; Liu, J.; Liu, L.; Li, R.; Men, L.; Zhang, Z. Effects of environmental factors on the spatial distribution pattern and diversity of insect communities along altitude gradients in Guandi Mountain, China. Insects 2023, 14, 224. [Google Scholar] [CrossRef] [PubMed]
- Santillán, V.; Quitián, M.; Tinoco, B.A.; Zárate, E.; Schleuning, M.; Böhning-Gaese, K.; Neuschulz, E.L. Spatio-temporal variation in bird assemblages is associated with fluctuations in temperature and precipitation along a tropical elevational gradient. PLoS ONE 2018, 13, e0196179. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.H.; Fraterrigo, J.M.; Harmon-Threatt, A. Bee functional traits and their relationship to pollination services depend on many factors: A meta-regression analysis. Insect Conserv. Divers. 2023, 16, 313–323. [Google Scholar] [CrossRef]
- Peters, M.K.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiken, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef]
- Graham, M.; Round, P. Thailand’s Vanishing Flora and Fauna, 1st ed.; Finance One Public Company Ltd.: Bangkok, Thailand, 1994. [Google Scholar]
- Hutacharern, C.; Tubtim, N.; Dokmai, C. Checklists of Insects and Mites in Thailand, 1st ed.; Department of National Parks, Wildlife and Plant Conservation, Ministry of Natural Resources and Environment: Bangkok, Thailand, 2007. [Google Scholar]
- Yotkham, S.; Suttiprapan, P.; Likhitrakarn, N.; Sulin, C.; Srisuka, W. Biodiversity and spatiotemporal variation of longhorn beetles (Coleoptera: Cerambycidae) in tropical forest of Thailand. Insects 2021, 12, 45. [Google Scholar] [CrossRef]
- Betz, O.; Srisuka, W.; Puthz, V. Elevational gradients of species richness, community structure, and niche occupation of tropical rove beetles (Coleoptera: Staphylinidae: Steninae) across mountain slopes in Northern Thailand. Evol. Ecol. 2020, 34, 193–216. [Google Scholar] [CrossRef]
- Plant, A.; Bickel, D.J.; Chatelain, P.; Hauser, M.; Cesne, M.; Surin, C.; Saokhod, R.; Nama, S.; Soulier-Perkins, A.; Daugeron, C.; et al. Spatiotemporal dynamics of insect diversity in tropical seasonal forests is linked to season and elevation, a case from northern Thailand. Raffles Bull. Zool. 2018, 66, 382–393. [Google Scholar]
- Chatelain, P.; Plant, A.; Soulier-Perkins, A.; Daugeron, C. Diversity increases with elevation: Empidine dance flies (Diptera, Empididae) challenge a predominant pattern. Biotropica 2018, 50, 633–640. [Google Scholar] [CrossRef]
- Srisuka, W.; Sulin, C.; Sommitr, W.; Rattanarithikul, R.; Aupalee, K.; Saeung, A.; Harbach, R.E. Mosquito (Diptera: Culicidae) diversity and community structure in Doi Inthanon National Park, northern Thailand. Insects 2022, 13, 814. [Google Scholar] [CrossRef]
- Climatological Group, Meteorological Development Bureau, Meteorological Department. The Climate of Thailand. 2015. Available online: https://www.tmd.go.th/en/archive/thailand_climate.pdf (accessed on 9 December 2021).
- Webb, D.W.; Penny, N.D. Neopanorpa byersi (Mecoptera: Panorpidae), a new species from Thailand. Pac. Insects 1979, 20, 63–66. [Google Scholar]
- Bicha, J.W. A review of the scorpionflies (Mecoptera) of Indochina with the description of a new species of Neopanorpa from Northern Thailand. Zootaxa 2010, 2480, 61–67. [Google Scholar] [CrossRef]
- Byers, G.W. The Mecoptera of Indochina. Pac. Insects. 1965, 7, 705–748. [Google Scholar]
- Byers, G.W. Thirteen new Panorpidae from northern Burma. Entomol. Scand. 1999, 30, 197–218. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Peet, R.K. The measurement of species diversity. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Manly, B.F.J. Randomization, Bootstrap and Monte Carlo Methods in Biology, 3rd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2018. [Google Scholar]
- Tan, J.L.; Hua, B.Z. Terrobittacus, a new genus of the Chinese Bittacidae (Mecoptera) with descriptions of two new species. J. Nat. Hist. 2009, 43, 2937–2954. [Google Scholar] [CrossRef]
- Du, W.; Hua, B.Z. Two new species of the genus Terrobittacus Tan & Hua, 2009 (Mecoptera: Bittacidae) from southwestern China with a key to species. Eur. J. Taxon. 2017, 294, 1–13. [Google Scholar]
- He, L.L.; Hua, B.Z. Two new species of the genus Terrobittacus Tan & Hua, 2009 (Mecoptera, Bittacidae). ZooKeys 2023, 1148, 29–39. [Google Scholar]
- Byers, G.W. A new species of Neopanorpa (Mecoptera: Panorpidae) from Laos. Pan-Pac. Entomol. 1982, 58, 92–95. [Google Scholar]
- Bicha, J.W.; Schiff, N.; Pham, T.H.; Lancaster, A.; Scheffler, B. New species of Neopanorpa (Mecoptera) from Vietnam, with a key to the species of Mecoptera of Vietnam. Proc. Entomol. Soc. Wash. 2017, 119, 529–544. [Google Scholar] [CrossRef]
- Forests and Forestry in Myanmar: An Overview of Forestry Laws, Policies, and Strategies. Available online: https://afocosec.org/newsroom/news/forestry-news/forests-and-forestry-in-myanmar-an-overview-of-forestry-laws-policies-and-strategies/ (accessed on 18 August 2023).
- Zhu, H. Floristic characteristics and affinities in Lao PDR, with a reference to the biogeography of the Indochina peninsula. PLoS ONE 2017, 12, e0179966. [Google Scholar] [CrossRef]
- Thi Hoan, L. Forest resources and forestry in Vietnam. J. Viet. Env. 2014, 6, 171–177. [Google Scholar]
- Gao, K.; Hua, Y.; Xing, L.X.; Hua, B.Z. Speciation of the cold-adapted scorpionfly Cerapanorpa brevicornis (Mecoptera: Panorpidae) via interglacial refugia. Insect Conserv. Divers. 2022, 15, 114–127. [Google Scholar] [CrossRef]
- Wang, J.S.; Hau, B.Z. An annotated checklist of the Chinese Mecoptera with description of male Panorpa guttata Navás, 1908. Entomotaxonomia 2017, 39, 24–42. [Google Scholar]
- Dvorak, L.; Japoshvili, G. The scorpionflies (Mecoptera) from Lagodekhi Reserve, with some distributional and seasonal activity data. Zootaxa 2018, 4407, 137–140. [Google Scholar] [CrossRef] [PubMed]
- World Checklist of Extant Mecoptera Species. Available online: https://researcharchive.calacademy.org/research/entomology/Entomology_Resources/mecoptera/ (accessed on 25 August 2023).
- Wilkie, K.T.R.; Mertl, A.L.; Traniello, J.F.A. Species diversity and distribution patterns of the ants of Amazonian Ecuador. PLoS ONE 2010, 5, e13146. [Google Scholar]
- Hwang, J.H.; Yim, M.Y.; Kim, S.Y.; Ji, S.J.; Lee, W.H. Sweep sampling comparison of terrestrial insect communities associated with herbaceous stratum in the riparian zone of the Miho River, Korea. Insects 2022, 13, 497. [Google Scholar] [CrossRef] [PubMed]
- Barnard, P.C.; Brooks, S.J.; Stork, N.E. The seasonality and distribution of Neuroptera, Raphidioptera and Mecoptera on oaks in Richmond Park, Surrey, as revealed by insecticide knock-down sampling. J. Nat. Hist. 1986, 20, 1321–1331. [Google Scholar] [CrossRef]
- Duelli, P.; Wermelinger, B.; Moretti, M.; Obrist, M.K. Fire and windthrow in forests: Winners and losers in Neuropterida and Mecoptera. Alp. Entomol. 2019, 3, 39–50. [Google Scholar] [CrossRef]
- Faithfull, M.J.; Majer, J.D.; Postle, A.C. Some notes on the occurrence and seasonality of Austromerope poultoni Killington (Mecoptera) in western Australia. Aust. Ent. Mag. 1985, 12, 57–60. [Google Scholar]
- Mundaca, A.; Rojas-Miranda, K.; Munoz-Concha, D. New insights of seasonal fluctuations and distribution of Notiothauma reedi, MacLachlan, 1877 (Mecoptera: Eomeroidae) a rare living fossil from the Chilean biodiversity hotspot. In Proceedings of the Presented at the Ento23, Cornwall, UK, 5–7 September 2023. [Google Scholar]
- Jara-Soto, E.; Muńoz-Escobar, C.; Jerez, V. Record of Notiothauma reedi Mclachlan 1877 (Mecoptera: Eomeropidae) in vertebrates corpses in concepción commune, Chile. Rev. Chilena Ent. 2007, 33, 35–40. [Google Scholar]
- McCoy, E.D. The distribution of insects along elevational gradients. Oikos 1990, 58, 313–322. [Google Scholar] [CrossRef]
- Uhey, D.; Haubensak, K.; Hofstetter, R. Mid-elevational peaks in diversity of ground-dwelling arthropods with high species turnover on the Colorado Plateau. Environ. Entomol. 2021, 50, 337–347. [Google Scholar] [CrossRef]
- Cancino-López, R.J.; Moreno, C.E.; Contreras-Ramos, A. Diversity of lacewings (Neuroptera) in an altitudinal gradient of the Tacaná Volcano, Southern Mexico. Insects 2022, 13, 652. [Google Scholar] [CrossRef]
- Chen, A.; Li, Z.; Zheng, Y.; Zhan, J.; Yang, B.; Yang, Z. Decreasing species richness with increase in elevation and positive Rapoport effects of Crambidae (Lepidoptera) on Mount Taibai. Insects 2022, 13, 1125. [Google Scholar] [CrossRef]
- Wang, J.S. New and little-known species of the genus Dicerapanorpa from northwestern Yunnan, China (Mecoptera: Panorpidae). Acta Entomol. 2022, 62, 1–13. [Google Scholar] [CrossRef]
- Byers, G.W. Descriptions and distributional records of American Mecoptera. II. J. Kans. Entomol. Soc. 1962, 35, 299–307. [Google Scholar]
- Byers, G.W. Additions to the Mecoptera of Mexico. J. Kans. Entomol. Soc. 2011, 84, 1–11. [Google Scholar] [CrossRef]
- Vanin, S.; Bubacco, L.; Beltramini, M. Seasonal variation of trehalose and glycerol concentrations in winter snow-active insects. CryoLetters 2008, 29, 485–491. [Google Scholar] [PubMed]
- Palmer, C.M.; Siebke, K. Cold hardiness of Apteropanorpa tasmanica Carpenter (Mecoptera: Apteropanorpidae). J. Insect Physiol. 2008, 54, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.L.; Hua, Y.; Hebert, P.D.; Hua, B.Z. Evolutionary history of the scorpionfly Dicerapanorpa magna (Mecoptera, Panorpidae). Zool. Scr. 2019, 48, 93–105. [Google Scholar] [CrossRef]
- Thornhill, R. Competition and coexistence among Panorpa scorpionflies (Mecoptera: Panorpidae). Ecol. Monogr. 1980, 50, 179–197. [Google Scholar] [CrossRef]
- Jiang, L.; Hua, Y.; Hu, G.-L.; Hua, B.-Z. Habitat divergence shapes the morphological diversity of larval insects: Insights from scorpionflies. Sci. Rep. 2019, 9, 12708. [Google Scholar] [CrossRef]
- Wanthathaen, C.; Chiu, C.-I.; Chiu, M.-C.; Leelayouthyotin, S.; Krutmuang, P.; Li, H.-F.; Sripontan, Y. Desiccation tolerance of Termitidae termites in relation to their nest type. Environ. Entomol. 2023, 52, 555–564. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| National Parks | Locality Name | Elevation (m, msl) | B. leptocaudus | N. angustipennis | N. appendicema | N. arcuata | N. byersi | N. cuspidata | N. harmandi | N. inchoata | N. infuscata | N. latiseparata | N. malaisei | N. nielseni | N. normpennyi | N. pendulifera | N. setosiloba | N. siamensis | N. spatulata | N. thai | N. tuberosa | P. apscisacera | T. sp. | No. of Specimens | Species Richness/Sites |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Doi Pha Hom Pok | Mae Fang Hotspring | 569 | 3 | 10 | 4 | 17 | 3 | ||||||||||||||||||
| Doi Pha Luang | 1449 | 2 | 28 | 14 | 3 | 6 | 15 | 1 | 69 | 7 | |||||||||||||||
| Kiewlom | 2112 | 2 | 1 | 1 | 19 | 15 | 3 | 41 | 6 | ||||||||||||||||
| Doi Inthanon | Vachiratharn | 700 | 9 | 2 | 1 | 12 | 3 | ||||||||||||||||||
| Checkpoint 2 | 1639 | 8 | 14 | 1 | 2 | 8 | 33 | 5 | |||||||||||||||||
| Kew Mae Pan | 2200 | 2 | 1 | 35 | 38 | 3 | |||||||||||||||||||
| Summit | 2500 | 14 | 15 | 29 | 2 | ||||||||||||||||||||
| Doi Chiang Dao | Pha Tang substation | 526 | 5 | 10 | 5 | 25 | 38 | 1 | 84 | 6 | |||||||||||||||
| Huai Nam Dang | Headquarters | 1670 | 3 | 25 | 1 | 1 | 30 | 4 | |||||||||||||||||
| Doi Phu Kha | Headquarters | 1374 | 1 | 1 | 4 | 6 | 3 | ||||||||||||||||||
| Chae Son | Waterfall | 507 | 1 | 12 | 2 | 10 | 25 | 4 | |||||||||||||||||
| Doi Lan | 1413 | 1 | 1 | 2 | 2 | ||||||||||||||||||||
| Phu Ruea | Rong Huay Maklaow | 1167 | 18 | 11 | 1 | 30 | 3 | ||||||||||||||||||
| Pa Hin Ngam | Tung Dok Grajeaw | 780 | 9 | 4 | 13 | 2 | |||||||||||||||||||
| Khao Yai | Headquarters | 770 | 7 | 1 | 2 | 3 | 9 | 22 | 5 | ||||||||||||||||
| Nam Nao | Checkpoint | 921 | 43 | 18 | 61 | 2 | |||||||||||||||||||
| Sam Makao forest unit | 528 | 34 | 1 | 1 | 36 | 3 | |||||||||||||||||||
| Khao Kho | Headquarters | 230 | 66 | 1 | 67 | 2 | |||||||||||||||||||
| Thung Salaeng Luang | Gang Wang Nam Yen | 580 | 14 | 2 | 1 | 16 | 33 | 4 | |||||||||||||||||
| Mae Wong | Chong Yen | 1306 | 5 | 16 | 21 | 2 | |||||||||||||||||||
| Khao Khitchakut | Prabaht unit | 107 | 1 | 1 | 1 | ||||||||||||||||||||
| Khao Prabaht peak | 875 | 24 | 24 | 1 | |||||||||||||||||||||
| Umphang | Mae Klong Kee | 1231 | 1 | 1 | 1 | 3 | 3 | ||||||||||||||||||
| Thi Lor Su | 567 | 2 | 1 | 19 | 1 | 3 | 5 | 4 | 35 | 7 | |||||||||||||||
| Khuean Srinagarindra | Tham Nanya | 750 | 3 | 3 | 1 | ||||||||||||||||||||
| Kaeng Krachan | Panern Thung | 790 | 24 | 2 | 26 | 2 | |||||||||||||||||||
| Pa La-U waterfall | 320 | 20 | 1 | 21 | 2 | ||||||||||||||||||||
| Namtok Yong | Protection unit 3 | 372 | 3 | 2 | 2 | 7 | 3 | ||||||||||||||||||
| TV aerial | 952 | 8 | 8 | 1 | |||||||||||||||||||||
| No. of specimens (n = 797) | 7 | 20 | 25 | 30 | 82 | 2 | 219 | 52 | 49 | 1 | 70 | 6 | 16 | 4 | 15 | 90 | 58 | 5 | 38 | 3 | 5 | 797 | |||
| Relative abundance (%RA) | 0.9 | 2.6 | 3.1 | 3.8 | 10.3 | 0.2 | 27.4 | 6.5 | 6.1 | 0.1 | 8.8 | 0.8 | 2.0 | 0.5 | 1.9 | 11.3 | 7.3 | 0.6 | 4.8 | 0.4 | 0.6 | ||||
| No. of positive sites (n = 29) | 2 | 4 | 3 | 2 | 10 | 2 | 10 | 8 | 6 | 1 | 6 | 1 | 1 | 2 | 1 | 15 | 8 | 1 | 6 | 1 | 2 | ||||
| Species occurrence (% SO) | 6.9 | 13.8 | 10.3 | 6.9 | 34.5 | 6.9 | 34.5 | 27.6 | 20.7 | 3.4 | 20.7 | 3.4 | 3.4 | 6.9 | 3.4 | 51.7 | 27.6 | 3.4 | 20.7 | 3.4 | 6.9 | ||||
| Regional distribution * | N, W | N, S, NE | N | N | N, C | N, W | N, C, W, N, E | N, C, NE, W | NE, C, W, S | W | N | N | C | W | N | N, C, NE, E, W | N, NE | W | N, NE, C, E, S | N | W | ||||
| Dependent Variable (Value Range): Formula | Formula | Model R2 | Statistics of Independent Variable | |||
|---|---|---|---|---|---|---|
| Independent a Variable | Slope b | t-Value | p Value | |||
| Species richness (1–7) | Species richness = 19. 8343 −0.0028 × Elevation −0.7836 × AMCT | 0.38 | Intercept | 19. 8343 ± 4.4557 | 4.45 | <0.001 |
| Elevation | −0.0028 ± 0.0009 | −2.92 | <0.01 | |||
| AMCT | −0.7836 ± 0.2023 | −3.87 | <0.001 | |||
| Abundance (1–84) | Abundance = 182.1138 −0.0275 × Elevation −7.1669 × AMCT | 0.20 | Intercept | 182.1138 ± 62.0492 | 2.94 | <0.01 |
| Elevation | −0.0275 ± 0.0132 | −2.09 | <0.05 | |||
| AMCT | −7.1669 ± 2.8178 | −2.54 | <0.05 | |||
| Shannon (H′) | Shannon (H′) = 5.5581 −0.0008 × Elevation −0.2266 × AMCT | 0.37 | Intercept | 5.5581 ± 1.3554 | 4.10 | <0.001 |
| (0–1.578) | Elevation | −0.0008 ± 0.0003 | −2.65 | <0.05 | ||
| AMCT | −0.2266 ± 0.0616 | −3.68 | <0.01 | |||
| Simpson (1-D) | Simpson (1-D) = 2.8059 −0.0003 × Elevation −0.1134 × AMCT | 0.30 | Intercept | 2.8059 ± 0.8437 | 3.33 | <0.01 |
| (0–1) | Elevation | −0.0003 ± 0.0002 | −1.93 | 0.06 | ||
| AMCT | −0.1134 ± 0.0383 | −2.96 | <0.01 | |||
| Evenness (e^H/S) (0.441–1.396) | Evenness (e^H/S) = 0.4599 +0.0002 × AAP | 0.17 | Intercept | 0.4599 ± 0.1611 | 2.86 | <0.01 |
| AAP | 0.0002 ± 0.0001 | 2.35 | <0.05 | |||
| Chao1 (1.000–7.486) | Chao1 = 20.6737 −0.0028 × Elevation −0.8189 × AMCT | 0.38 | Intercept | 20.6737 ± 4.7513 | 4.35 | <0.001 |
| Elevation | −0.0028 ± 0.0010 | −2.74 | <0.05 | |||
| AMCT | −0.8189 ± 0.2158 | −3.80 | <0.001 | |||
| ACE (1.000–8.184) | ACE = 23.0892 −0.0028 × Elevation −0.9252 × AMCT | 0.35 | Intercept | 23.0892 ± 6.1352 | 3.76 | <0.001 |
| Elevation | −0.0028 ± 0.0013 | −2.14 | <0.05 | |||
| AMCT | −0.9252 ± 0.2786 | −3.32 | <0.01 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dokjan, T.; Bicha, W.J.; Suttiprapan, P.; Chuttong, B.; Chiu, C.-I.; Aupalee, K.; Saeung, A.; Sulin, C.; Srisuka, W. Biodiversity and Spatiotemporal Variations of Mecoptera in Thailand: Influences of Elevation and Climatic Factors. Insects 2024, 15, 151. https://doi.org/10.3390/insects15030151
Dokjan T, Bicha WJ, Suttiprapan P, Chuttong B, Chiu C-I, Aupalee K, Saeung A, Sulin C, Srisuka W. Biodiversity and Spatiotemporal Variations of Mecoptera in Thailand: Influences of Elevation and Climatic Factors. Insects. 2024; 15(3):151. https://doi.org/10.3390/insects15030151
Chicago/Turabian StyleDokjan, Theerapan, Wesley J. Bicha, Piyawan Suttiprapan, Bajaree Chuttong, Chun-I. Chiu, Kittipat Aupalee, Atiporn Saeung, Chayanit Sulin, and Wichai Srisuka. 2024. "Biodiversity and Spatiotemporal Variations of Mecoptera in Thailand: Influences of Elevation and Climatic Factors" Insects 15, no. 3: 151. https://doi.org/10.3390/insects15030151