
Sequential and Simultaneous Interactions of Plant Allelochemical Flavone, Bt Toxin Vip3A, and Insecticide Emamectin Benzoate in Spodoptera frugiperda

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Sources
2.2. Preparation of Toxin-Containing Diet
2.3. Bioassay
2.4. Data Analysis
3. Results
3.1. Toxicity of Vip3A, Flavone, and Emamectin against S. frugiperda Neonate Larvae
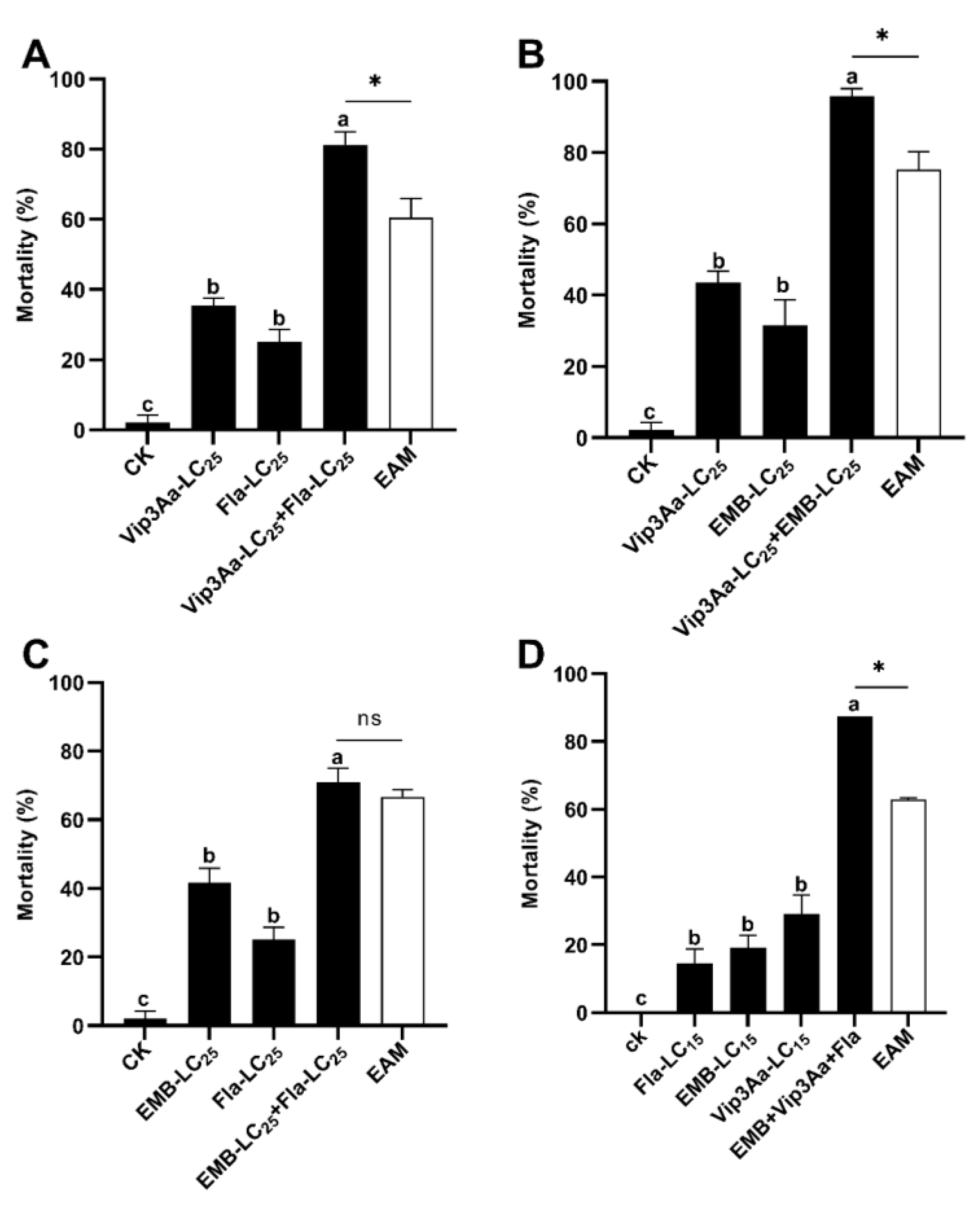
3.2. Toxic Synergy among Vip3A, Emamectin Benzoate, and Flavone against S. frugiperda Neonate Larvae
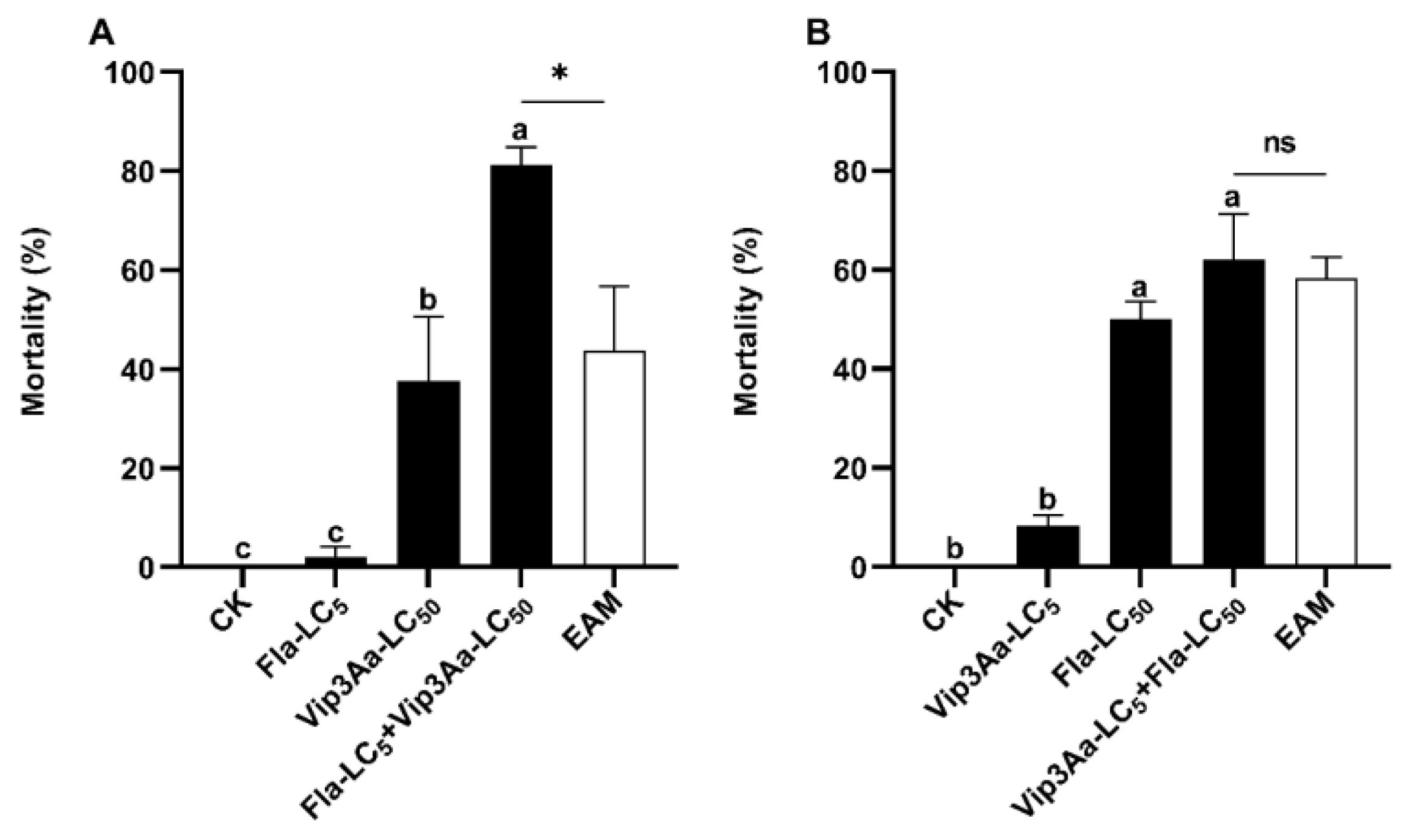
3.3. Asymmetrical Toxicity Induction between Flavone and Vip3A against S. frugiperda Neonates
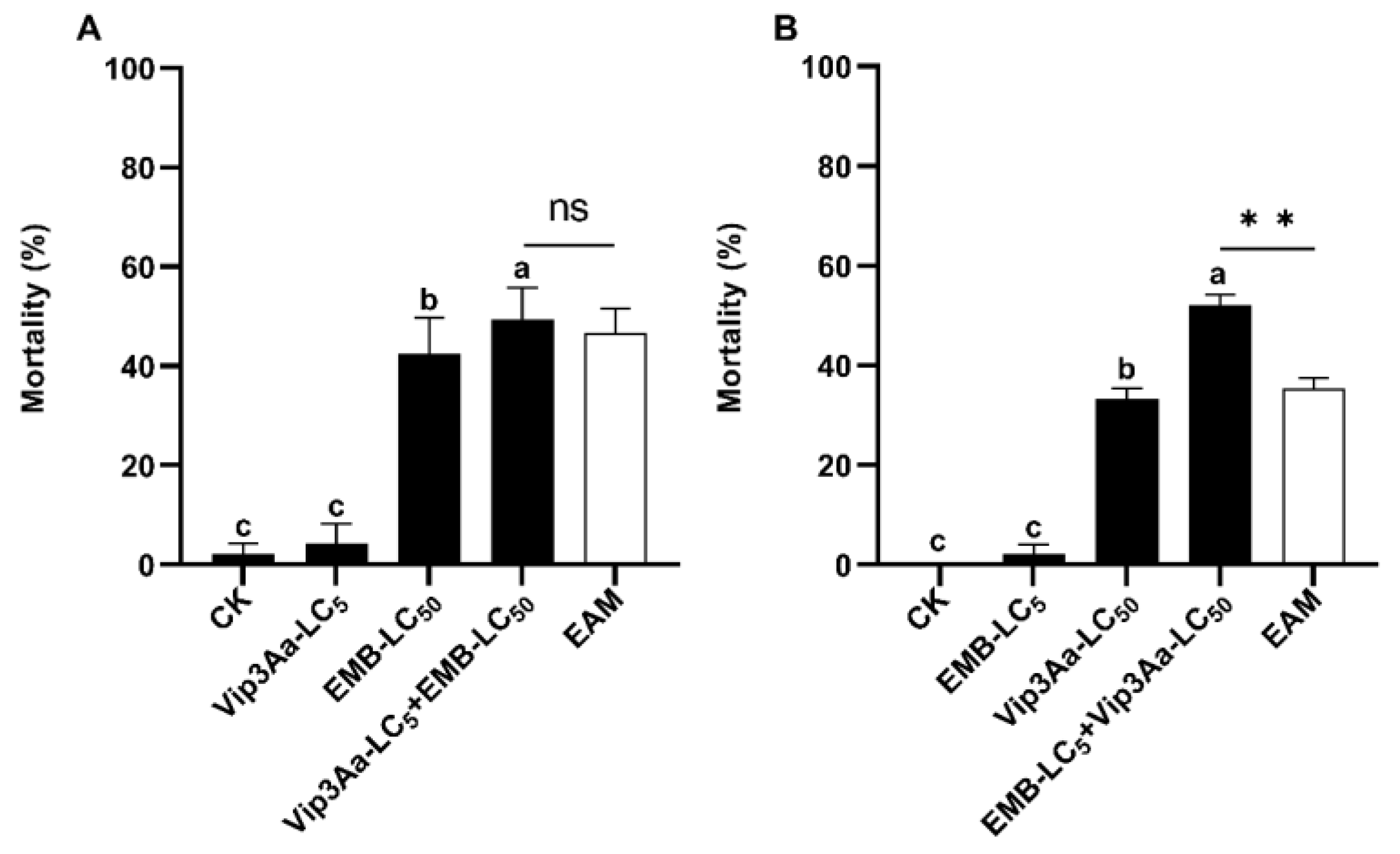
3.4. Asymmetrical Toxicity Induction between Vip3a and Emamectin Benzoate against S. frugiperda Larvae
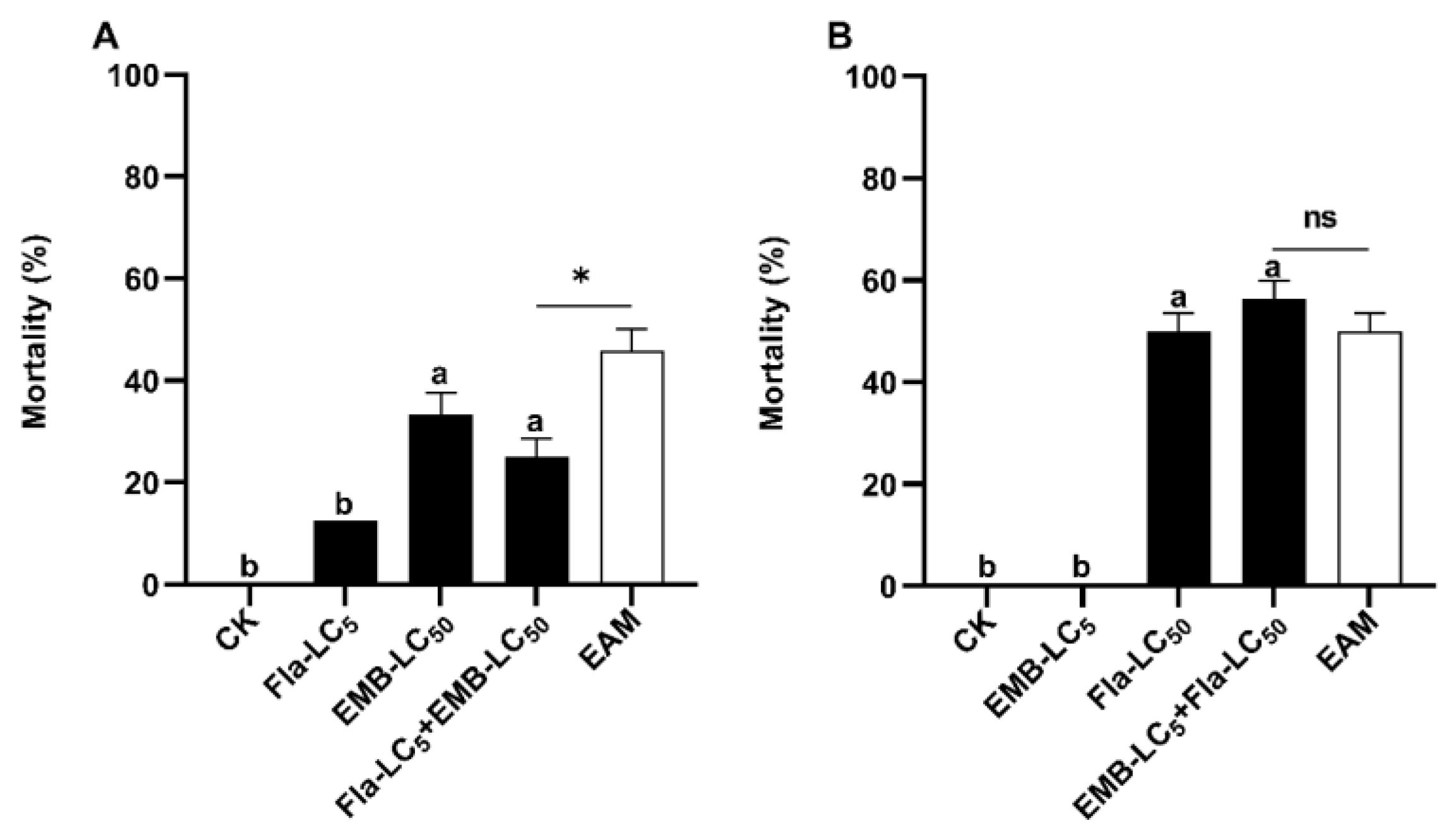
3.5. Asymmetrical Toxicity Inhibition between Flavone and Emamectin Benzoate against S. frugiperda Neonates
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, X.; Ni, X. Deciphering the plant-insect phenotypic arms race. Recent. Adv. Entomol. Res. 2011, 3, 3–33. [Google Scholar]
- Schuman, M.C.; Baldwin, I.T. The layers of plant responses to insect herbivores. Annu. Rev. Entomol. 2016, 61, 373–394. [Google Scholar] [CrossRef] [PubMed]
- Erb, M. Plant defenses against herbivory: Closing the fitness gap. Trends Plant Sci. 2018, 23, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Tanda, A.S. Mutualistic plant associations related to insect resistance. In Molecular Advances in Insect Resistance of Field Crops: Modern and Applied Approaches; Tanda, A.S., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–42. [Google Scholar]
- War, A.R.; Buhroo, A.A.; Hussain, B.; Ahmad, T.; Nair, R.M.; Sharma, H.C. Plant defense and insect adaptation with reference to secondary metabolites. In Co-Evolution of Secondary Metabolites; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 795–822. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.; Brennan, R.M.; Graham, J.; Karley, A.J. Plant Defense against herbivorous pests: Exploiting resistance and tolerance traits for sustainable crop protection. Front. Plant Sci. 2016, 7, 1132. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Malook, S.U.; Shen, G.; Gao, L.; Zhang, C.; Li, J.; Zhang, J.; Wang, L.; Wu, J. Current understanding of maize and rice defense against insect herbivores. Plant Divers. 2018, 40, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Karban, R. The ecology and evolution of induced responses to herbivory and how plants perceive risk. Ecol. Entomol. 2020, 45, 1–9. [Google Scholar] [CrossRef]
- Brooker, N.L.; Long, J.H.; Stephan, S.M. Field assessment of plant derivative compounds for managing fungal soybean diseases. Biochem. Soc. Trans. 2000, 28, 917–920. [Google Scholar] [CrossRef]
- Hilder, V.A.; Gatehouse, A.M.R.; Sheerman, S.E.; Barker, R.F.; Boulter, D. A novel mechanism of insect resistance engineered into tobacco. Nature 1987, 330, 160–163. [Google Scholar] [CrossRef]
- Adang, M.J.; Brody, M.S.; Cardineau, G.; Eagan, N.; Roush, R.T.; Shewmaker, C.K.; Jones, A.; Oakes, J.V.; McBride, K.E. The reconstruction and expression of a Bacillus thuringiensis cryIIIA gene in protoplasts and potato plants. Plant Mol. Biol. 1993, 21, 1131–1145. [Google Scholar] [CrossRef]
- Peferoen, M. Engineering of insect-resistant plants with Bacillus thuringiensis crystal protein genes. In Plant Genetic Manipulation for Crop Protection; Gatehouse, A., Hilder, V., Boulter, D., Eds.; CAB International: Wallingford, UK, 1992; pp. 135–153. [Google Scholar]
- Koziel, M.G.; Carozzi, N.B.; Currier, T.C.; Currier, T.C.; Warren, G.W.; Evola, S.V. The insecticidal crystal proteins of Bacillus thuringiensis: Past, present and future uses. Biotechnol. Genet. Eng. Rev. 1993, 11, 171–228. [Google Scholar] [CrossRef]
- Vaeck, M.; Reynaerts, A.; Höfte, H.; Jansens, S.; De Beuckeleer, M.; Dean, C.; Zabeau, M.; Montagu, M.V.; Leemans, J. Transgenic plants protected from insect attack. Nature 1987, 328, 33–37. [Google Scholar] [CrossRef]
- Potrykus, I. Gene transfer to plants: Assessment of published approaches and results. Ann. Rev. Plant Biol. 1991, 42, 205–225. [Google Scholar] [CrossRef]
- Estruch, J.J.; Carozzi, N.B.; Desai, N.; Duck, N.B.; Warren, G.W.; Koziel, M.G. Transgenic plants: An emerging approach to pest control. Nat. Biotechnol. 1997, 15, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Betz, F.S.; Hammond, B.G.; Fuchs, R.L. Safety and advantages of Bacillus thuringiensis-protected plants to control insect pests. Regul. Toxicol. Pharmacol. 2000, 32, 156–173. [Google Scholar] [CrossRef]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Control 2019, 130, 136–154. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Liesner, L.R.; Ellsworth, P.C.; Unnithan, G.C.; Fabrick, J.A.; Naranjo, S.E.; Li, X.; Dennehy, T.J.; Antilla, L.; Staten, R.T.; et al. Transgenic cotton and sterile insect releases synergize eradication of pink bollworm a century after it invaded the United States. Proc. Natl. Acad. Sci. USA 2021, 118, e2019115118. [Google Scholar] [CrossRef]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier; Cornell University: Ithaca, NY, USA, 2019. [Google Scholar]
- IRAC. Industry Perspectives on Insect Resistance Monitoring for Transgenic Insect-Protected Crops. Available online: https://irac-online.org/documents/industry-perspectives-on-ir-monitoring/?ext=pdf (accessed on 6 June 2023).
- Yates-Stewart, A.D.; Yorke, B.T.; Willse, A.; Fridley, J.; Head, G.P. Using sentinel plots to monitor for changes in thrips susceptibility to MON 88702 cotton containing the Cry51Aa2. 834_16 Bt Protein. Insects 2023, 14, 497. [Google Scholar] [CrossRef]
- Carrière, Y.; DeGain, B.; Unnithan, G.C.; Harpold, V.S.; Li, X.; Tabashnik, B.E. Seasonal declines in Cry1Ac and Cry2Ab concentration in maturing cotton favor faster evolution of resistance to pyramided Bt cotton in Helicoverpa zea (Lepidoptera: Noctuidae). J. Econ. Entomol. 2019, 112, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Fabrick, J.A.; Carrière, Y. Global patterns of insect resistance to transgenic Bt crops: The first 25 years. J. Econ. Entomol. 2023, 116, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Sachs, E.S.; Benedict, J.H.; Taylor, J.F.; Stelly, D.M.; Davis, S.K.; Altman, D.W. Pyramiding CryIA (b) insecticidal protein and terpenoids in cotton to resist tobacco budworm (Lepidoptera: Noctuidae). Environ. Entomol. 1996, 25, 1257–1266. [Google Scholar] [CrossRef]
- Guan, X.; Liu, X.; Lu, Z.; Zhao, Z.; Zhang, Q.; Xia, J. Influence of tannic acid and Cry1Ac toxin of Bacillus thuringiensis on larval survival, growth, and development of Helicoverpa armigera. Entomol. Exp. Appl. 2009, 132, 50–58. [Google Scholar] [CrossRef]
- Coombs, J.J.; Douches, D.S.; Li, W.; Grafius, E.J.; Pett, W.L. Combining engineered (Bt-cry3A) and natural resistance mechanisms in potato for control of Colorado potato beetle. J. Am. Soc. Hortic. Sci. 2002, 127, 62–68. [Google Scholar] [CrossRef]
- Hagenbucher, S.; Eisenring, M.; Meissle, M.; Romeis, J. Interaction of transgenic and natural insect resistance mechanisms against Spodoptera littoralis in cotton. Pest. Manag. Sci. 2017, 73, 1670–1678. [Google Scholar] [CrossRef]
- Abedi, Z.; Saber, M.; Vojoudi, S.; Mahdavi, V.; Parsaeyan, E. Acute, sublethal, and combination effects of azadirachtin and Bacillus thuringiensis on the cotton bollworm, Helicoverpa armigera. J. Insect Sci. 2014, 14, 30. [Google Scholar] [CrossRef]
- Mohan, S.; Ma, P.W.; Williams, W.P.; Luthe, D.S. A naturally occurring plant cysteine protease possesses remarkable toxicity against insect pests and synergizes Bacillus thuringiensis toxin. PLoS ONE 2008, 3, e1786. [Google Scholar] [CrossRef]
- Anilkumar, K.J.; Sivasupramaniam, S.; Head, G.; Orth, R.; Van Santen, E.; Moar, W.J. Synergistic interactions between Cry1Ac and natural cotton defenses limit survival of Cry1Ac-resistant Helicoverpa zea (Lepidoptera: Noctuidae) on Bt cotton. J. Chem. Ecol. 2009, 35, 785–795. [Google Scholar] [CrossRef]
- Mészáros, A.; Beuzelin, J.M.; Stout, M.J.; Bommireddy, P.L.; Rita Riggio, M.; Rogers Leonard, B. Jasmonic acid-induced resistance to the fall armyworm, Spodoptera frugiperda, in conventional and transgenic cottons expressing Bacillus thuringiensis insecticidal proteins. Entomol. Exp. Appl. 2011, 140, 226–237. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, M.; Huang, J.; Li, L.; Huang, K.; Zhang, Y.; Li, Y.; Deng, Z.; Ni, X.; Li, X. Inductive and synergistic interactions between plant allelochemical flavone and Bt toxin Cry1Ac in Helicoverpa armigera. Insect Sci. 2021, 28, 1756–1765. [Google Scholar] [CrossRef] [PubMed]
- Olsen, K.M.; Daly, J.C. Plant-toxin interactions in transgenic Bt cotton and their effect on mortality of Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2000, 93, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guan, X.; Michaud, J.P.; Zhang, Q.; Liu, X. Quercetin interacts with Cry1Ac protein to affect larval growth and survival of Helicoverpa armigera. Pest. Manag. Sci. 2016, 72, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Cai, Y.S.; Zhu, J.L. The study on join-toxicity of Bacillus thuringiensis with insecticides. Jiangsu Agric. Sci. 2001, 010, 45–46. [Google Scholar]
- Nix, A.; Paull, C.; Colgrave, M. Flavonoid profile of the cotton plant, Gossypium hirsutum: A review. Plants 2017, 6, 43. [Google Scholar] [CrossRef]
- Mathesius, U. Flavonoid functions in plants and their interactions with other organisms. Plants 2018, 7, 30. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Xiao, Y.; Wu, K. Recent progress on the interaction between insects and Bacillus thuringiensis crops. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180316. [Google Scholar] [CrossRef]
- Koffi, D.; Kyerematen, R.; Osae, M.; Amouzou, K.; Eziah, V.Y. Assessment of Bacillus thuringiensis and emamectin benzoate on the fall armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) severity on maize under farmers’ fields in Ghana. Int. J. Trop. Insect Sci. 2022, 42, 1619–1626. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, J.; Wang, K.; Zhang, Y.; Liu, Z.; Yu, N. A Binary mixture of emamectin benzoate and chlorantraniliprole supplemented with an adjuvant effectively controls Spodoptera frugiperda. Insects 2022, 13, 1157. [Google Scholar] [CrossRef]
- Bird, L.; Miles, M.; Quade, A.; Spafford, H. Insecticide resistance in Australian Spodoptera frugiperda (J.E. Smith) and development of testing procedures for resistance surveillance. PLoS ONE 2022, 17, e0263677. [Google Scholar] [CrossRef]
- Zhang, D.-d.; Xiao, Y.-t.; Xu, P.-j.; Yang, X.-m.; Wu, Q.-l.; Wu, K.-m. Insecticide resistance monitoring for the invasive populations of fall armyworm, Spodoptera frugiperda in China. J. Integr. Agric. 2021, 20, 783–791. [Google Scholar] [CrossRef]
- Zhao, R.; Wang, H.H.; Gao, J.; Zhang, Y.J.; Li, X.; Zhou, J.; Liang, P.; Gao, X.W.; Gu, S.H. Plant volatile compound methyl benzoate is highly effective against Spodoptera frugiperda and safe to non-target organisms as an eco-friendly botanical-insecticide. Ecotoxicol. Environ. Saf. 2022, 245, 114101. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, G.P.; Friedman, S. Self-selection of optimal diets by insects. Ann. Rev. Entomol. 1991, 36, 43–63. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. 1925. J. Am. Mosq. Control Assoc. 1987, 3, 302–303. [Google Scholar] [PubMed]
- Mansour, N.A.; Eldefrawi, M.E.; Toppozada, A.; Zeid, M. Toxicological studies on the egyptian cotton leaf worm, Prodenia litura. VI. Potentiation and antagonism of organophosphorus and carbamate insecticides. J. Econ. Entomol. 1966, 59, 307–311. [Google Scholar] [CrossRef]
- Bergamasco, V.B.; Mendes, D.R.; Fernandes, O.A.; Desidério, J.A.; Lemos, M.V. Bacillus thuringiensis Cry1Ia10 and Vip3Aa protein interactions and their toxicity in Spodoptera spp. (Lepidoptera). J. Invertebr. Pathol. 2013, 112, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Zhang, D.; Zhang, R.; Wang, Y.; Meng, Z.; Zhou, T.; Liang, C.; Zhu, T.; Guo, S. Bt protein expression in the transgenic insect-resistant cotton in China. Sci. Bull. 2016, 61, 1555–1557. [Google Scholar] [CrossRef]
- Carrière, Y.; Degain, B.A.; Unnithan, G.C.; Harpold, V.S.; Heuberger, S.; Li, X.; Tabashnik, B.E. Effects of seasonal changes in cotton plants on the evolution of resistance to pyramided cotton producing the Bt toxins Cry1Ac and Cry1F in Helicoverpa zea. Pest. Manag. Sci. 2018, 74, 627–637. [Google Scholar] [CrossRef]
- Justesen, H.; Andersen, A.S.; Brandt, K. Accumulation of anthocyanins and flavones during bud and flower development in Campanula isophylla Moretti. Ann. Bot. 1997, 79, 355–360. [Google Scholar] [CrossRef]
- Sunila, A.V.; Murugan, K. Variation in phenolics, flavonoids at different stages of fruit development of Pouteria campechiana (Kunth) baehni and its antioxidant activity. Int. J. Pharm. Pharm. Sci. 2017, 9, 70–75. [Google Scholar] [CrossRef]
- Chang, X.; Lu, Y.; Lin, Z.; Qiu, J.; Guo, X.; Pan, J.; Abbasi, A.M. Impact of leaf development stages on polyphenolics profile and antioxidant activity in Clausena lansium (Lour.) Skeels. Biomed. Res. Int. 2018, 2018, 7093691. [Google Scholar] [CrossRef] [PubMed]
- Ayra-Pardo, C.; Ochagavía, M.E.; Raymond, B.; Gulzar, A.; Rodríguez-Cabrera, L.; Rodríguez de la Noval, C.; Morán Bertot, I.; Terauchi, R.; Yoshida, K.; Matsumura, H.; et al. HT-SuperSAGE of the gut tissue of a Vip3Aa-resistant Heliothis virescens (Lepidoptera: Noctuidae) strain provides insights into the basis of resistance. Insect Sci. 2019, 26, 479–498. [Google Scholar] [CrossRef]
- Jin, M.; Shan, Y.; Peng, Y.; Wang, P.; Li, Q.; Yu, S.; Zhang, L.; Xiao, Y. An Integrative Analysis of Transcriptomics and Proteomics Reveals novel insights into the response in the midgut of Spodoptera frugiperda Larvae to Vip3Aa. Toxins 2022, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Chen, C.; Wu, S.; Shao, E.; Li, M.; Guan, X.; Huang, Z. Transcriptional profiling analysis of Spodoptera litura larvae challenged with Vip3Aa toxin and possible involvement of trypsin in the toxin activation. Sci. Rep. 2016, 6, 23861. [Google Scholar] [CrossRef]
- Lu, K.; Cheng, Y.; Li, Y.; Li, W.; Zeng, R.; Song, Y. Phytochemical flavone confers broad-spectrum tolerance to insecticides in Spodoptera litura by activating ROS/CncC-mediated xenobiotic detoxification pathways. J. Agric. Food Chem. 2021, 69, 7429–7445. [Google Scholar] [CrossRef]
- Deng, Z.; Zhang, Y.; Fang, L.; Zhang, M.; Wang, L.; Ni, X.; Li, X. Identification of the flavone-inducible counter-defense genes and their cis-elements in Helicoverpa armigera. Toxins 2023, 15, 365. [Google Scholar] [CrossRef]
- Khan, M.M.; Khan, A.H.; Ali, M.W.; Hafeez, M.; Ali, S.; Du, C.; Fan, Z.; Sattar, M.; Hua, H. Emamectin benzoate induced enzymatic and transcriptional alternation in detoxification mechanism of predatory beetle Paederus fuscipes (Coleoptera: Staphylinidae) at the sublethal concentration. Ecotoxicology 2021, 30, 1227–1241. [Google Scholar] [CrossRef]
- Sun, Y.; Tao, S.; Zhang, W.; Jiang, B.; Dai, H.-Y.; Liu, B.-S.; Zhang, Y.-J.; Kong, X.-d.; Zhao, J.; Bai, L.-X. Transcriptome profile analysis reveals the emamectin benzoate-induced genes associated with olfaction and metabolic detoxification in Spodoptera exigua Hübner (Lepidoptera: Noctuidae). All. Life 2022, 15, 340–357. [Google Scholar] [CrossRef]
- Kennedy, G.G.; Farrar Jr, R.R. Response of insecticide-resistant and susceptible Colorado potato beetles, Leptinotarsa decemlineata to 2-tridecanone and resistant tomato foliage: The absence of cross resistance. Entomol. Exp. Appl. 1987, 45, 187–192. [Google Scholar] [CrossRef]
- Gould, F.; Carroll, C.R.; Futuyma, D.J. Cross-resistance to pesticides and plant defenses: A study of the two-spotted spider mite. Entomol. Exp. Appl. 1982, 31, 175–180. [Google Scholar] [CrossRef]
- Li, X.; Zangerl, A.R.; Schuler, M.A.; Berenbaum, M.R. Cross-resistance to alpha-cypermethrin after xanthotoxin ingestion in Helicoverpa zea (Lepidoptera: Noctuidae). J. Econ. Entomol. 2000, 93, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, F.; Tong, H.; Hu, Y.; Zhang, X.; Tian, T.; Zhang, Y.; Su, Q. Plant flavonoids enhance the tolerance to thiamethoxam and flupyradifurone in whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Pestic. Biochem. Physiol. 2021, 171, 104744. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Elzaki, M.E.A.; Ding, C.; Li, Z.F.; Wang, J.; Zeng, R.S.; Song, Y.Y. Plant allelochemicals affect tolerance of polyphagous lepidopteran pest Helicoverpa armigera (Hübner) against insecticides. Pestic. Biochem. Physiol. 2019, 154, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Trapero, C.; Wilson, I.W.; Stiller, W.N.; Wilson, L.J. Enhancing integrated pest management in GM cotton systems using host plant resistance. Front. Plant Sci. 2016, 7, 500. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxins | N a | LD-P Line | LC5 (95% FL b) | LC25 (95% FL b) | LC50 (95% FL b) | R2 c |
|---|---|---|---|---|---|---|
| Vip3Aa | 285 | Y = 1.97X + 1.56 | 24 d (1–50) | 73 (19–130) | 161 (90–370) | 0.89 |
| Flavone | 275 | Y = 3.05X − 8.27 | 147.95 e (27.0–244.6) | 307.5 (142.1–348.1) | 511.1 (354.6–801.8) | 0.91 |
| Emamectin benzoate | 285 | Y = 3.50X + 2.99 | 0.047 d (0.03–0.06) | 0.09 (0.074–0.1) | 0.14 (0.12–0.16) | 0.96 |
| Exposure to | Observed Mortality (%) | EAM * (%) | Co-Toxicity Factor | Interaction | |
|---|---|---|---|---|---|
| Concurrent Vip3Aa_LC25 + Fla_LC25 | Vip3Aa | 35.42 ± 2.08 a | |||
| Fla b | 25.00 ± 3.61 | ||||
| Vip3Aa + Fla | 81.25 ± 3.61 | 60.42 | +20 < 34.48 | Synergism | |
| Concurrent Vip3Aa_LC25 + EMB_LC25 | Vip3Aa | 43.61 ± 3.06 | |||
| EMB c | 31.65 ± 7.01 | ||||
| Vip3Aa + EMB | 95.83 ± 2.08 | 75.26 | +20 < 27.34 | Synergism | |
| Concurrent Fla_LC25 + EMB_LC25 | Fla | 25.00 ± 3.61 | |||
| EMB | 41.67 ± 4.17 | ||||
| Vip3Aa + EMB | 70.83 ± 4.17 | 66.67 | +20 > 6.25 | Addition | |
| Concurrent Fla_LC15 + EMB_LC15 + Vip3Aa_LC15 | Fla | 14.58 ± 4.17 | |||
| EMB | 19.17 ± 3.63 | ||||
| Vip3Aa | 29.17 ± 5.51 | ||||
| Fla + EMB + Vip3Aa | 87.50 ± 0.00 | 62.92 | +20 < 40.00 | Synergism | |
| Exposure to | Observed Mortality (%) | EAM * (%) | Co-Toxicity Factor | Interaction | |
|---|---|---|---|---|---|
| Sequential Vip3Aa_LC5 + Fla_LC50 | Vip3Aa | 8.33 ± 2.08 a | |||
| Fla | 50.00 ± 3.61 | ||||
| Vip3Aa + Fla b | 62.08 ± 9.14 | 58.33 | −20 < 6.43 < +20 | Addition | |
| Sequential Fla_LC5 + Vip3Aa_LC50 | Fla | 6.25 ± 0.00 | |||
| Vip3Aa | 37.50 ± 13.01 | ||||
| Fla + Vip3Aa | 81.25 ± 3.61 | 43.75 | +20 < 85.71 | Induction | |
| Sequential Fla_LC5 + EMB_LC50 | Fla | 12.5 ± 0.00 | |||
| EMB c | 33.33 ± 4.17 | ||||
| Fla + EMB | 25.00 ± 3.61 | 45.83 | −45.45 < −20 | Inhibition | |
| Sequential EMB_LC5 + Fla_LC50 | EMB | 0.00 ± 0.00 | |||
| Fla | 50.00 ± 3.61 | ||||
| EMB + Fla | 56.25 ± 3.61 | 50.00 | −20 < 12.50 < +20 | Addition | |
| Sequential Vip3Aa_LC5 + EMB_LC50 | Vip3Aa | 4.17 ± 4.17 | |||
| EMB | 42.50 ± 7.32 | ||||
| Vip3Aa + EMB | 49.40 ± 6.37 | 46.67 | −20 < 5.86 < +20 | Addition | |
| Sequential EMB_LC5 + Vip3Aa_LC50 | EMB | 2.08 ± 2.08 | |||
| Vip3Aa | 33.33 ± 2.08 | ||||
| EMB + Vip3Aa | 52.08 ± 2.08 | 35.42 | +20 < 47.06 | Induction | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, K.; He, H.; Wang, S.; Zhang, M.; Chen, X.; Deng, Z.; Ni, X.; Li, X. Sequential and Simultaneous Interactions of Plant Allelochemical Flavone, Bt Toxin Vip3A, and Insecticide Emamectin Benzoate in Spodoptera frugiperda. Insects 2023, 14, 736. https://doi.org/10.3390/insects14090736
Huang K, He H, Wang S, Zhang M, Chen X, Deng Z, Ni X, Li X. Sequential and Simultaneous Interactions of Plant Allelochemical Flavone, Bt Toxin Vip3A, and Insecticide Emamectin Benzoate in Spodoptera frugiperda. Insects. 2023; 14(9):736. https://doi.org/10.3390/insects14090736
Chicago/Turabian StyleHuang, Kaiyuan, Haibo He, Shan Wang, Min Zhang, Xuewei Chen, Zhongyuan Deng, Xinzhi Ni, and Xianchun Li. 2023. "Sequential and Simultaneous Interactions of Plant Allelochemical Flavone, Bt Toxin Vip3A, and Insecticide Emamectin Benzoate in Spodoptera frugiperda" Insects 14, no. 9: 736. https://doi.org/10.3390/insects14090736