
Acaricidal Toxicity of Four Essential Oils, Their Predominant Constituents, Their Mixtures against Varroa Mite, and Their Selectivity to Honey Bees (Apis cerana and A. mellifera)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Plant Materials
2.1.2. Chemicals
2.1.3. GC-MS Instrument
2.1.4. Varroa Mite Collection
2.1.5. Honey Bees
2.2. Methods
2.2.1. Extraction of Essential Oils from Plant Species
2.2.2. GC-MS Analysis Conditions
2.2.3. Toxicity of Essential Oils and Their Major Constituents against V. destructor
2.2.4. Toxicity of T. ammi and T. schimperi on Honey Bees
Surface Treatment Bioassay
Topical Application Bioassay
2.2.5. Learning and Memory Bioassay
2.2.6. Statistical Analysis
3. Results
3.1. Chemical Composition of Essential Oils
3.2. Acaricidal Activities of EOs and Their Main Components against V. destructor
3.3. Honey Bee Toxicity
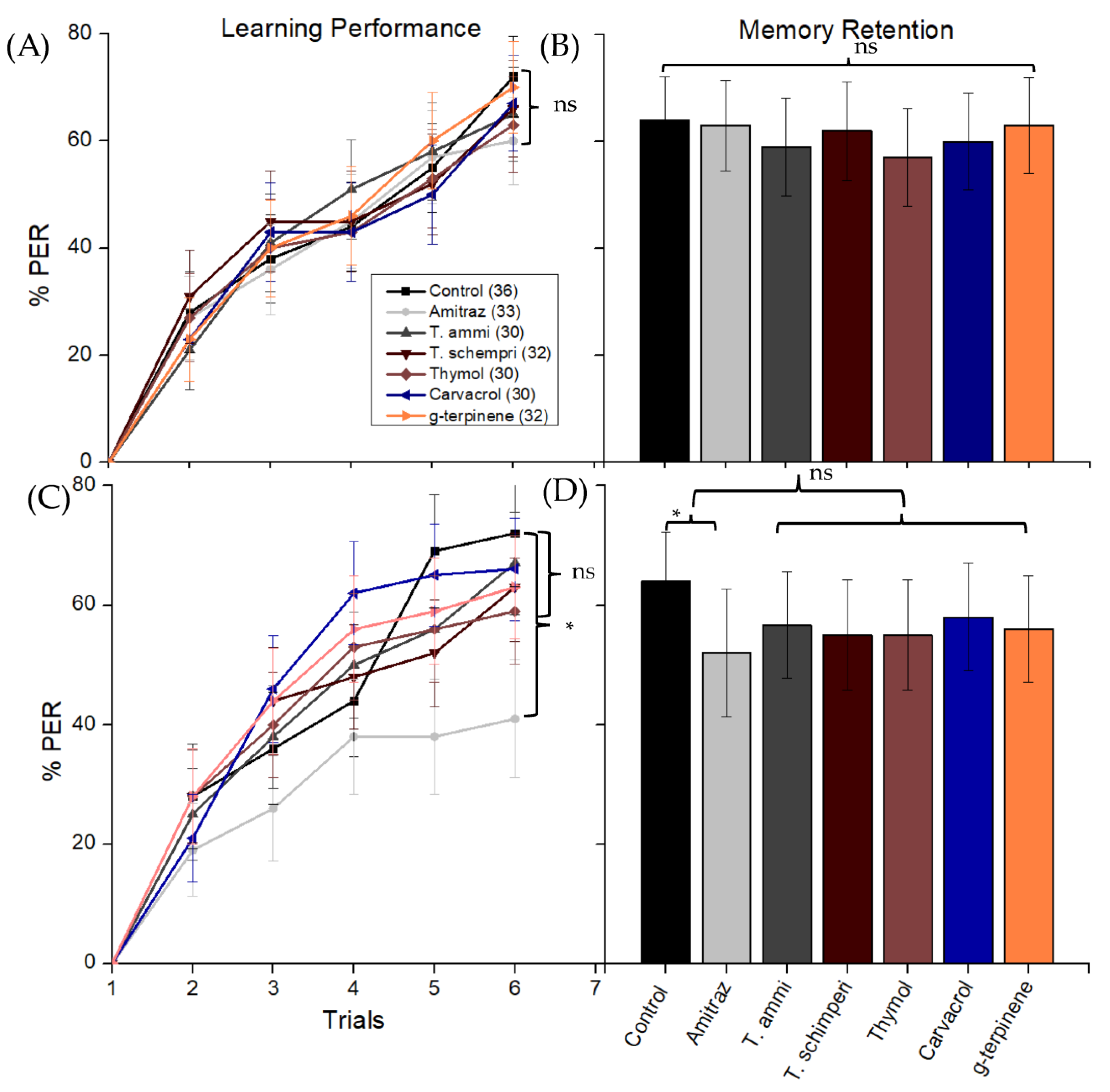
3.4. Learning and Memory Bioassay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, W.; Wang, C.; Huang, Z.Y.; Chen, Y.; Han, R. Reproduction of distinct Varroa destructor genotypes on honey bee worker brood. Insects 2019, 10, 372. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Leclercq, G.; Gengler, N.; Francis, F. How human reshaped diversity in honey bees (Apis mellifera L.): A review. Entomol. Faun. 2018, 71, 1–13. [Google Scholar]
- Goblirsch, M. Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie 2018, 49, 131–150. [Google Scholar] [CrossRef]
- Le Conte, Y.; Huang, Z.; Roux, M.; Zeng, Z.; Christidès, J.P.; Bagnères, A.G. Varroa destructor changes its cuticular hydrocarbons to mimic new hosts. Biol. Lett. 2015, 11, 20150233. [Google Scholar] [CrossRef]
- Giacobino, A.; Molineri, A.I.; Pacini, A.; Fondevila, N.; Pietronave, H.; Rodríguez, G.; Palacio, A.; Bulacio Cagnolo, N.; Orellano, E.; Salto, C.E. Varroa destructor and viruses association in honey bee colonies under different climatic conditions. Environ. Microbiol. Rep. 2016, 8, 407–412. [Google Scholar] [CrossRef]
- Guichard, M.; Dietemann, V.; Neuditschko, M.; Dainat, B. Advances and perspectives in selecting resistance traits against the parasitic mite Varroa destructor in honey bees. Genet. Sel. Evol. 2020, 52, 1–22. [Google Scholar] [CrossRef]
- Schüler, V.; Liu, Y.C.; Gisder, S.; Horchler, L.; Groth, D.; Genersch, E. Significant, but not biologically relevant: Nosema ceranae infections and winter losses of honey bee colonies. Commun. Biol. 2023, 6, 229. [Google Scholar] [CrossRef]
- Kluser, S.; Neumann, P.; Chauzat, M.P.; Pettis, J.S.; Peduzzi, P.; Witt, R.; Fernandez, N.; Theuri, M. UNEP Emerging Issue: Global honey bee colony disorders and other threats to insect pollinators. 2010, pp. 1–17. Available online: http://archive-ouverte.unige.ch/unige:32251 (accessed on 3 February 2023).
- Jeong, S.; Lee, C.; Kim, D.; Jung, C. Questionnaire study on the overwintering success and pest management of honey bee and damage assessment of Vespa hornets in Korea. Korean J. Apic. 2016, 31, 201–210. [Google Scholar] [CrossRef]
- Steinhauer, N.; Aurell, D.; Bruckner, S.; Wilson, M.; Rennich, K.; VanEngelsdorp, D.; Williams, G.; for the Bee Informed Partnership. United States Honey Bee Colony Losses 2020–2021: Preliminary Results. Available online: https://beeinformed.org/wpcontent/uploads/2021/06/BIP_2020_21_Losses_Abstract_2021.06.14_FINAL_R1.pdf (accessed on 28 March 2023).
- Stahlmann-Brown, P.; Hall, R.J.; Pragert, H.; Robertson, T. Varroa appears to drive persistent increases in New Zealand colony losses. Insects 2022, 13, 589. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, V.; Moharramipour, S.; Tahmasbi, G. Biological activity of some plant essential oils against Varroa destructor (Acari: Varroidae), an ectoparasitic mite of Apis mellifera (Hymenoptera: Apidae). Exp. Appl. Acarol. 2011, 55, 147–154. [Google Scholar] [CrossRef]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; VanEngelsdorp, D.; Pettis, J.S. High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef]
- Bogdanov, S.; Charrière, J.D.; Imdorf, A.; Kilchenmann, V.; Fluri, P. Determination of residues in honey after treatments with formic and oxalic acid under field conditions. Apidologie 2002, 33, 399–409. [Google Scholar] [CrossRef]
- de Mattos, I.M.; Soares, A.E.; Tarpy, D.R. Effects of synthetic acaricides on honey bee grooming behavior against the parasitic Varroa destructor mite. Apidologie 2017, 48, 483–494. [Google Scholar] [CrossRef]
- Tihelka, E. Effects of synthetic and organic acaricides on honey bee health: A review. Slov. Vet. Res. 2018, 55, 114–140. [Google Scholar] [CrossRef]
- Chaimanee, V.; Warrit, N.; Boonmee, T.; Pettis, J.S. Acaricidal activity of essential oils for the control of honey bee (Apis mellifera) mites Tropilaelaps mercedesae under laboratory and colony conditions. Apidologie 2021, 52, 561–575. [Google Scholar] [CrossRef]
- Stanimirović, Z.; Glavinić, U.; Ristanić, M.; Jelisić, S.; Vejnović, B.; Niketić, M.; Stevanović, J. Diet supplementation helps honey bee colonies in combat infections by enhancing their hygienic behaviour. Acta. Vet. Beogr. 2022, 72, 145–166. [Google Scholar] [CrossRef]
- Stanimirović, Z.; Glavinić, U.; Lakić, N.; Radović, D.; Ristanić, M.; Tarić, E.; Stevanović, J. Efficacy of plant-derived formulation Argus Ras in Varroa destructor control. Acta Vet. Beogr. 2017, 67, 191–200. [Google Scholar] [CrossRef]
- Zengin, G.; Mahomoodally, M.F.; Yıldıztugay, E.; Jugreet, S.; Khan, S.U.; Dall’Acqua, S.; Mollica, A.; Bouyahya, A.; Montesano, D. Chemical composition, biological activities and in silico analysis of essential oils of three endemic Prangos species from Turkey. Molecules 2022, 27, 1676. [Google Scholar] [CrossRef]
- Djilani, A.; Dicko, A. The therapeutic benefits of essential oils. Nutr. Well-Being Health 2012, 7, 155–179. [Google Scholar]
- Lindberg, C.M.; Melathopoulos, A.P.; Winston, M.L. Laboratory evaluation of miticides to control Varroa jacobsoni (Acari: Varroidae), a honey bee (Hymenoptera: Apidae) parasite. Econ. Entomol. 2000, 93, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Gashout, H.A.; Guzmán-Novoa, E. Acute toxicity of essential oils and other natural compounds to the parasitic mite, Varroa destructor, and to larval and adult worker honey bees (Apis mellifera L.). J. Apic. Res. 2009, 48, 263–269. [Google Scholar] [CrossRef]
- Calderone, N.W.; Wilson, W.T.; Spivak, M. Plant extracts used for control of the parasitic mites Varroa jacobsoni (Acari: Varroidae) and Acarapis woodi (Acari: Tarsonemidae) in colonies of Apis mellifera (Hymenoptera: Apidae). J. Econ. Entomol. 1997, 90, 1080–1086. [Google Scholar] [CrossRef]
- Glavan, G.; Novak, S.; Božič, J.; Kokalj, A.J. Comparison of sublethal effects of natural acaricides carvacrol and thymol on honey bees. Pestic. Biochem. Physiol. 2020, 166, 104567. [Google Scholar] [CrossRef] [PubMed]
- Imdorf, A.; Bogdanov, S.; Kilchenmann, V.; Berger, T. Toxic effects of essential oils and some of their components on Varroa destructor Oud and Apis mellifera L. under laboratory conditions. ALP Sci. 2006, 495, 1–18. [Google Scholar]
- Bisrat, D.; Begna, T.; Ulziibayar, D.; Jung, C. Acaricidal activity of essential oil-derived components from Thymus schimperi Ronninger against Varroa destructor Anderson and Trueman. J. Apic. Res. 2022, 1–7. [Google Scholar] [CrossRef]
- Bava, R.; Castagna, F.; Palma, E.; Marrelli, M.; Conforti, F.; Musolino, V.; Carresi, C.; Lupia, C.; Ceniti, C.; Tilocca, B. Essential oils for a sustainable control of honey bee Varroosis. Vet. Sci. 2023, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Umpiérrez, M.L.; Santos, E.; González, A.; Rossini, C. Plant essential oils as potential control agents of varroatosis. Phytochem. Rev. 2011, 10, 227–244. [Google Scholar] [CrossRef]
- Aglagane, A.; Laghzaoui, E.M.; Ben Elfakir, S.; Er-Rguibi, O.; Abbad, A.; El Mouden, E.H.; Aourir, M. Essential oils as sustainable control agents against Varroa destructor (Acari, Varroidae), an ectoparasitic mite of the western honey bees Apis mellifera (Hymenoptera: Apidae): Review of recent literature (2010-onwards). Int. J. Acarol. 2021, 47, 436–445. [Google Scholar] [CrossRef]
- Aglagane, A.; Laghzaoui, E.M.; Soulaimani, B.; Er-Rguibi, O.; Abbad, A.; Mouden, E.H.E.; Aourir, M. Acaricidal activity of Mentha suaveolens subsp. timija, Chenopodium ambrosioides, and Laurus nobilis essential oils, and their synergistic combinations against the ectoparasitic bee mite, Varroa destructor (Acari: Varroidae). J. Apic. Res. 2022, 61, 9–18. [Google Scholar] [CrossRef]
- Kebebe, D.; Gela, A.; Damto, T.; Gemeda, M.; Leggese, G. Evaluating the Effect of Plants Extracts against Varroa Mites (Varroa destructors) of honey bees (Apis mellifera). J. Chem. Mater. Res. 2022, 14, 26–30. [Google Scholar]
- Sabahi, Q.; Kelly, P.G.; Guzman-Novoa, E. Carvone and citral, two promising compounds for controlling the honey bee ectoparasitic mite, Varroa destructor. J. Appl. Entomol. 2022, 146, 1003–1010. [Google Scholar] [CrossRef]
- Giménez-Martínez, P.; Ramirez, C.; Mitton, G.; Arcerito, F.M.; Ramos, F.; Cooley, H.; Fuselli, S.; Maggi, M. Lethal concentrations of Cymbopogon nardus essential oils and their main component citronellal on Varroa destructor and Apis mellifera. Exp. Parasitol. 2022, 238, 108279. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R.; Alaux, C.; Costa, C.; Csaki, T.; Doublet, V.; Eisenhardt, D.; Fries, I.; Kuhn, R.; McMahon, D.P.; Medrzycki, P. Standard methods for maintaining adult Apis mellifera in cages under in vitro laboratory conditions. J. Apic. Res. 2013, 52, 1–36. [Google Scholar] [CrossRef]
- Piiroinen, S.; Goulson, D. Chronic neonicotinoid pesticide exposure and parasite stress differentially affects learning in honey bees and bumblebees. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160246. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Menzel, R.; Sandoz, J.C.; Giurfa, M. Revisiting olfactory classical conditioning of the proboscis extension response in honey bees: A step toward standardized procedures. J. Neurosci. Methods 2012, 211, 159–167. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publ. Corp.: Carol Stream, IL, USA, 2007. [Google Scholar]
- Babushok, V.; Linstrom, P.; Zenkevich, I. Retention indices for frequently reported compounds of plant essential oils. J. Phys. Chem. Ref. Data 2011, 40, 043101. [Google Scholar] [CrossRef]
- Tak, J.H.; Jovel, E.; Isman, M.B. Comparative and synergistic activity of Rosmarinus officinalis L. essential oil constituents against the larvae and an ovarian cell line of the cabbage looper, Trichoplusia ni (Lepidoptera: Noctuidae). Pest Manag. Sci. 2016, 72, 474–480. [Google Scholar] [CrossRef]
- Sabahi, Q.; Hamiduzzaman, M.M.; Barajas-Pérez, J.S.; Tapia-Gonzalez, J.M.; Guzman-Novoa, E. Toxicity of anethole and the essential oils of lemongrass and sweet marigold to the parasitic mite Varroa destructor and their selectivity for honey bee (Apis mellifera) workers and larvae. Psyche 2018, 2018, 6196289. [Google Scholar] [CrossRef]
- da Silva, I.M.; Zanuncio, J.C.; Brügger, B.P.; Soares, M.A.; Zanuncio, A.J.V.; Wilcken, C.F.; Tavares, W.D.; Serrão, J.E.; Sediyama, C.S. Selectivity of the botanical compounds to the pollinators Apis mellifera and Trigona hyalinata (Hymenoptera: Apidae). Sci. Rep. 2020, 10, 4820. [Google Scholar] [CrossRef]
- Human, H.; Brodschneider, R.; Dietemann, V.; Dively, G.; Ellis, J.D.; Forsgren, E.; Fries, I.; Hatjina, F.; Hu, F.L.; Jaffé, R. Miscellaneous standard methods for Apis mellifera research. J. Apic. Res. 2013, 52, 1–53. [Google Scholar] [CrossRef]
- Stanley, J.; Preetha, G. Pesticide toxicity to arthropod predators: Exposure, Toxicity and Risk Assessment Methodologies. In Pesticide Toxicity to Non-Target Organisms; Springer Nature: Dordrecht, The Netherlands, 2016; pp. 1–98. [Google Scholar]
- Wang, Y.; Zhu, Y.C.; Li, W. Interaction patterns and combined toxic effects of acetamiprid in combination with seven pesticides on honey bee (Apis mellifera L.). Ecotoxicol. Environ. Saf. 2020, 190, 110100. [Google Scholar] [CrossRef]
- Türkmen, M.; Kara, M.; Maral, H.; Soylu, S. Determination of chemical component of essential oil of Origanum dubium plants grown at different altitudes and antifungal activity against Sclerotinia sclerotiorum. J. Food Process. Preserv. 2021, 46, e15787. [Google Scholar] [CrossRef]
- Chauhan, B.; Kumar, G.; Ali, M. A review on phytochemical constituents and activities of Trachyspermum ammi (L.) Sprague fruits. Am. J. Pharmtech. Res. 2012, 2, 329–340. [Google Scholar]
- Singh, G.; Maurya, S.; Catalan, C.; De Lampasona, M. Chemical constituents, antifungal and antioxidative effects of ajwain essential oil and its acetone extract. J. Agric. Food Chem. 2004, 52, 3292–3296. [Google Scholar] [CrossRef]
- Bisrat, D.; Jung, C. Insecticidal toxicities of three main constituents derived from Trachyspermum ammi (L.) Sprague ex Turrill fruits against the small hive beetles, Aethina tumida Murray. Molecules 2020, 25, 1100. [Google Scholar] [CrossRef] [PubMed]
- Baran, A.I.; Jahanghiri, F.; Hajipour, N.; Sparagano, O.A.E.; Norouzi, R.; Moharramnejad, S. In vitro acaricidal activity of essential oil and alcoholic extract of Trachyspermum ammi against Dermanyssus gallinae. Vet. Parasitol. 2020, 278, 109030. [Google Scholar] [CrossRef]
- Pandiyan, G.N.; Mathew, N.; Munusamy, S. Larvicidal activity of selected essential oil in synergized combinations against Aedes aegypti. Ecotoxicol. Environ. Saf. 2019, 174, 549–556. [Google Scholar] [CrossRef]
- Piri, A.; Sahebzadeh, N.; Zibaee, A.; Sendi, J.J.; Shamakhi, L.; Shahriari, M. Toxicity and physiological effects of ajwain (Carum copticum, Apiaceae) essential oil and its major constituents against Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Chemosphere 2020, 256, 127103. [Google Scholar] [CrossRef]
- Imdorf, A.; Bogdanov, S.; Kilchenmann, V.; Maquelin, C. Apilife VAR: A new varroacide with thymol as the main ingredient. Bee World 1995, 76, 77–83. [Google Scholar] [CrossRef]
- Kouache, B.; Brada, M.; Saadi, A.; Fauconnier, M.L.; Lognay, G.; Heuskin, S. Chemical composition and acaricidal activity of Thymus algeriensis essential oil against Varroa destructor. Nat. Prod. Commun. 2017, 12, 135–138. [Google Scholar] [CrossRef]
- Çetin, H.; Cilek, J.E.; Oz, E.; Aydin, L.; Deveci, O.; Yanikoglu, A. Acaricidal activity of Satureja thymbra L. essential oil and its major components, carvacrol and γ-terpinene against adult Hyalomma marginatum (Acari: Ixodidae). Vet. Parasitol. 2010, 170, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, J.R.; Choi, D.R.; Ahn, Y.J. Toxicity of cassia and cinnamon oil compounds and cinnamaldehyde-related compounds to Sitophilus oryzae (Coleoptera: Curculionidae). J. Econ. Entomol. 2008, 101, 1960–1966. [Google Scholar] [CrossRef]
- Brasesco, C.; Gende, L.; Negri, P.; Szawarski, N.; Iglesias, A.; Eguaras, M.; Ruffinengo, S.; Maggi, M. Assessing in vitro acaricidal effect and joint action of a binary mixture between essential oil compounds (thymol, phellandrene, eucalyptol, cinnamaldehyde, myrcene, carvacrol) over ectoparasitic mite Varroa destructor (Acari: Varroidae). J. Apic. Sci. 2017, 61, 203–215. [Google Scholar] [CrossRef]
- Rahimi, A.; Del, Y.K.; Moradpour, F. The effect of thyme (Thymus caucasicus) ethanol extract on Varroa mite (Varroa destructor), an ectoparasite mite of Apis mellifera meda (Hym: Apidae). Biologija 2017, 63, 177–184. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Soliman, R.M.; Salam, R.A.A.; Eid, B.G.; Khayyat, A.; Neamatallah, T.; Mesbah, M.K.; Hadad, G.M. Stability study of thymoquinone, carvacrol and thymol using HPLC-UV and LC-ESI-MS. Acta Pharm. 2020, 70, 325–342. [Google Scholar] [CrossRef]
- Atmani-Merabet, G.; Fellah, S.; Belkhiri, A. Comparative study of two Eucalyptus species from Algeria: Chemical composition, toxicity and acaricidal effect on Varroa destructor. Curr. Issues Pharm. Med. Sci. 2020, 33, 144–148. [Google Scholar] [CrossRef]
- El-Zemity, S.; Rezk, H.; Zaitoon, A. Acaricidal activity of some essential oils and their monoterpenoidal constituents against the parasitic bee mites, Varroa destructor (Acari: Varroidae). J. Appl. Sci. Res. 2006, 2, 1032–1036. [Google Scholar]
- Abdelgaleil, S.A.; Mohamed, M.I.; Badawy, M.E.; El-arami, S.A. Fumigant and contact toxicities of monoterpenes to Sitophilus oryzae (L.) and Tribolium castaneum (Herbst) and their inhibitory effects on acetylcholinesterase activity. J. Chem. Ecol. 2009, 35, 518–525. [Google Scholar] [CrossRef]
- Chen, Z.; van Mol, W.; Vanhecke, M.; Duchateau, L.; Claerebout, E. Acaricidal activity of plant-derived essential oil components against Psoroptes ovis in vitro and in vivo. Parasites Vectors 2019, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ruffinengo, S.; Maggi, M.; Faverin, C.; Rosa, S.B.; Bailac, P.; Principal, J.; Eguaras, M. Essential oils toxicity related to Varroa destructor and Apis mellifera under laboratory conditions. Zootec. Trop. 2007, 25, 63–69. [Google Scholar]
- Albo, G.N.; Henning, C.; Ringuelet, J.; Reynaldi, F.J.; De Giusti, M.R.; Alippi, A.M. Evaluation of some essential oils for the control and prevention of American Foulbrood disease in honey bees. Apidologie 2003, 34, 417–427. [Google Scholar] [CrossRef]
- Tiwari, R.; Mathur, V.; Bisht, B. Efficacy of cow urine and plant products against honey bee mite, Varroa destructor (Anderson and Trueman) in Apis mellifera (Linnaeus) colonies in Uttarakhand—A novel approach. Bioscan 2014, 9, 1425–1429. [Google Scholar]
- Rashid, B.; Khani, A.; Ghasemi, V.; Ghadamyari, M.; Sahebzadeh, N.; Moharramipour, S. Evaluation of a new plant-based formulation for the treatment of varroosis in the honey bee colonies: Efficacy and safety. Apidologie 2020, 51, 1074–1090. [Google Scholar] [CrossRef]
- Yue, M.; Luo, S.; Liu, J.; Wu, J. Apis cerana is less sensitive to most neonicotinoids, despite of their smaller body mass. J. Econ. Entomol. 2018, 111, 39–42. [Google Scholar] [CrossRef]
- Rafael Valdovinos-Nunez, G.; Quezada-Euán, J.J.G.; Ancona-Xiu, P.; Moo-Valle, H.; Carmona, A.; Ruiz Sanchez, E. Comparative toxicity of pesticides to stingless bees (Hymenoptera: Apidae: Meliponini). J. Econ. Entomol. 2009, 102, 1737–1742. [Google Scholar] [CrossRef]
- Cho, S.; Lee, S.H.; Kim, S. Determination of the optimal maturation temperature for adult honey bee toxicity testing. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 257, 109359. [Google Scholar] [CrossRef]
- Begna, T.; Jung, C. Effects of sequential exposures of sub-lethal doses of amitraz and thiacloprid on learning and memory of honey bee foragers, Apis mellifera. J. Asia Pac. Entomol. 2021, 24, 77–83. [Google Scholar] [CrossRef]
- Williamson, S.M.; Wright, G.A. Exposure to multiple cholinergic pesticides impairs olfactory learning and memory in honey bees. J. Exp. Biol. 2013, 216, 1799–1807. [Google Scholar] [PubMed]
- Tison, L.; Holtz, S.; Adeoye, A.; Kalkan, Ö.; Irmisch, N.S.; Lehmann, N.; Menzel, R. Effects of sublethal doses of thiacloprid and its formulation Calypso® on the learning and memory performance of honey bees. J. Exp. Biol. 2017, 220, 3695–3705. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Plant Name | Yield (%) (v/w) | Odor | Color |
|---|---|---|---|
| Eucalyptus globulus (leaf) | 1.10 | strong pungent | coloress |
| Rosmarinus officinalis (leaf) | 0.83 | intense spicy aroma. | pale yellow |
| Trachyspermum ammi (seed) | 3.7 | Aromatic odor | Pale yellow |
| Trachyspermum ammi a (seed) | 4.1 | Aromatic odor | Pale yellow |
| No | Compounds a | RI b | Ri c | E. globulus | R. officinalis | T. ammi d | T. ammi e |
|---|---|---|---|---|---|---|---|
| 1 | α-Pinene | 931.7 | 936.1 | 15.19 | 3.84 | 0.58 | 0.04 |
| 2 | β-Pinene | 973.5 | 977.7 | 1.84 | 2.42 | 4.56 | 0.78 |
| 3 | p-Cymene | 1027.3 | 1024.3 | 27.92 | 17.72 | ||
| 4 | 1,8- Cineole | 1033.1 | 1031.8 | 56.72 | 29.29 | ||
| 5 | γ-Terpinene | 1061.2 | 1059.7 | 1.88 | 32.72 | 17.02 | |
| 6 | Camphor | 1140.6 | 1143.4 | 16.08 | |||
| 7 | Isoborneol | 1156.9 | 1158.2 | 7.32 | |||
| 8 | α-Terpineol | 1190.4 | 1187.7 | 7.28 | 0.28 | 0.17 | |
| 9 | Carvestrene | 1195.2 | 8.58 | ||||
| 10 | Thymol | 1292.5 | 1290.1 | 0.94 | 24.36 | 59.40 | |
| 11 | Carvacrol | 1301.4 | 1300.4 | 0.51 | 0.12 | ||
| 12 | Cis-Caryophyllene | 1404.0 | 1406.5 | 6.69 |
| Treatments | Probit Analysis | |||||
|---|---|---|---|---|---|---|
| N | LC50 (95% CL) (µg/mL) | Slope ± SE | Intercept | χ2 | df | |
| Eucalyptus globulus | 75 | 4341.0 (2218.0–11,833.6) | 3.9 ± 0.8 | 4.5 | 13 | |
| Rosmarinus officinalis | 75 | 2577.0 (2013.0–3284.0) | 4.5 ± 0.9 | 20.3 | 47.8 | 13 |
| Trachyspermum ammi a | 75 | 98.0 (84.0–133.6) | 2.7 ± 0.5 | 0.5 | 10.4 | 13 |
| Trachyspermum ammi b | 75 | 81.0 (60.2–108.7) | 3.2 ± 0.6 | −1.1 | 5.6 | 13 |
| Thymol | 75 | 71.0 (52.5–95.8) | 3.0 ± 0.6 | −0.5 | 10.9 | 13 |
| Carvacrol | 75 | 106.0 (76.5–137.4) | 3.9 ± 0.9 | −2.9 | 5.5 | 13 |
| Γ-Terpinene | 75 | 1339.0 (848.0–3032.2) | 1.6 ± 0.4 | −0.1 | 5.1 | 13 |
| p-cymene | 75 | >5000.0 (-) | - | - | - | - |
| 1,8-Cineole | 75 | >10,000.0 (-) | - | - | - | - |
| Thymol:carvacrol | 75 | 42.0 (35.2–47.9) | 10.4 ± 2.9 | 11.8 | 4.1 | 13 |
| Thymol:γ-terpinene | 75 | 146.0 (112.7–222.8) | 7.8 ± 2.2 | −12.1 | 2.3 | 13 |
| Thymol:carvacrol:γ-terpinene | 75 | 113.0 (80.2–164.7) | 1.5 ± 0.3 | 1.1 | 7.6 | 13 |
| Fluvalinate | 75 | 143.0 (51.6–1576) | 0.8 ± 0.2 | 1.8 | 85.4 | 13 |
| Samples | Probit Analysis | |||||
|---|---|---|---|---|---|---|
| N | LD50 (95% CL) µg/bee | Slope ± SE | χ2 | Intercept | df | |
| Thymus schimperi | 180 | 7.9 (3.1–14.6) | 0.5 ± 0.2 | 10.1 | 4.6 | 13 |
| Trachyspermum ammi | 180 | 14.7 (6.9–156.0) | 0.8 ± 0.2 | 11.8 | 4.1 | 13 |
| Thymol | 180 | 4.6 (3.0–10.2) | 1.4 ± 0.3 | 19.3 | 4.1 | 13 |
| Carvacrol | 180 | 9.9 (6.5–21.3) | 1.5 ± 0.3 | 12.3 | 3.5 | 13 |
| γ-Terpinene | 180 | 86.5 (47.6–410.8) | 0.8 ± 0.2 | 8.6 | 3.4 | 13 |
| Amitraz | 180 | 5.3 (2.1–47.7) | 1.1 ± 0.2 | 39.2 | 4.2 | 13 |
| Treatments | |||
|---|---|---|---|
| Thymus schimperi | 27 | 91 | 3.4 |
| Trachyspermum ammi | 9.2 | 48 | 5.2 |
| Carvacrol | 28 | 38 | 1.4 |
| Thymol | 4.4 | 6.5 | 1.5 |
| γ-terpinene | 11.8 | 38 | 3.2 |
| Fluvalinate | 0.8 | 0.9 | 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Begna, T.; Ulziibayar, D.; Bisrat, D.; Jung, C. Acaricidal Toxicity of Four Essential Oils, Their Predominant Constituents, Their Mixtures against Varroa Mite, and Their Selectivity to Honey Bees (Apis cerana and A. mellifera). Insects 2023, 14, 735. https://doi.org/10.3390/insects14090735
Begna T, Ulziibayar D, Bisrat D, Jung C. Acaricidal Toxicity of Four Essential Oils, Their Predominant Constituents, Their Mixtures against Varroa Mite, and Their Selectivity to Honey Bees (Apis cerana and A. mellifera). Insects. 2023; 14(9):735. https://doi.org/10.3390/insects14090735
Chicago/Turabian StyleBegna, Tekalign, Delgermaa Ulziibayar, Daniel Bisrat, and Chuleui Jung. 2023. "Acaricidal Toxicity of Four Essential Oils, Their Predominant Constituents, Their Mixtures against Varroa Mite, and Their Selectivity to Honey Bees (Apis cerana and A. mellifera)" Insects 14, no. 9: 735. https://doi.org/10.3390/insects14090735